
Potential of the Liquid Fermentation of Fishery Waste by Paenibacillus elgii for Metalloprotease Production

 ,
,  ,
, 
 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Protease Assay
2.3. Screening the Production Conditions
2.4. Optimization of Production
2.5. Enzyme Purification
2.6. Biochemical Characterization of Purified Paenibacillus elgii TKU051 Protease
2.7. Hydrolysis of Fishery Wastes
2.8. Assay to Determine Free-Radical Scavenging Activity
3. Results and Discussion
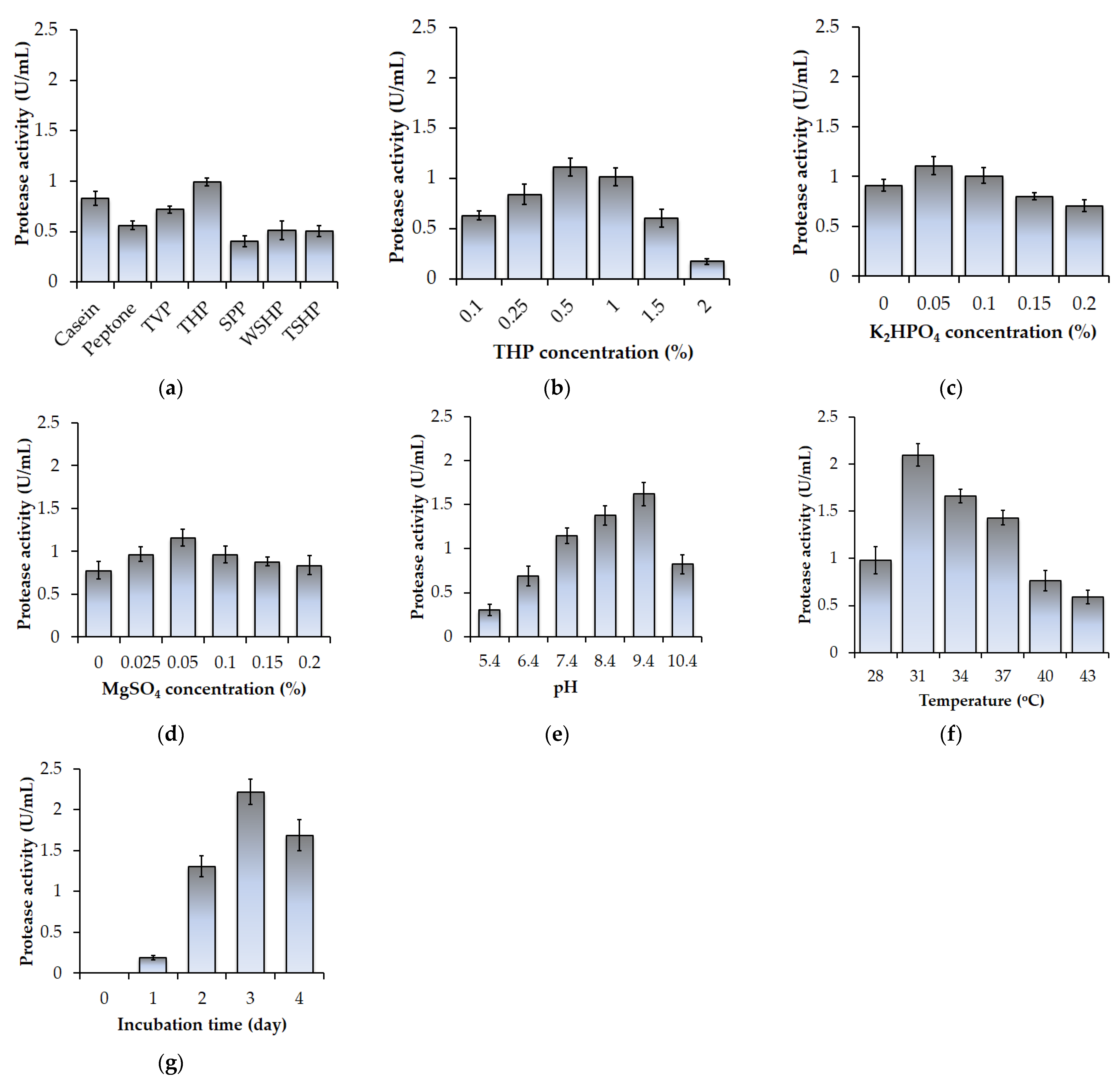
3.1. Screening for Optimal Protease-Producing Conditions Using Fishery Wastes as the Unique Carbon and Nitrogen (C/N) Source
3.2. Optimization Production of Paenibacillus elgii TKU051 Protease
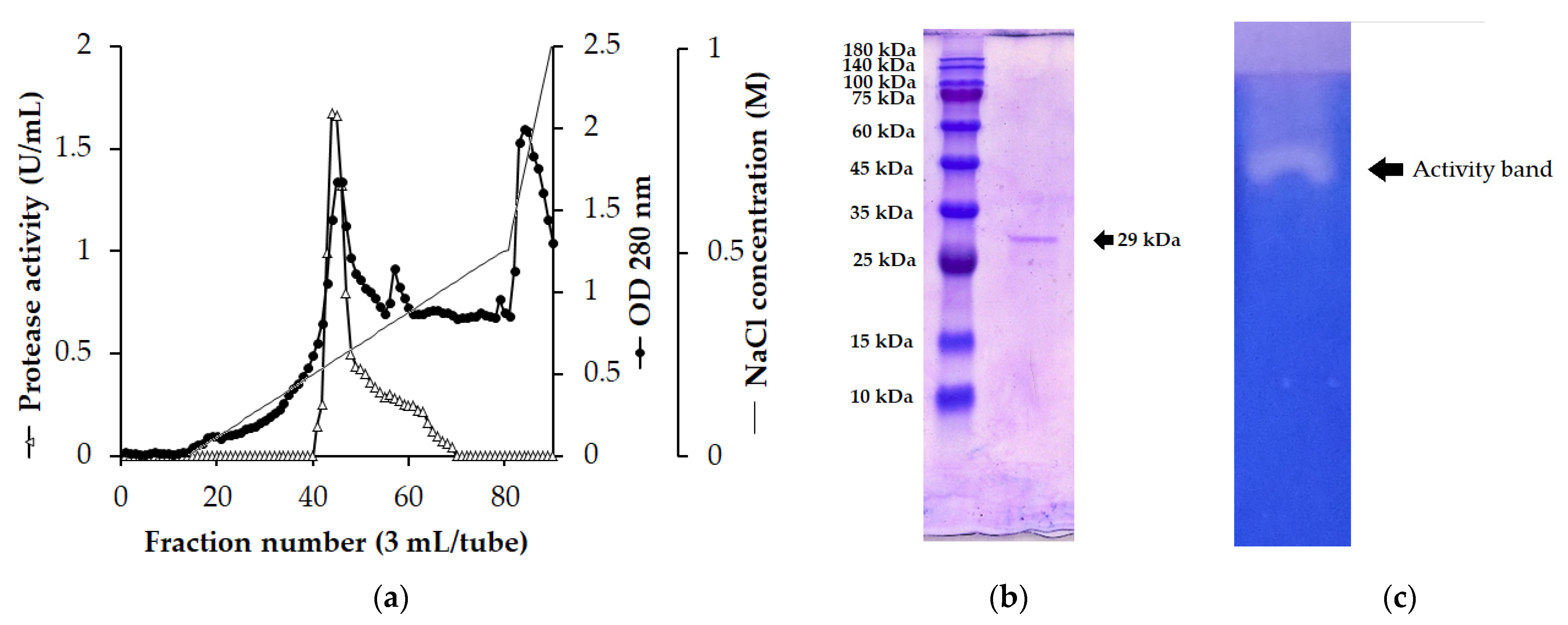
3.3. Enzyme Purification
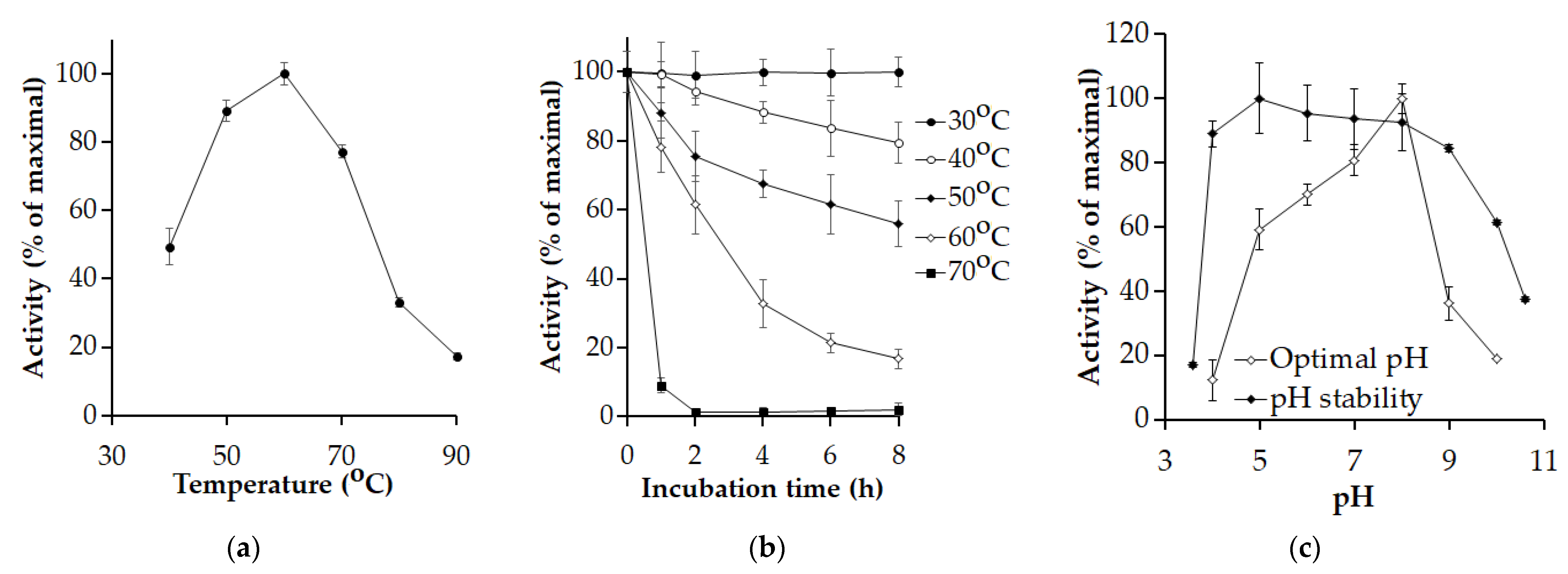
3.4. Enzyme Biochemical Properties
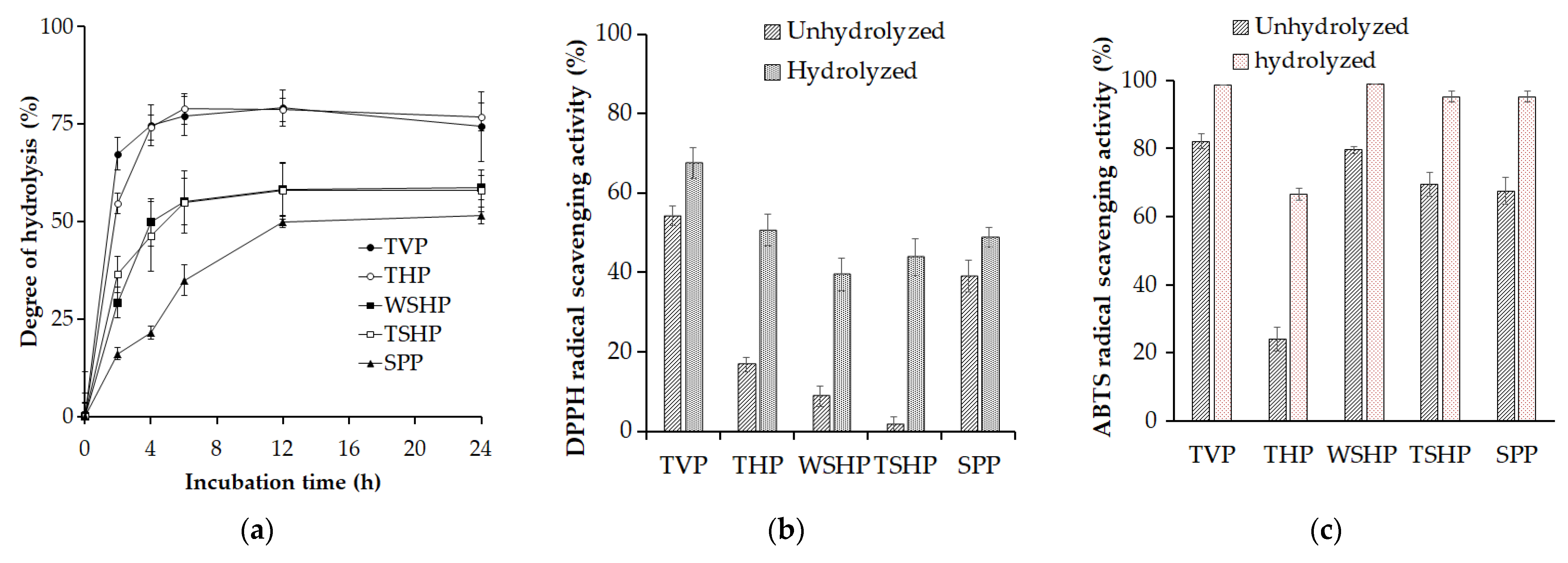
3.5. Application of Paenibacillus elgii TKU051 Protease in Hydrolyzing Fishery Wastes and Evaluation of the Antioxidant Activity of Fishery Waste Hydrolysates
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- López-Otín, C.; Bond, J.S. Proteases: Multifunctional enzymes in life and disease. J. Biol. Chem. 2008, 283, 30433–30437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.-H.; Doan, C.T.; Tran, T.N.; Nguyen, V.B.; Nguyen, A.D.; Wang, C.-L.; Wang, S.-L. Proteases production and chitin preparation from the liquid fermentation of chitinous fishery by-products by Paenibacillus elgii. Mar. Drugs 2021, 19, 477. [Google Scholar] [CrossRef]
- Ben Rebah, F.; Miled, N. Fish processing wastes for microbial enzyme production: A review. 3 Biotech 2013, 3, 255–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adetunji, A.I.; Olaniran, A.O. Statistical modelling and optimization of protease production by an autochthonous Bacillus aryabhattai Ab15-ES: A response surface methodology approach. Biocatal. Agric. Biotechnol. 2020, 24, 101528. [Google Scholar] [CrossRef]
- Doan, C.T.; Tran, T.N.; Nguyen, M.T.; Nguyen, V.B.; Nguyen, A.D.; Wang, S.L. Anti-α-glucosidase activity by a protease from Bacillus licheniformis. Molecules 2019, 24, 691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emran, M.A.; Ismail, S.A.; Hashem, A.M. Production of detergent stable thermophilic alkaline protease by Bacillus licheniformis ALW1. Biocatal. Agric. Biotechnol. 2020, 26, 101631. [Google Scholar] [CrossRef]
- Doan, C.T.; Tran, T.N.; Nguyen, V.B.; Nguyen, A.D.; Wang, S. Conversion of squid pens to chitosanases and proteases via Paenibacillus sp. TKU042. Mar. Drugs 2018, 16, 83. [Google Scholar] [CrossRef] [Green Version]
- Doan, C.T.; Tran, T.N.; Wang, C.-L.; Wang, S.-L. Microbial conversion of shrimp heads to proteases and chitin as an effective dye adsorbent. Polymers 2020, 12, 2228. [Google Scholar] [CrossRef]
- Jadhav, H.P.; Sonawane, M.S.; Khairnar, M.H.; Sayyed, R.Z. Production of alkaline protease by rhizospheric Bacillus cereus HP_RZ17 and Paenibacillus xylanilyticus HP_RZ19. Environ. Sustain. 2020, 3, 5–13. [Google Scholar] [CrossRef]
- El-Khonezy, M.I.; Elgammal, E.W.; Ahmed, E.F.; Abd-Elaziz, A.M. Detergent stable thiol-dependant alkaline protease produced from the endophytic fungus Aspergillus ochraceus BT21: Purification and kinetics. Biocatal. Agric. Biotechnol. 2021, 35, 102046. [Google Scholar] [CrossRef]
- da Silva, O.S.; de Almeida, E.M.; de Melo, A.H.F.; Porto, T.S. Purification and characterization of a novel extracellular serine-protease with collagenolytic activity from Aspergillus tamarii URM4634. Int. J. Biol. Macromol. 2018, 117, 1081–1088. [Google Scholar] [CrossRef] [PubMed]
- Dhamodharan, D.; Jemimah, N.S.; Merlyn, K.S.; Subathra, D.C. Novel fibrinolytic protease producing Streptomyces radiopugnans VITSD8 from marine sponges. Mar. Drugs 2019, 17, 164. [Google Scholar]
- Sarkar, G.; Suthindhiran, K. Extraction and characterization of alkaline protease from Streptomyces sp. GS-1 and its application as dehairing agent. Biocatal. Agric. Biotechnol. 2020, 25, 101590. [Google Scholar] [CrossRef]
- Sivaprakasam, S.; Dhandapani, B.; Mahadevan, S. Optimization studies on production of a salt-tolerant protease from Pseudomonas aeruginosa strain BC1 and its application on tannery saline wastewater treatment. Braz. J. Microbiol. 2011, 42, 1506–1511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Chio, S. Deproteinization of shrimp and crab shell with the protease of Pseudomonas aeruginosa K-187. Enzyme Microb. Technol. 1998, 22, 629–633. [Google Scholar] [CrossRef]
- Doan, C.T.; Tran, T.N.; Vo, T.P.K.; Nguyen, A.D.; Wang, S. Chitin extraction from shrimp waste by liquid fermentation using an alkaline protease-producing strain, Brevibacillus parabrevis. Int. J. Biol. Macromol. 2019, 131, 706–715. [Google Scholar] [CrossRef]
- Gomri, M.A.; Rico-Díaz, A.; Escuder-Rodríguez, J.-J.; El Moulouk Khaldi, T.; González-Siso, M.-I.; Kharroub, K. Production and characterization of an extracellular acid protease from thermophilic Brevibacillus sp. OA30 isolated from an algerian hot spring. Microorganisms 2018, 6, 31. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.-H.; Zhang, X.-Y.; Wang, Z.; Zhang, X.; Liu, S.-C.; Song, X.-Y.; Zhang, Y.-Z.; Ding, J.-M.; Chen, X.-L.; Xu, F. Potential of thermolysin-like protease a69 in preparation of bovine collagen peptides with moisture-retention ability and antioxidative activity. Mar. Drugs 2021, 19, 676. [Google Scholar] [CrossRef]
- Doan, C.T.; Tran, T.N.; Nguyen, V.B.; Nguyen, A.D.; Wang, S.-L. Utilization of seafood processing by-products for production of proteases by Paenibacillus sp. TKU052 and their application in biopeptides’ preparation. Mar. Drugs 2020, 18, 574. [Google Scholar] [CrossRef]
- Chakrabarti, S.; Guha, S.; Majumder, K. Food-Derived Bioactive Peptides in Human Health: Challenges and Opportunities. Nutrients 2018, 10, 1738. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.H.; Doan, C.T.; Nguyen, V.B.; Nguyen, A.D.; Wang, S.L. Reclamation of fishery processing waste: A mini-review. Molecules 2019, 24, 2234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messina, C.M.; Manuguerra, S.; Arena, R.; Renda, G.; Ficano, G.; Randazzo, M.; Fricano, S.; Sadok, S.; Santulli, A. In Vitro Bioactivity of Astaxanthin and Peptides from Hydrolisates of Shrimp (Parapenaeus longirostris) By-Products: From the Extraction Process to Biological Effect Evaluation, as Pilot Actions for the Strategy “From Waste to Profit”. Mar. Drugs 2021, 19, 216. [Google Scholar] [CrossRef]
- Joo, H.S.; Kumar, C.G.; Park, G.C.; Paik, S.R.; Chang, C.S. Oxidant and SDS-stable alkaline protease from Bacillus clausii I-52: Production and some properties. J. Appl. Microbiol. 2003, 95, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Haddar, A.; Fakhfakh-Zouari, N.; Hmidet, N.; Frikha, F.; Nasri, M.; Kamoun, A.S. Low-cost fermentation medium for alkaline protease production by Bacillus mojavensis A21 using hulled grain of wheat and sardinella peptone. J. Biosci. Bioeng. 2010, 110, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Doan, C.T.; Tran, T.N.; Wen, I.H.; Nguyen, V.B.; Nguyen, A.D.; Wang, S.L. Conversion of shrimp head waste for production of a thermotolerant, detergent-stable, alkaline protease by Paenibacillus sp. Catalysts 2019, 9, 798. [Google Scholar] [CrossRef] [Green Version]
- Hammami, A.; Bayoudh, A.; Abdelhedi, O.; Nasri, M. Low-cost culture medium for the production of proteases by Bacillus mojavensis SA and their potential use for the preparation of antioxidant protein hydrolysate from meat sausage by-products. Ann. Microbiol. 2018, 68, 473–484. [Google Scholar] [CrossRef]
- Elumalai, P.; Lim, J.M.; Park, Y.J.; Cho, M.; Shea, P.J.; Oh, B.T. Agriculture waste materials enhance protease production by Bacillus subtilis B22 in submerged fermentation under blue light-emitting diodes. Bioprocess Biosyst. Eng. 2020, 43, 821–830. [Google Scholar] [CrossRef]
- Ramkumar, A.; Sivakumar, N.; Gujarathi, A.M.; Victor, R. Production of thermotolerant, detergent stable alkaline protease using the gut waste of Sardinella longiceps as a substrate: Optimization and characterization. Sci. Rep. 2018, 8, 12442. [Google Scholar] [CrossRef]
- Vazquez, J.A.; Docasal, S.F.; Miron, J.; Gonzalez, M.P.; Murado, M.A. Proteases production by two Vibrio species on residuals marine media. J. Ind. Microbiol. Biotechnol. 2006, 33, 661–668. [Google Scholar] [CrossRef] [Green Version]
- Ramkumar, A.; Sivakumar, N.; Victor, R. Fish Waste-Potential Low Cost Substrate for Bacterial Protease Production: A Brief Review. Open Biotechnol. J. 2016, 10, 335–341. [Google Scholar] [CrossRef]
- Sellami-Kamoun, A.; Ghorbel-Frikha, G.; Haddar, A.; Nasri, M. Enhanced Bacillus cereus BG1 protease production by the use of sardinelle (Sardinella aurita) powder. Ann. Microbiol. 2011, 61, 273–280. [Google Scholar] [CrossRef]
- Liu, C.L.; Lin, T.H.; Juang, R.S. Optimization of recombinant hexaoligochitin-producing chitinase production with response surface methodology. Int. J. Biol. Macromol. 2013, 62, 518–522. [Google Scholar] [CrossRef]
- Rai, S.K.; Roy, J.K.; Mukherjee, A.K. Characterisation of a detergent-stable alkaline protease from a novel thermophilic strain Paenibacillus tezpurensis sp. nov. AS-S24-II. Appl. Microbiol. Biotechnol. 2010, 85, 1437–1450. [Google Scholar] [CrossRef] [PubMed]
- Hrabák, J.; Martínek, K. Screening of secreted proteases of Paenibacillus larvae by using substrate-SDS-polyacrylamide gel electrophoresis. J. Apic. Res. 2007, 46, 160–164. [Google Scholar] [CrossRef]
- Lu, F.; Lu, Z.; Bie, X.; Yao, Z.; Wang, Y.; Lu, Y.; Guo, Y. Purification and characterization of a novel anticoagulant and fibrinolytic enzyme produced by endophytic bacterium Paenibacillus polymyxa EJS-3. Thromb. Res. 2010, 126, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Pan, Y.; She, Q.; Chen, L. A novel carboxyl-terminal protease derived from Paenibacillus lautus CHN26 exhibiting high activities at multiple sites of substrates. BMC Biotechnol. 2013, 13, 89. [Google Scholar] [CrossRef] [Green Version]
- Hang, F.; Liu, P.; Wang, Q.; Han, J.; Wu, Z.; Gao, C.; Liu, Z.; Zhang, H.; Chen, W. High milk-clotting activity expressed by the newly isolated Paenibacillus spp. strain BD3526. Molecules 2016, 21, 73. [Google Scholar] [CrossRef] [Green Version]
- Hang, F.; Wang, Q.; Hong, Q.; Gao, C.; Zhang, H.; Chen, W. Structural insight into a novel neutral metalloproteinase from Paenibacillus spp. BD3526: Implications for mechanisms of rapid inactivation and calcium-dependent stability. Int. J. Biol. Macromol. 2017, 95, 1082–1090. [Google Scholar] [CrossRef]
- Ruf, A.; Stihle, M.; Benz, J.; Schmidt, M.; Sobek, H. Structure of gentlyase, the neutral metalloprotease of Paenibacillus polymixa. Acta Crystallogr. D Biol. Crystallogr. 2013, 69, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Nam, K.H. Structural analysis of metal chelation of the metalloproteinase thermolysis by 1,10-phenanthroline. J. Inorg. Biochem. 2021, 215, 111319. [Google Scholar] [CrossRef]
- Hasan, R.; Rony, M.N.H.; Ahmed, R. In silico characterization and structural modeling of bacterial metalloprotease of family M4. J. Genet. Eng. Biotechnol. 2021, 19, 25. [Google Scholar] [CrossRef] [PubMed]
- Mahmoodani, F.; Ghassem, M.; Babji, A.S.; Yusop, S.M.; Khosrokhavar, R. ACE inhibitory activity of pangasius catfish (Pangasius sutchi) skin and bone gelatin hydrolysate. J. Food Sci. Technol. 2014, 51, 1847–1856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, X.; Zhu, Z.; Wu, S.; Chen, M.; Huang, R.; Wang, J.; Wu, Q.; Ding, Y. Preparation of antioxidant protein hydrolysates from Pleurotus geesteranus and their protective effects on H2O2 oxidative damaged PC12 cells. Molecules 2020, 25, 5408. [Google Scholar] [CrossRef] [PubMed]
- Wołosiak, R.; Drużyńska, B.; Derewiaka, D.; Piecyk, M.; Majewska, E.; Ciecierska, M.; Worobiej, E.; Pakosz, P. Verification of the conditions for determination of antioxidant activity by ABTS and DPPH assays—A practical approach. Molecules 2022, 27, 50. [Google Scholar] [CrossRef] [PubMed]
- Amorati, R.; Valgimigli, L. Advantages and limitations of common testing methods for antioxidants. Free Radic. Res. 2015, 49, 633–649. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Term | Estimate | Standard Error | t Value | Pr (>|t|) | |
|---|---|---|---|---|---|
| (Intercept) | 2.3106984 | 0.0934004 | 24.7397 | <2.2 × 10−16 | *** |
| A | 0.4152748 | 0.0467002 | 8.8924 | 2.303 × 10−9 | *** |
| B | 0.0426658 | 0.0467002 | 0.9136 | 0.3693164 | |
| C | 0.0011805 | 0.0467002 | 0.0253 | 0.9800264 | |
| D | −0.5694113 | 0.0467002 | −12.1929 | 2.941 × 10−12 | *** |
| E | 0.0834766 | 0.0467002 | 1.7875 | 0.0855205 | |
| F | 0.1249619 | 0.0467002 | 2.6758 | 0.0127280 | * |
| AB | 0.0303551 | 0.0808871 | 0.3753 | 0.7104983 | |
| AC | 0.1373569 | 0.0808871 | 1.6981 | 0.1014219 | |
| AD | −0.1831425 | 0.0571958 | −3.2020 | 0.0035847 | ** |
| AE | 0.1016896 | 0.0808871 | 1.2572 | 0.2198650 | |
| AF | 0.0594454 | 0.0808871 | 0.7349 | 0.4689667 | |
| BC | 0.0713345 | 0.0808871 | 0.8819 | 0.3859106 | |
| BD | 0.0435090 | 0.0808871 | 0.5379 | 0.5952226 | |
| BE | −0.1457045 | 0.0571958 | −2.5475 | 0.0171091 | * |
| BF | −0.1654353 | 0.0808871 | −2.0453 | 0.0510727 | |
| CD | −0.0999189 | 0.0808871 | −1.2353 | 0.2277696 | |
| CE | 0.0672872 | 0.0808871 | 0.8319 | 0.4130605 | |
| CF | −0.0012648 | 0.0571958 | −0.0221 | 0.9825263 | |
| DE | 0.1431749 | 0.0808871 | 1.7701 | 0.0884449 | |
| DF | −0.0306081 | 0.0808871 | −0.3784 | 0.7082021 | |
| EF | −0.3268234 | 0.0808871 | −4.0405 | 0.0004206 | *** |
| A2 | −0.4784022 | 0.0713357 | −6.7063 | 4.084 × 10−7 | *** |
| B2 | −0.2828647 | 0.0713357 | −3.9653 | 0.0005118 | *** |
| C2 | −0.2403675 | 0.0713357 | −3.3695 | 0.0023591 | ** |
| D2 | −0.7733526 | 0.0713357 | −10.8410 | 3.844 × 10−11 | *** |
| E2 | −0.2434030 | 0.0713357 | −3.4121 | 0.0021192 | ** |
| F2 | −0.6719160 | 0.0713357 | −9.4191 | 7.262 × 10−10 | *** |
| Degrees of Freedom | Sum of Squares | Mean Square | F Value | Pr (>F) | |
|---|---|---|---|---|---|
| FO(A, B, C, D, E, F) | 6 | 12.5061 | 2.08435 | 39.8219 | 6.794 × 10−12 |
| TWI(A, B, C, D, E, F) | 15 | 2.5626 | 0.17084 | 3.2639 | 0.003989 |
| PQ(A, B, C, D, E, F) | 6 | 13.9118 | 2.31863 | 44.2979 | 1.972 × 10−12 |
| Residuals | 26 | 1.3609 | 0.05234 | ||
| Lack-of-fit | 21 | 1.0724 | 0.05107 | 0.8851 | 0.624650 |
| Pure error | 5 | 0.2885 | 0.05770 |
| Variable | Before Optimization | After Optimization | |
|---|---|---|---|
| By One-Factor-at-a-Time | By Response Surface Methodology | ||
| THP (%) | 1 | 0.5 | 0.811 |
| K2HPO4 (%) | 0.1 | 0.05 | 0.052 |
| MgSO4 (%) | 0.05 | 0.05 | 0.073 |
| Intitial pH | 7.4 | 9.4 | 8.96 |
| Temperature (°C) | 37 | 31 | 31.4 |
| Incubation time (day) | 3 | 3 | 3.092 |
| Protease activity (U/mL) | 1.111 ± 0.041 | 2.341 ± 0.076 | 2.635 ± 0.124 |
| Purification Step | Total Activity (U) | Total Protein (mg) | Specific Activity (U/mg) | Yield (%) | Purification Fold |
|---|---|---|---|---|---|
| Culture supernatant | 2153.452 | 3287.595 | 0.655 | 100.000 | 1.000 |
| (NH4)2SO4 precipitation | 995.930 | 349.447 | 2.850 | 46.248 | 4.351 |
| High Q chromatography | 201.220 | 44.642 | 4.507 | 9.344 | 6.881 |
| DEAE sepharose chromatography | 34.489 | 2.009 | 17.165 | 1.602 | 26.205 |
| Sephacryl S-200 chromatography | 24.953 | 0.815 | 30.603 | 1.159 | 46.721 |
| Chemical | Relative Activity (%) |
|---|---|
| None | 100 ± 5 |
| EDTA | 18 ± 6 |
| E-64 | 100 ± 4 |
| DTNB | 99 ± 9 |
| PMSF | 99 ± 5 |
| 1,10-phenanthroline | 5 ± 7 |
| 2-mercaptoethanol | 98 ± 7 |
| ZnCl2 | 85 ± 2 |
| FeCl2 | 115 ± 2 |
| CaCl2 | 120 ± 3 |
| CuCl2 | 21 ± 3 |
| MgCl2 | 107 ± 7 |
| MnCl2 | 72 ± 4 |
| BaCl2 | 103 ± 3 |
| 1 ZnCl2 | 84 ± 3 |
| 1 FeCl2 | 16 ± 1 |
| 1 CaCl2 | 72 ± 4 |
| 1 CuCl2 | 19 ± 7 |
| 1 MgCl2 | 18 ± 2 |
| 1 MnCl2 | 53 ± 8 |
| 1 BaCl2 | 45 ± 4 |
| Cetrimonium bromide | 8 ± 1 |
| Triton X-100 | 53 ± 1 |
| Tween 40 | 95 ± 2 |
| Tween 20 | 94 ± 4 |
| SDS | 0 ± 0 |
| 2 Ekos | 94 ± 1 |
| 2 Amah | 63 ± 9 |
| 2 Yeuhyang | 93 ± 4 |
| Substrate | Relative Activity (%) |
|---|---|
| Casein | 100 ± 6 |
| Gelatin | 16 ± 2 |
| Albumin | 54 ± 8 |
| Keratin | 57 ± 8 |
| Hemoglobin | 47 ± 5 |
| Fibrinogen | 52 ± 4 |
| TVP | 313 ± 23 |
| THP | 117 ± 22 |
| TSHP | 166 ± 22 |
| WSHP | 124 ± 26 |
| SPP | 163 ± 24 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doan, C.T.; Tran, T.N.; Nguyen, M.T.; Nguyen, H.K.; Tran, T.K.T.; Nguyen, T.H.; Tran, T.P.H.; Nguyen, V.B.; Nguyen, A.D.; Wang, S.-L. Potential of the Liquid Fermentation of Fishery Waste by Paenibacillus elgii for Metalloprotease Production. Polymers 2022, 14, 2741. https://doi.org/10.3390/polym14132741
Doan CT, Tran TN, Nguyen MT, Nguyen HK, Tran TKT, Nguyen TH, Tran TPH, Nguyen VB, Nguyen AD, Wang S-L. Potential of the Liquid Fermentation of Fishery Waste by Paenibacillus elgii for Metalloprotease Production. Polymers. 2022; 14(13):2741. https://doi.org/10.3390/polym14132741
Chicago/Turabian StyleDoan, Chien Thang, Thi Ngoc Tran, Minh Trung Nguyen, Huu Kien Nguyen, Thi Kim Thi Tran, Thi Hanh Nguyen, Thi Phuong Hanh Tran, Van Bon Nguyen, Anh Dzung Nguyen, and San-Lang Wang. 2022. "Potential of the Liquid Fermentation of Fishery Waste by Paenibacillus elgii for Metalloprotease Production" Polymers 14, no. 13: 2741. https://doi.org/10.3390/polym14132741