
Hydrodynamic and Polyelectrolyte Properties of Actin Filaments: Theory and Experiments


Abstract
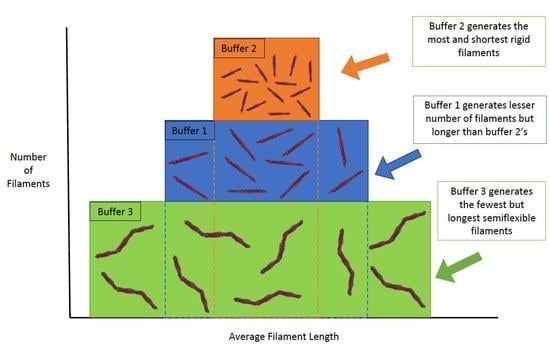
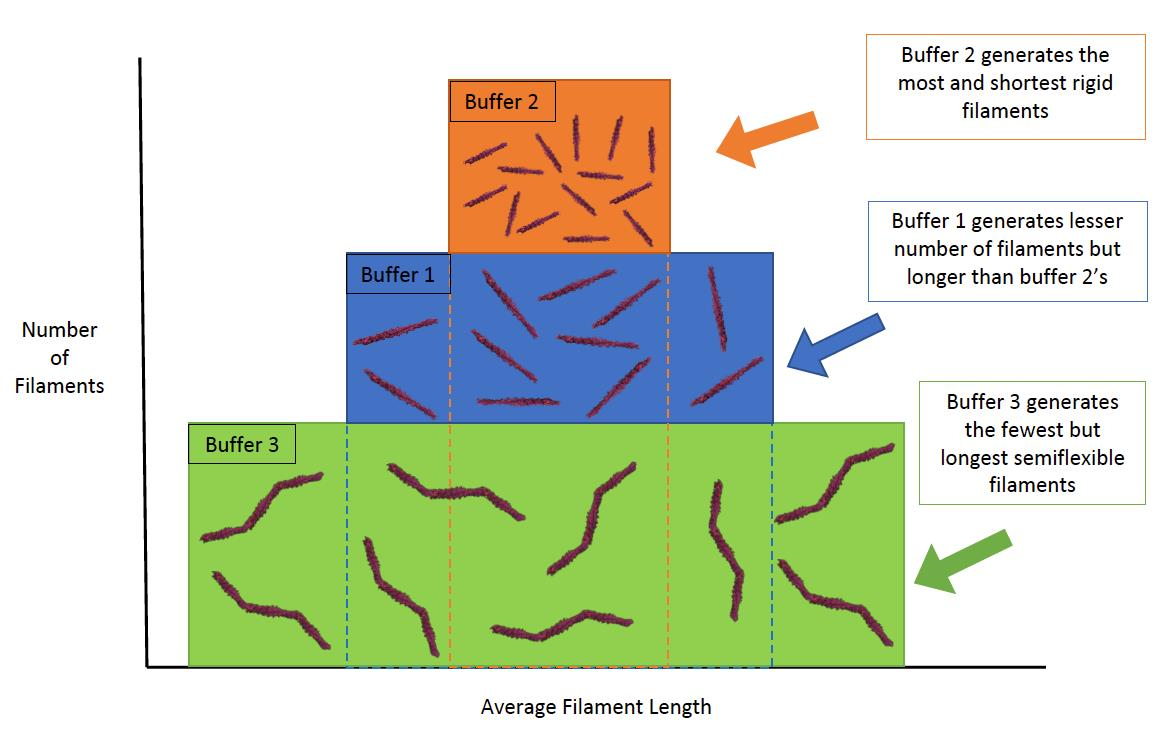
:
1. Introduction
2. Materials and Methods: Theory
2.1. Diffusion Theory
2.2. Electrophoretic Mobility Theory
2.3. Length Distribution Theory
2.4. Polydispersity Theory
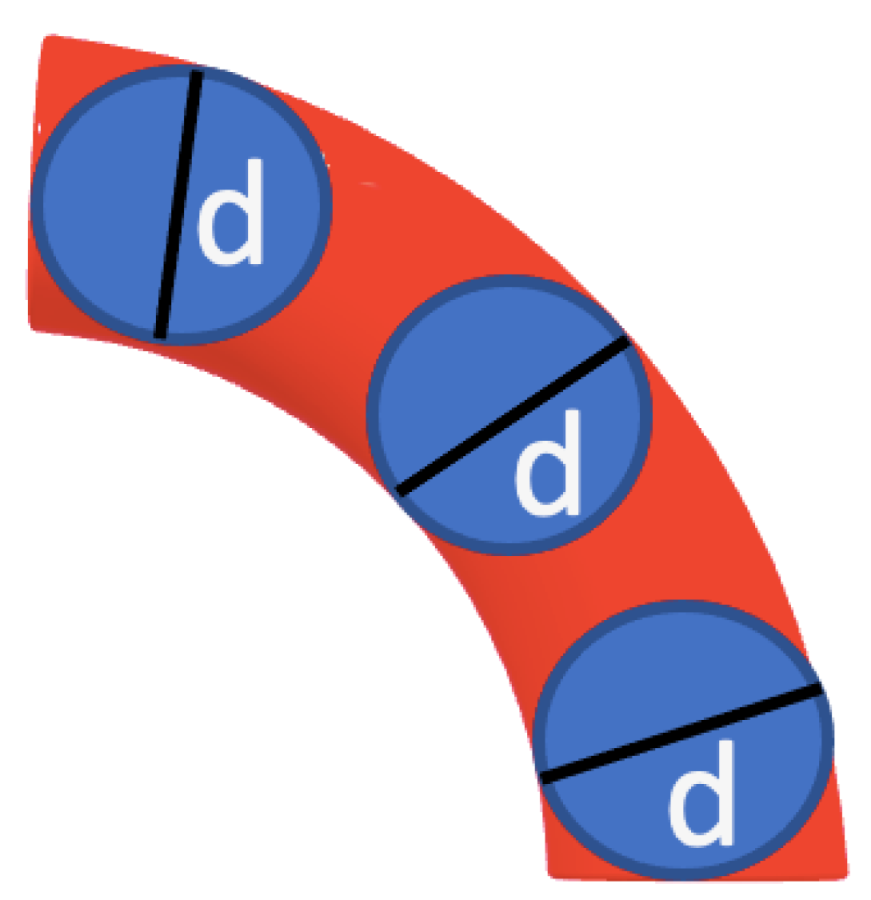
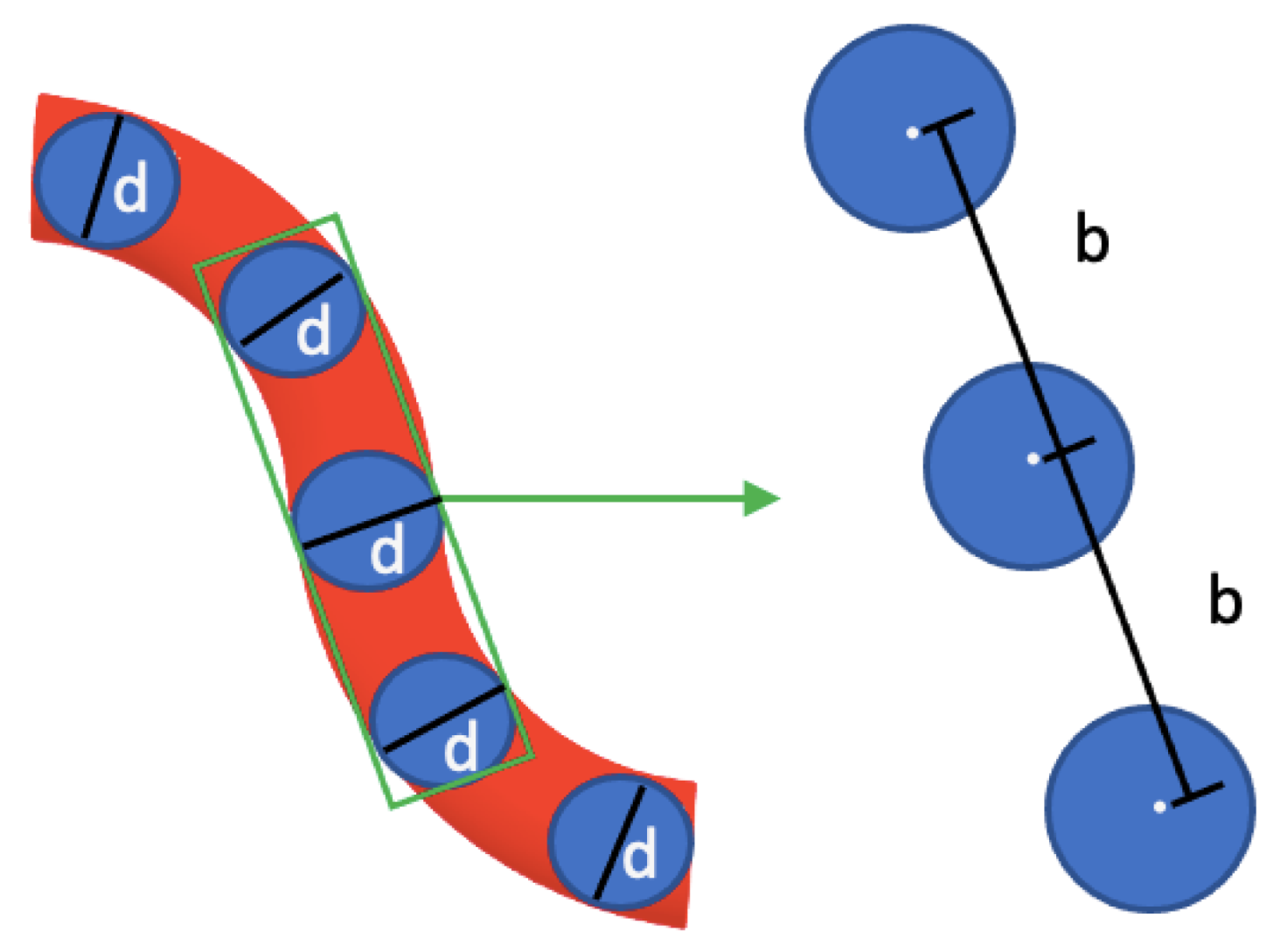
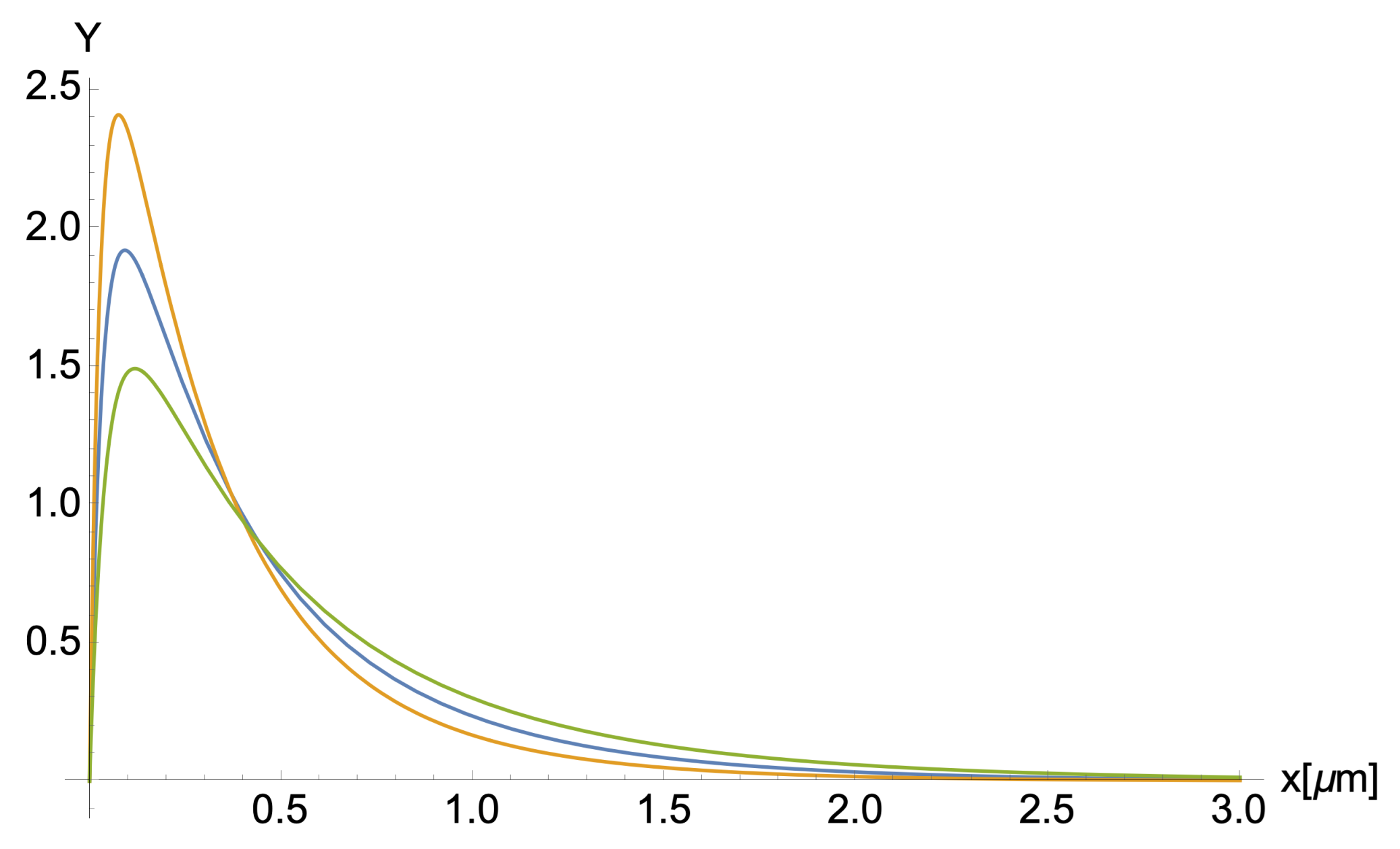
2.5. Dynamic Structure Factor Theory for Semiflexible Polymers
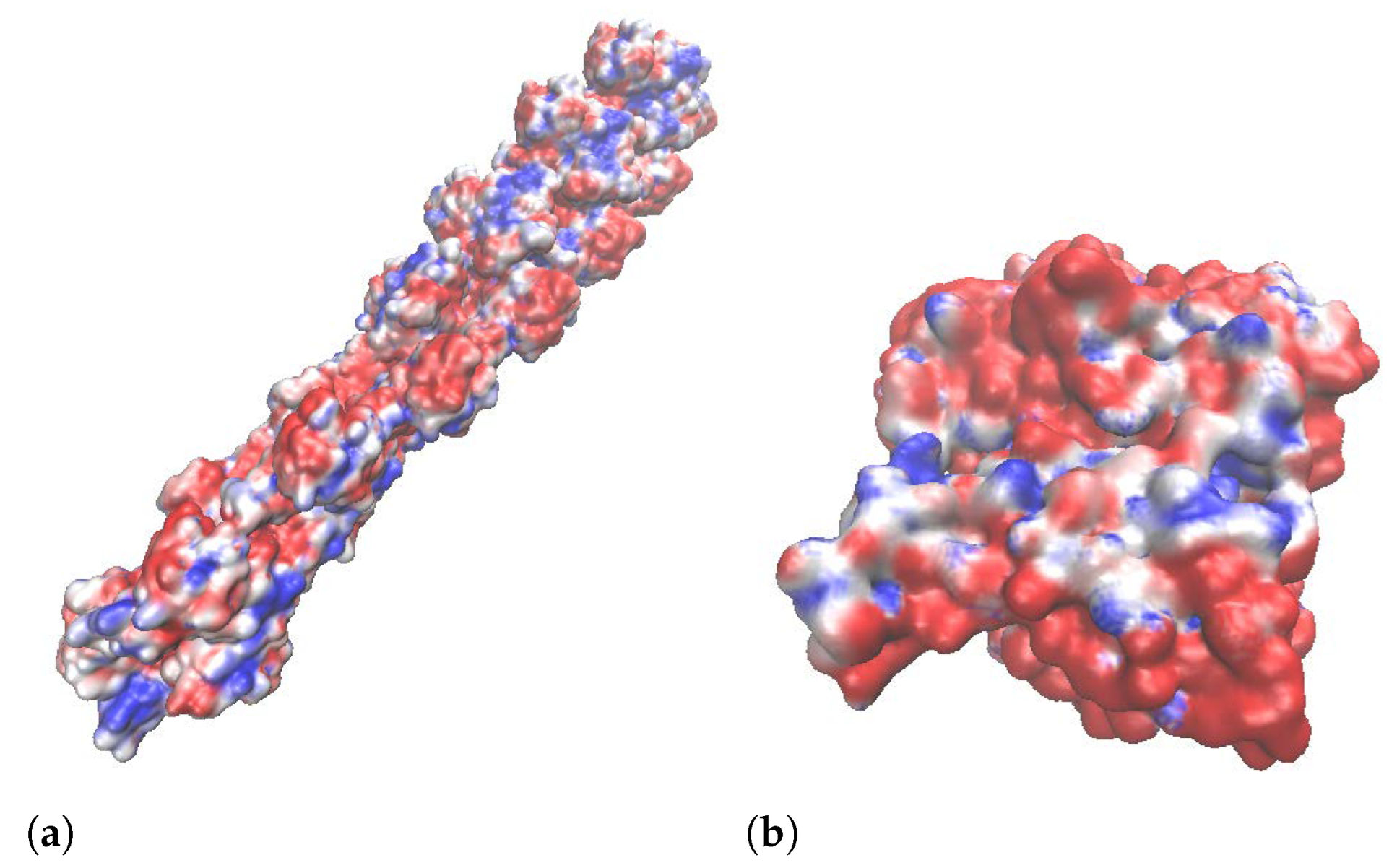
2.6. Zeta Potential
2.7. Parameters Calculation
3. Materials and Methods: Experiments
3.1. Sample Preparation
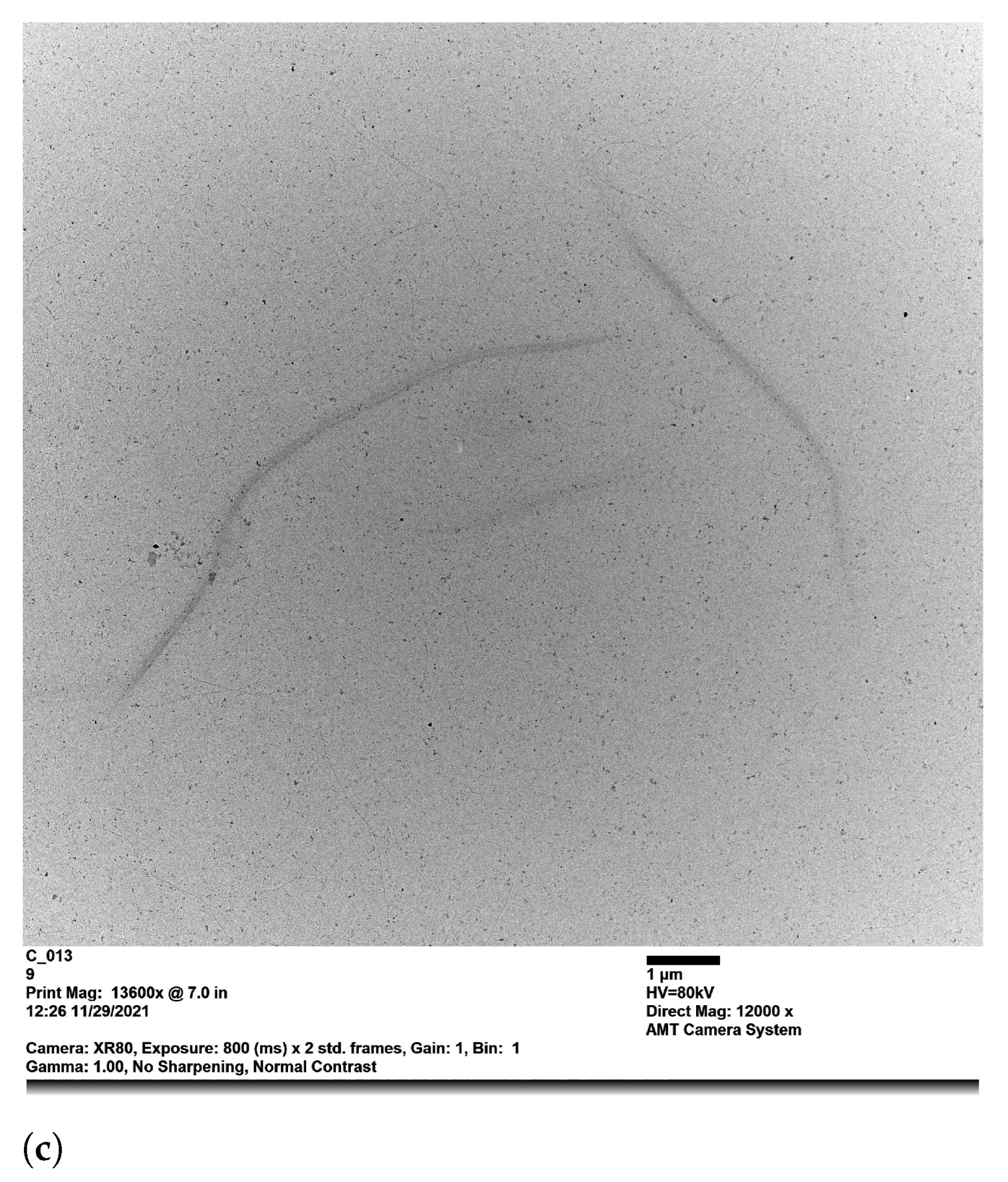
3.2. Light Scattering Experiments
4. Results
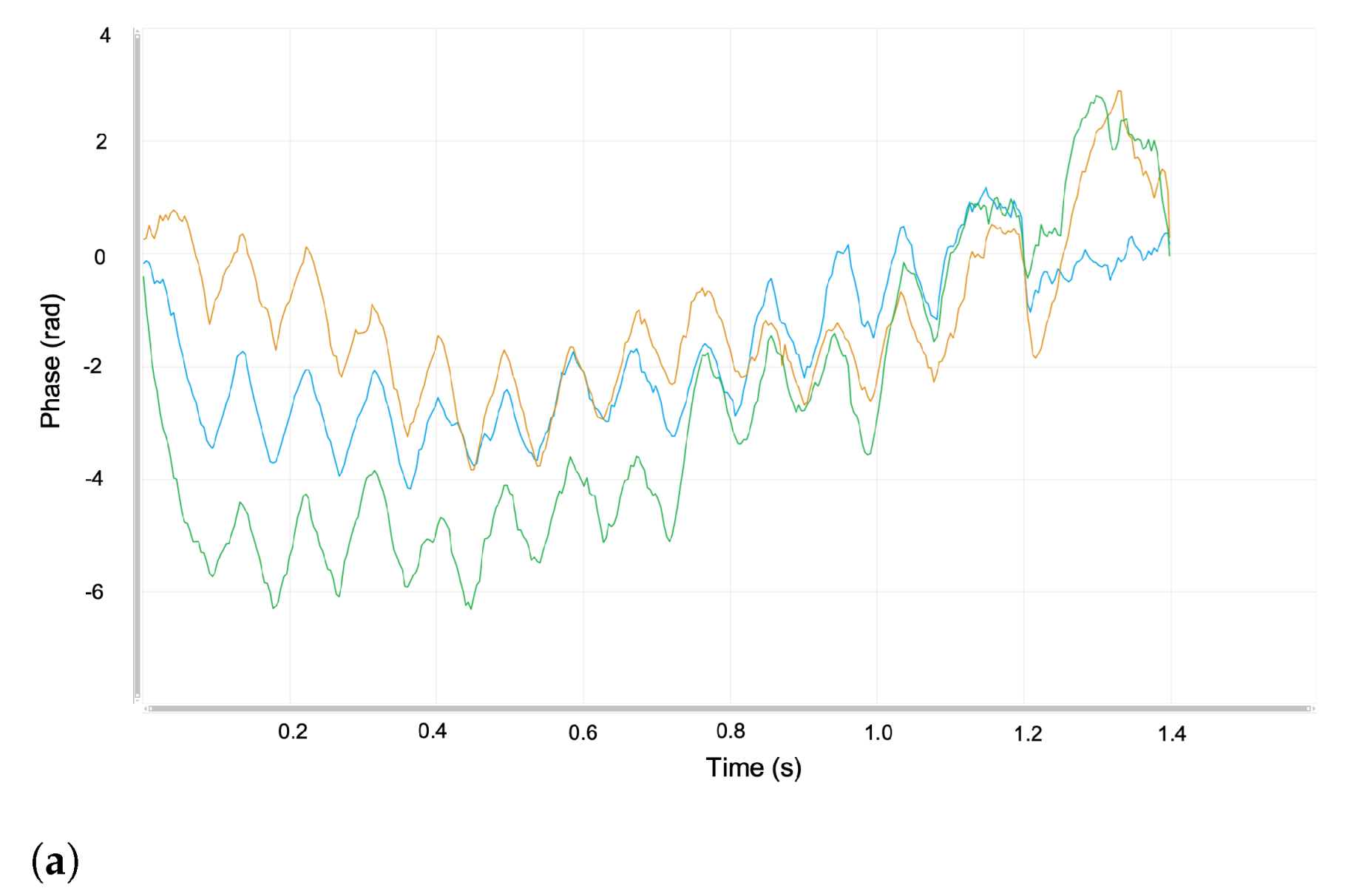
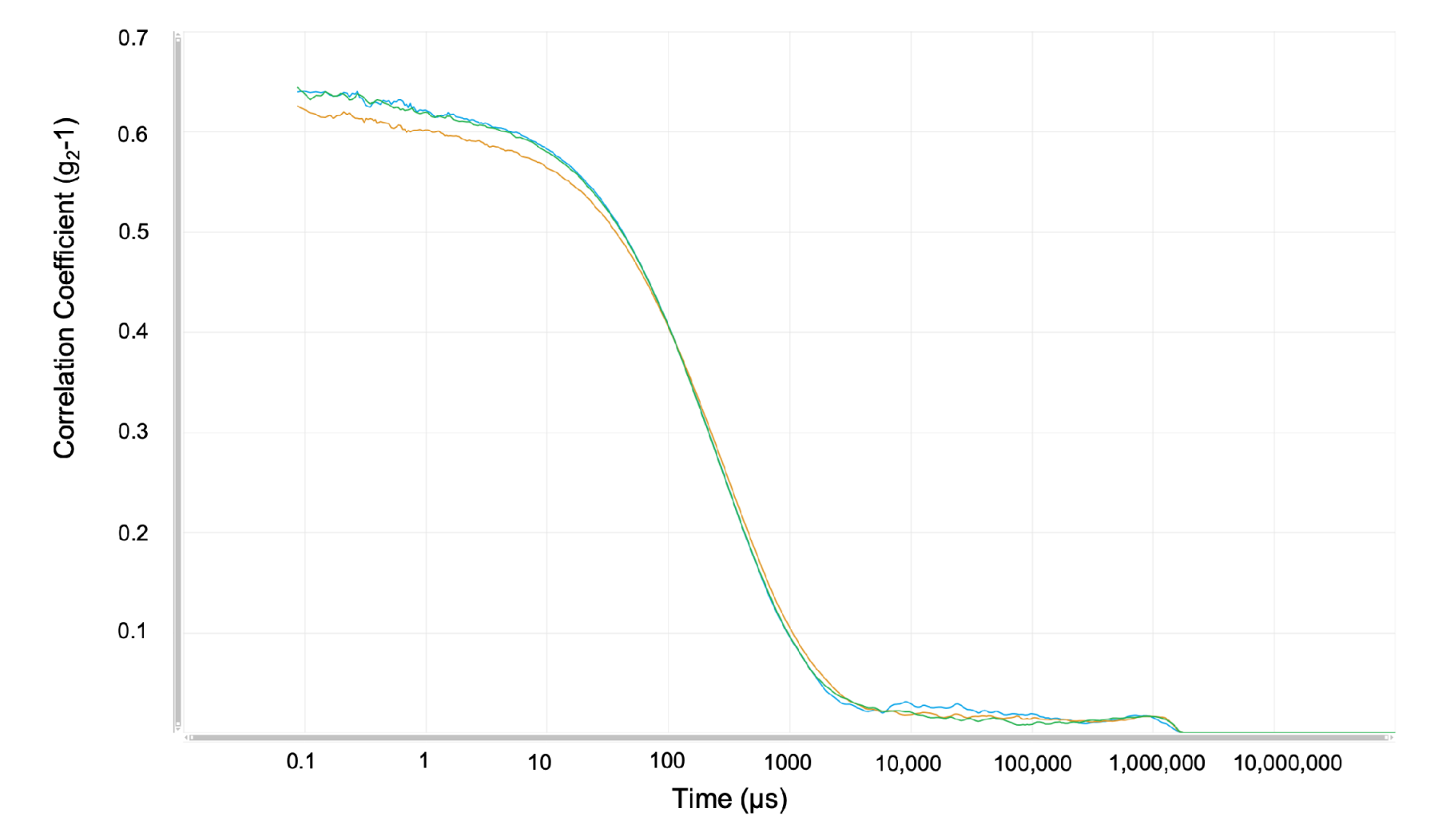
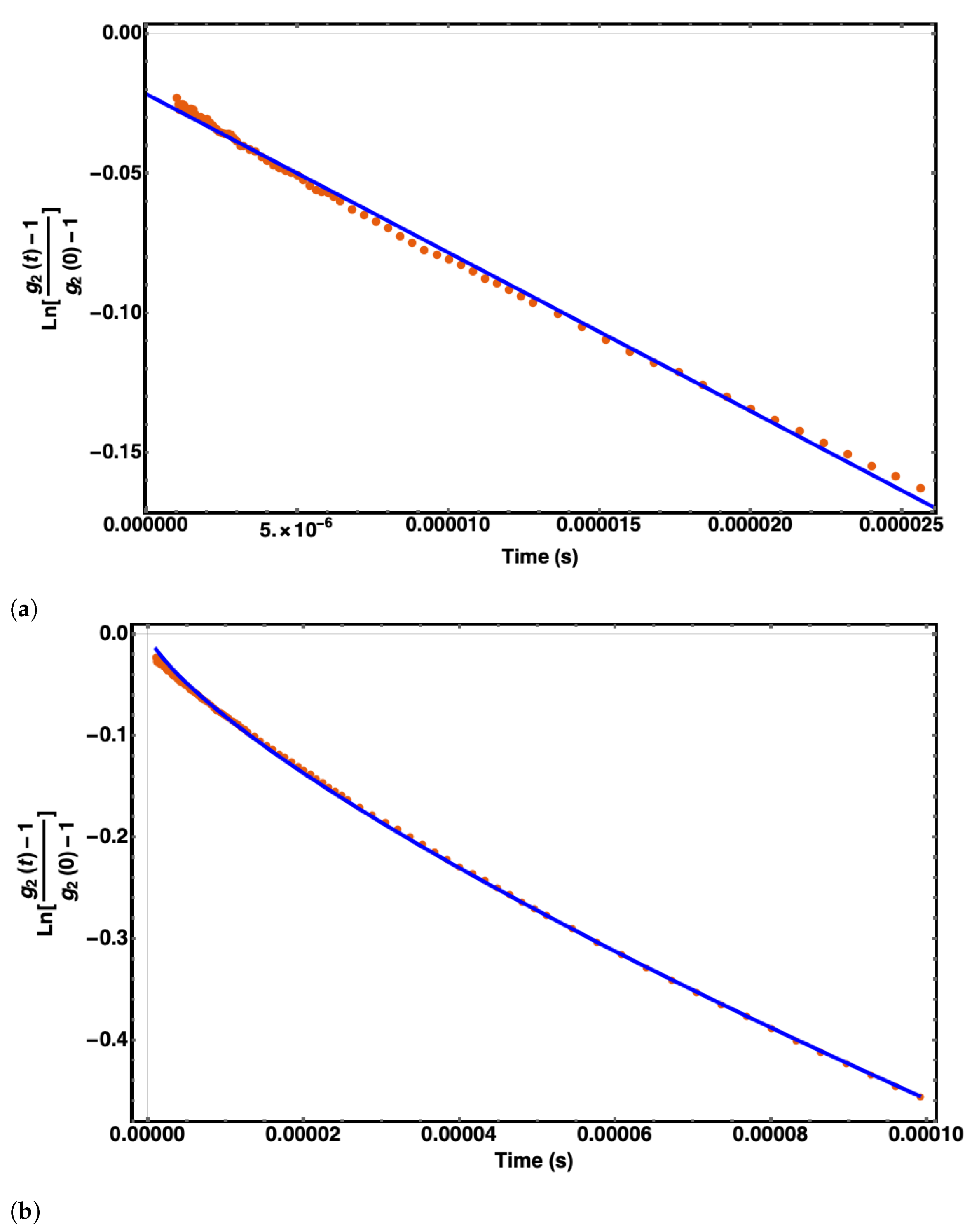
4.1. Experiments
4.2. Theory
5. Discussion
5.1. Length Distribution
5.2. Structural Parameters
5.3. Translational Diffusion Coefficient and Electrophoretic Mobility
5.4. Other Properties
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, F.; Sampogna, R.V.; Ware, B.R. pH Dependence of Actin Self-Assembly. Biophys. J. 1989, 55, 293–298. [Google Scholar] [CrossRef] [Green Version]
- Lanni, F.; Ware, B.R. Detection and Characterization of Actin Monomers, Oligomers, and Filaments in Solution by Measurement of Fluorescence Photobleaching Recovery. Biophys. J. 1984, 46, 97–110. [Google Scholar] [CrossRef] [Green Version]
- Kas, J.; Strey, H.; Tang, J.X.; Finger, D.; Ezzell, R.; Sackmann, E.; Janmey, P.A. F-actin, A Model Polymer for Semiflexible Chains in Dilute, Semidilute, and Liquid Crystalline Solutions. Biophys. J. 1996, 70, 609–625. [Google Scholar] [CrossRef] [Green Version]
- Hou, L.; Lanni, F.; Luby-Phelps, K. Tracer Diffusion in F-Actin and Ficoll Mixtures. Toward a Model for Cytoplasm. Biophys. J. 1990, 58, 31–43. [Google Scholar] [CrossRef] [Green Version]
- Pollard, T.D. Rate Constants for the Reactions of ATP- and ADP-Actin with the Ends of Actin Filaments. J. Cell Biol. 1986, 103, 2747–2754. [Google Scholar] [CrossRef] [Green Version]
- Bonet, C.; Ternent, D.; Maciver, S.K.; Mozo-Villarias, A. Rapid Formation and High Diffusibility of Actin-Cofilin Cofilaments at Low pH. Eur. J. Biochem. 2000, 267, 3378–3384. [Google Scholar] [CrossRef]
- Crevenna, A.H.; Naredi-Rainer, N.; Schonichen, A.; Dzubiella, J.; Barber, D.L.; Lamb, D.C.; Wedlich-Soldner, R. Electrostatics Control Actin Filament Nucleation and Elongation Kinetics. J. Biol. Chem. 2013, 288, 12102–12113. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Schwarz, W.H.; Kas, J.A.; Stossel, T.P.; Janmey, P.A.; Pollard, T.D. Mechanical Properties of Actin Filament Networks Depend on Preparation, Polymerization Conditions, and Storage of Actin Monomers. Biophys. J. 1998, 74, 2731–2740. [Google Scholar] [CrossRef] [Green Version]
- Janmey, P.A.; Peetermans, J.; Zaner, K.S.; Stossel, T.P.; Tanaka, T. Structure and Mobility of Actin Filaments as Measured by Quasielastic Light Scattering, Viscometry, and Electron Microscopy. J. Biol. Chem. 1986, 261, 8357–8362. [Google Scholar] [CrossRef]
- Janmey, P.A.; Hvidt, S.; Kas, J.; Lerche, D.; Maggs, A.; Sackmann, E.; Schliwa, M.; Stossel, T.P. The Mechanical Properties of Actin Gels. Elastic Modulus and Filament Motions. J. Biol. Chem. 1994, 269, 32503–32513. [Google Scholar] [CrossRef]
- Kroy, K.; Frey, E. Dynamic Scattering From Solutions of Semiflexible Polymers. Phys. Rev. E 1997, 55, 3092–3101. [Google Scholar] [CrossRef] [Green Version]
- Tassieri, M.; Evans, R.M.L.; Barbu-Tudoran, L.; Trinick, J.; Waigh, T.A. The Self-Assembly, Elasticity, and Dynamics of Cardiac Thin Filaments. Biophys. J. 2008, 94, 2170–2178. [Google Scholar] [CrossRef] [Green Version]
- McDonald, J.H. Handbook of Biological Statistics; Sparky House Publishing: Baltimore, MD, USA, 2009. [Google Scholar]
- Walker, J.M. The Protein Protocols Handbook; Humana Press: Totowa, NJ, USA, 2009. [Google Scholar]
- Simonian, M.H. Spectrophotometric Determination of Protein Concentration. Curr. Protoc. Toxicol. 2004, 21, 1002. [Google Scholar] [CrossRef]
- Mansfield, M.L.; Douglas, J.F. Transport Properties of Rodlike Particles. Macromolecules 2008, 41, 5422–5432. [Google Scholar] [CrossRef]
- Mansfield, M.; Douglas, J.F. Transport Properties of Wormlike Chains with Applications to Double Helical DNA and Carbon Nanotubes. Macromolecules 2008, 41, 5412–5421. [Google Scholar] [CrossRef]
- Volkel, A.R.; Noolandi, J. On the Mobility of Stiff Polyelectrolytes. J. Chem. Phys. 1995, 102, 5506–5511. [Google Scholar] [CrossRef]
- Jeune-Smith, Y.; Hess, H. Engineering the Length Distribution of Microtubules Polymerized In Vitro. Soft Matter. 2010, 6, 1778. [Google Scholar] [CrossRef]
- Yamakawa, H.; Fujii, M. Translational Friction Coefficient of Wormlike Chains. Macromolecules 1973, 6, 407–415. [Google Scholar] [CrossRef]
- Van Der Drift, W.P.J.T.; De Keizer, A.; Overbeek, J.T.G. Electrophoretic Mobility of a Cylinder with High Surface Charge Density. J. Colloid Interface Sci. 1979, 71, 67–78. [Google Scholar] [CrossRef]
- Kirkwood, J.G.; Riseman, J. The Intrinsic Viscosities and Diffusion Constants of Flexible Macromolecules in Solution. J. Chem. Phys. 1948, 16, 565–573. [Google Scholar] [CrossRef]
- Muthukumar, M. Theory of Electrophoretic Mobility of Polyelectrolyte Chains. Macromol. Theory Simul. 1994, 3, 61–71. [Google Scholar] [CrossRef]
- Yeh, I.C.; Hummer, G. Diffusion and Electrophoretic Mobility of Single-Stranded RNA from Molecular Dynamics Simulations. Biophys. J. 2004, 86, 681–689. [Google Scholar] [CrossRef] [Green Version]
- Burlacu, S.; Janmey, P.A.; Borejdo, J. Distribution of Actin Filament Lengths Measured by Fluorescence Microscopy. Am. J. Physiol.-Cell Physiol. 1992, 262, C569–C577. [Google Scholar] [CrossRef]
- Tang, J.X.; Janmey, P.A. The Polyelectrolyte Nature of F-Actin and the Mechanism of Actin Bundle Formation. J. Biol. Chem. 1996, 271, 8556–8563. [Google Scholar] [CrossRef] [Green Version]
- Drenckhahn, D.; Pollard, T.D. Elongation of Actin Filaments is a Diffusion-Limited Reaction at the Barbed End and is Accelerated by Inert Macromolecules. J. Biol. Chem. 1986, 261, 12754–12758. [Google Scholar] [CrossRef]
- Ginell, R.; Simha, R. On the Kinetics of Polymerization Reactions. II. Second and Combined First and Second Order Initiation Reactions. Mutual Stabilization of Growing Chains. J. Am. Chem. Soc. 1943, 65, 715–727. [Google Scholar] [CrossRef]
- Gotter, R.; Kroy, K.; Frey, E.; Barmann, M.; Sackmann, E. Dynamic Light Scattering from Semidilute Actin Solutions: A Study of Hydrodynamic Screening, Filament Bending Stiffness, and the Effect of Tropomyosin/Troponin-Binding. Macromolecules 1996, 29, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Ohshima, H. Approximate Analytic Expression for the Electrophoretic Mobility of Moderately Charged Cylindrical Colloidal Particles. Langmuir 2015, 31, 13633–13638. [Google Scholar] [CrossRef]
- Henry, D.C. The Cataphoresis of Suspended Particles. Part I.—The Equation of Cataphoresis. Proc. R. Soc. Lond. Ser. A Contain. Pap. A Math. Phys. Character 1931, 133, 106–129. [Google Scholar] [CrossRef]
- Vergara-Perez, S.; Marucho, M. MPBEC, a Matlab Program for Biomolecular Electrostatic Calculations. Comput. Phys. Commun. 2016, 198, 179–194. [Google Scholar] [CrossRef] [Green Version]
- Marucho, M. A Java Application to Characterize Biomolecules and Nanomaterials in Electrolyte Aqueous Solutions. Comput. Phys. Commun. 2019, 242, 104–119. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Tan, W.; Lee, W.L. An In Vitro Microscopy-Based Assay for Microtubule-Binding and Microtubule-Crosslinking by Budding Yeast Microtubule-Associated Protein. Bio-Protocol 2018, 8, e3110. [Google Scholar] [CrossRef]
- Patnaik, S.R.; Zhang, X.; Biswas, L.; Akhtar, S.; Zhou, X.; Kusuluri, D.K.; Reilly, J.; May-Simera, H.; Chalmers, S.; McCarron, J.G.; et al. RPGR Protein Complex Regulates Proteasome Activity and Mediates Store-Operated Calcium Entry. Oncotarget 2018, 9, 23183–23197. [Google Scholar] [CrossRef] [Green Version]
- Taylor, J.R. Introduction to Error Analysis: The Study of Uncertainties in Physical Measurements; University Science Books: Melville, New York, NY, USA, 1997. [Google Scholar]
- Tang, J.X.; Janmey, P.A.; Stossel, T.P.; Ito, T. Thiol Oxidation of Actin Produces Dimers That Enhance the Elasticity of the F-Actin Network. Biophys. J. 1999, 76, 2208–2215. [Google Scholar] [CrossRef] [Green Version]
- Pantaloni, D.; Hill, T.L.; Carlier, M.F.; Korn, E.D. A Model for Actin Polymerization and The Kinetic Effects of ATP Hydrolysis. Proc. Natl. Acad. Sci. USA 1985, 82, 7207–7211. [Google Scholar] [CrossRef] [Green Version]
- Tassieri, M. Dynamics of Semiflexible Polymer Solutions in the Tightly Entangled Concentration Regime. Macromolecules 2017, 50, 5611–5618. [Google Scholar] [CrossRef] [Green Version]
- Cong, Y.; Topf, M.; Sali, A.; Matsudaira, P.; Dougherty, M.; Chiu, W.; Schmid, M.F. Crystallographic Conformers of Actin in a Biologically Active Bundle of Filaments. J. Mol. Biol. 2008, 375, 331–336. [Google Scholar] [CrossRef] [Green Version]
- Stigter, D. Interactions of Highly Charged Colloidal Cylinders with Applications to Double-Stranded DNA. Biopolymers 1977, 16, 1435–1448. [Google Scholar] [CrossRef]
- Medasani, B.; Ovanesyan, Z.; Thomas, D.G.; Sushko, M.L.; Marucho, M. Ionic Asymmetry and Solvent Excluded Volume Effects on Spherical Electric Double Layers: A Density Functional Approach. J. Chem. Phys. 2014, 140, 204510. [Google Scholar] [CrossRef] [Green Version]
- Ovanesyan, Z.; Aljzmi, A.; Almusaynid, M.; Khan, A.; Valderrama, E.; Nash, K.L.; Marucho, M. Ion-Ion Correlation, Solvent Excluded Volume and pH Effects on Physicochemical Properties of Spherical Oxide Nanoparticles. J. Colloid Interface Sci. 2016, 462, 325–333. [Google Scholar] [CrossRef] [Green Version]
- Hunley, C.; Marucho, M. Electrical Double Layer Properties of Spherical Oxide Nanoparticles. Phys. Chem. Chem. Phys. 2017, 19, 5396–5404. [Google Scholar] [CrossRef] [Green Version]
- Haviv, L.; Gov, N.; Ideses, Y.; Bernheim-Groswasser, A. Thickness Distribution of Actin Bundles in Vitro. Eur. Biophys. J. 2007, 37, 447–454. [Google Scholar] [CrossRef]
- Arii, Y.; Hatori, K. Relationship Between the Flexibility and The Motility of Actin Filaments: Effects of pH. Biochem. Biophys. Res. Commun. 2008, 371, 772–776. [Google Scholar] [CrossRef]
- Edwards, S.F. The Statistical Mechanics of Polymerized Material. Proc. Phys. Soc. 1967, 92, 9–16. [Google Scholar] [CrossRef]
- de Gennes, P.G. Reptation of a Polymer Chain in the Presence of Fixed Obstacles. J. Chem. Phys. 1971, 55, 572–579. [Google Scholar] [CrossRef]
- Zimmerle, C.T.; Frieden, C. Effect of pH on the Mechanism of Actin Polymerization. Biochemistry 1988, 27, 7766–7772. [Google Scholar] [CrossRef]
- Takatsuki, H.; Chilakamarri, R.; Famouri, P.; Kohama, K. Electrophoretic Mobility of Nano-sized Actin Filaments in Biomolecular Device. In Proceedings of the Sixth IEEE Conference on Nanotechnology, Cincinnati, OH, USA, 17–20 July 2006; Volume 1, pp. 166–169. [Google Scholar] [CrossRef]
- Li, G.; Wen, Q.; Tang, J.X. Single Filament Electrophoresis of F-Actin and Filamentous Virus fd. J. Chem. Phys. 2005, 122, 104708. [Google Scholar] [CrossRef]
- Kamble, S.; Agrawal, S.; Cherumukkil, S.; Sharma, V.; Jasra, R.V.; Munshi, P. Revisiting Zeta Potential, the Key Feature of Interfacial Phenomena, with Applications and Recent Advancements. ChemistrySelect 2022, 7, e202103084. [Google Scholar] [CrossRef]
- Steinmetz, M.O.; Goldie, K.N.; Aebi, U. A Correlative Analysis of Actin Filament Assembly, Structure, and Dynamics. J. Cell Biol. 1997, 138, 559–574. [Google Scholar] [CrossRef] [Green Version]
- Cruz, E.M.D.L.; Mandinova, A.; Steinmetz, M.O.; Stoffler, D.; Aebi, U.; Pollard, T.D. Polymerization and Structure of Nucleotide-free Actin Filaments. J. Mol. Biol. 2000, 295, 517–526. [Google Scholar] [CrossRef] [Green Version]
- Chang, R. Physical Chemistry for the Biosciences; University Science Books: Melville, New York, NY, USA, 2005. [Google Scholar]
- Liu, X.; Pollack, G.H. Mechanics of F-Actin Characterized with Microfabricated Cantilevers. Biophys. J. 2002, 83, 2705–2715. [Google Scholar] [CrossRef] [Green Version]
- Palmer, J.S.; Boyce, M.C. Constitutive Modeling of the Stress-Strain Behavior of F-Actin Filament Networks. Acta Biomater. 2008, 4, 597–612. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Buffer 1 | Buffer 2 | Buffer 3 | |
|---|---|---|---|
| Tris Base | 2 mM | 2 mM | 2 mM |
| CaCl | 0.2 mM | 0.2 mM | 0.2 mM |
| BME (Beta-Mercaptoethanol) | - | 0.1 mM | - |
| ATP (Adenosine triphosphate) | 0.5 mM | 0.2 mM | 0.5 mM |
| DTT (1.4-Dithiothreitol) | 0.2 mM | - | 0.5 mM |
| pH | 7.80 | 7.66 | 8.23 |
| Ionic Strength | 0.0057 | 0.0037 | 0.006 |
| Buffer 1 | Buffer 2 | Buffer 3 | |
|---|---|---|---|
| KCl | 150 mM | 150 mM | 50 mM |
| MgCl | 2 mM | 2 mM | 2 mM |
| pH | 7.56 | 7.64 | 8.07 |
| Ionic strength | 0.155 | 0.155 | 0.055 |
| Buffer 1 | Buffer 2 | Buffer 3 | |
|---|---|---|---|
| KCl | 0.1 M | 0.1 M | 0.1 M |
| pH | 7.72 | 7.66 | 8.06 |
| Ionic strength | 0.1 | 0.1 | 0.1 |
| Buffer 1 | Buffer 2 | Buffer 3 | ||||
|---|---|---|---|---|---|---|
| pH | 7.72 | 7.66 | 8.06 | |||
| x | x | x | x | x | x | |
| D | ||||||
| Buffer 1 | Buffer 2 | Buffer 3 | ||||
|---|---|---|---|---|---|---|
| pH | 7.72 | 7.66 | 8.06 | |||
| x | x | x | x | x | x | |
| (mV) | ||||||
| Buffer | [1/s] | d (nm) | [m] |
|---|---|---|---|
| 1 | |||
| 2 | |||
| 3 |
| Buffer 1 | Buffer 2 | Buffer 3 | |
|---|---|---|---|
| 0.05767 | 0.05291 | 0.07348 | |
| Buffer 1 | Buffer 2 | Buffer 3 | |
|---|---|---|---|
| Buffer 1 | Buffer 2 | Buffer 3 | |
|---|---|---|---|
| Radius of Gyration, | |||
| Polarizability, | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alva, E.; George, A.; Brancaleon, L.; Marucho, M. Hydrodynamic and Polyelectrolyte Properties of Actin Filaments: Theory and Experiments. Polymers 2022, 14, 2438. https://doi.org/10.3390/polym14122438
Alva E, George A, Brancaleon L, Marucho M. Hydrodynamic and Polyelectrolyte Properties of Actin Filaments: Theory and Experiments. Polymers. 2022; 14(12):2438. https://doi.org/10.3390/polym14122438
Chicago/Turabian StyleAlva, Ernesto, Annitta George, Lorenzo Brancaleon, and Marcelo Marucho. 2022. "Hydrodynamic and Polyelectrolyte Properties of Actin Filaments: Theory and Experiments" Polymers 14, no. 12: 2438. https://doi.org/10.3390/polym14122438