Expression and Characterization of a Dye-Decolorizing Peroxidase from Pseudomonas Fluorescens Pf0-1


Abstract
:1. Introduction
2. Results
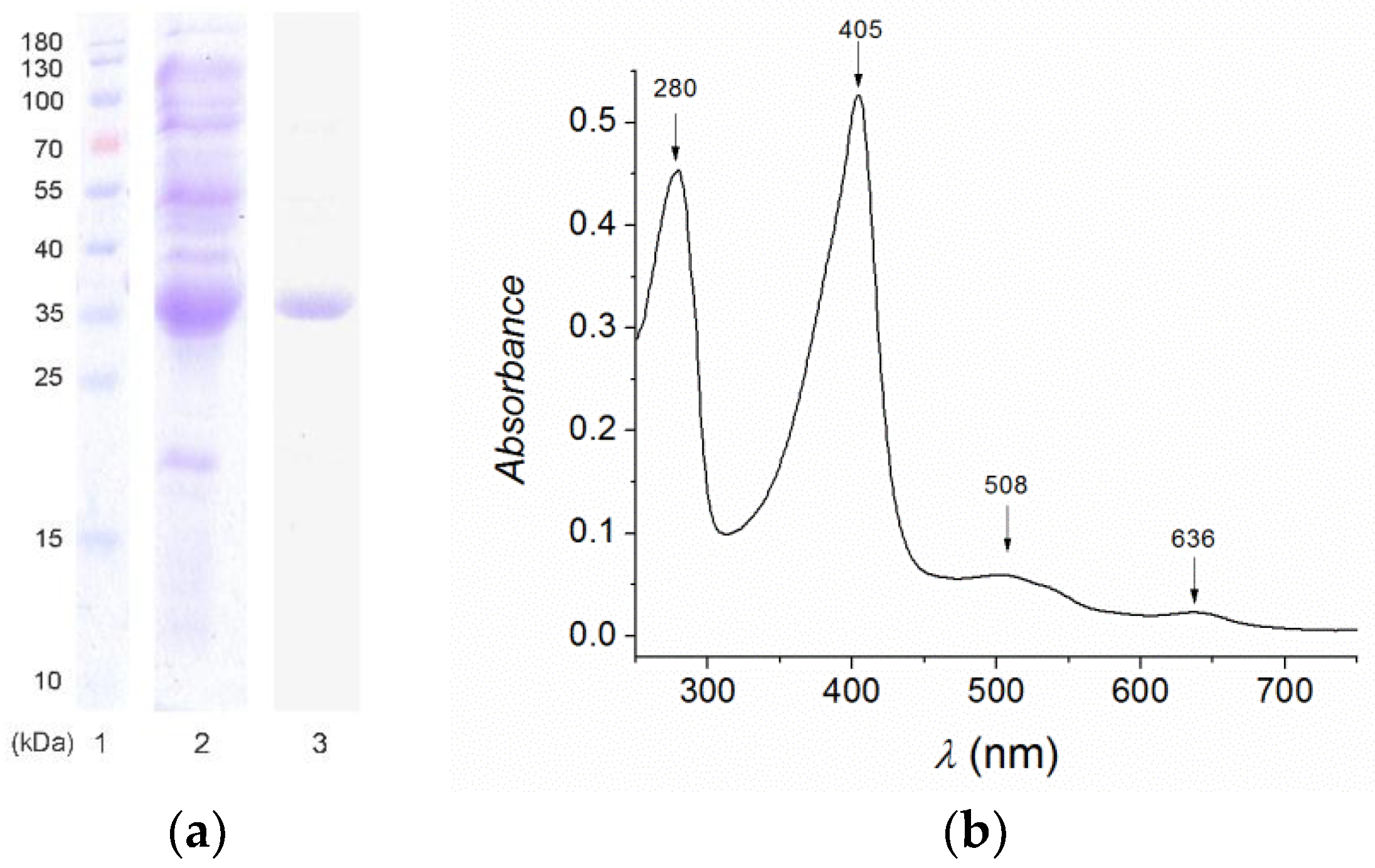
2.1. Expression and Purification
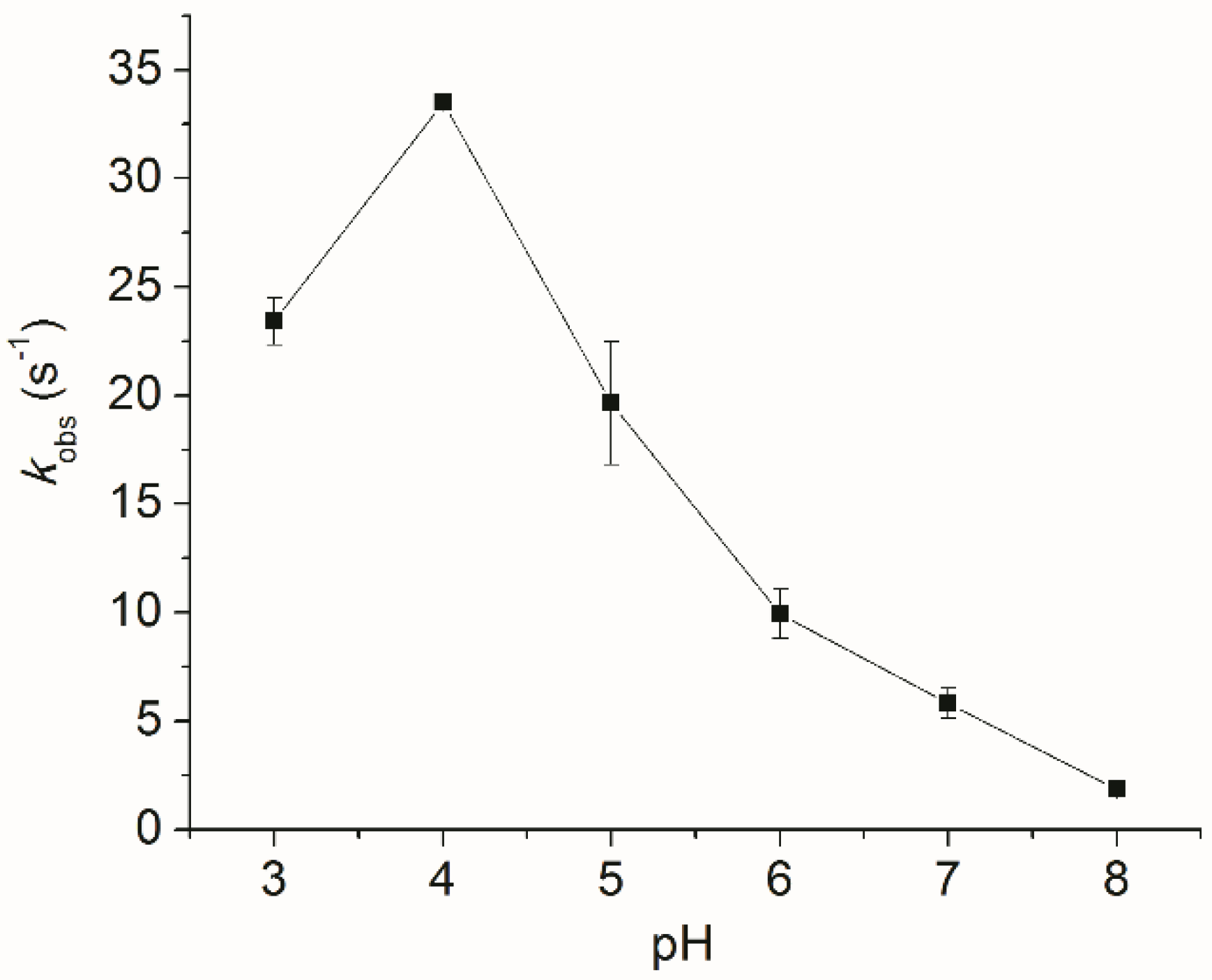
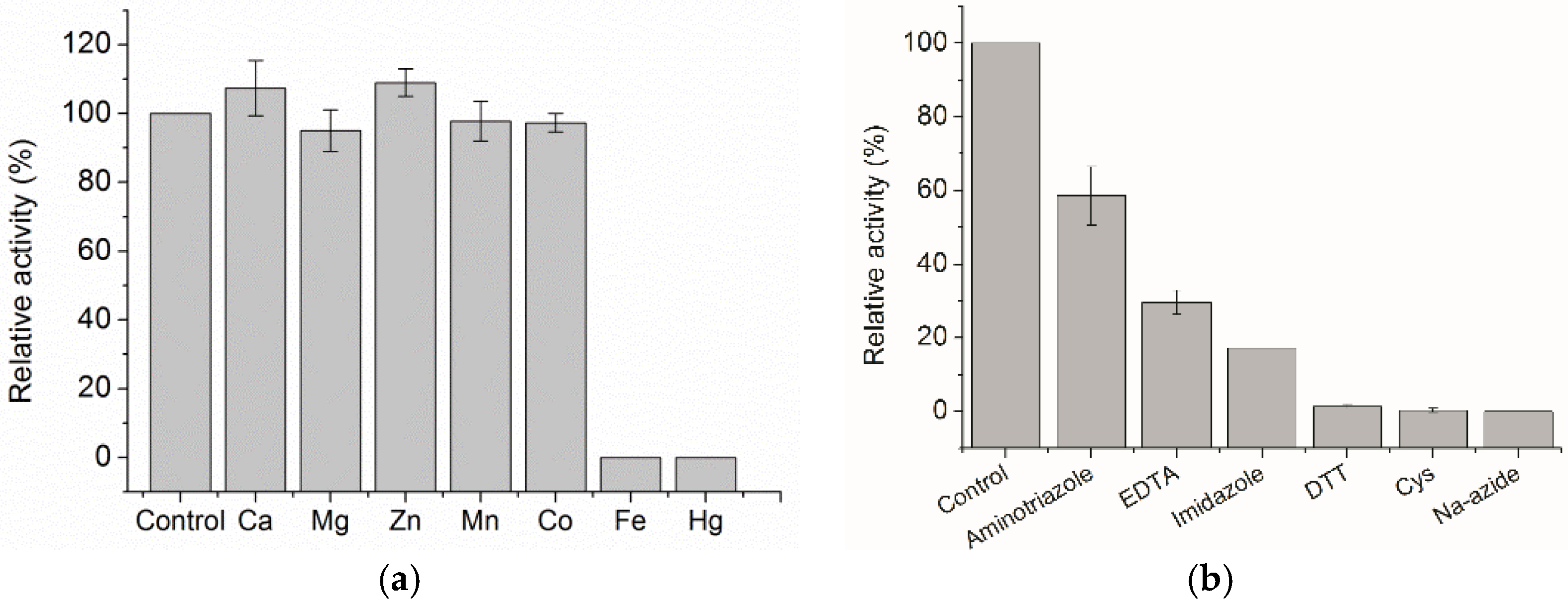
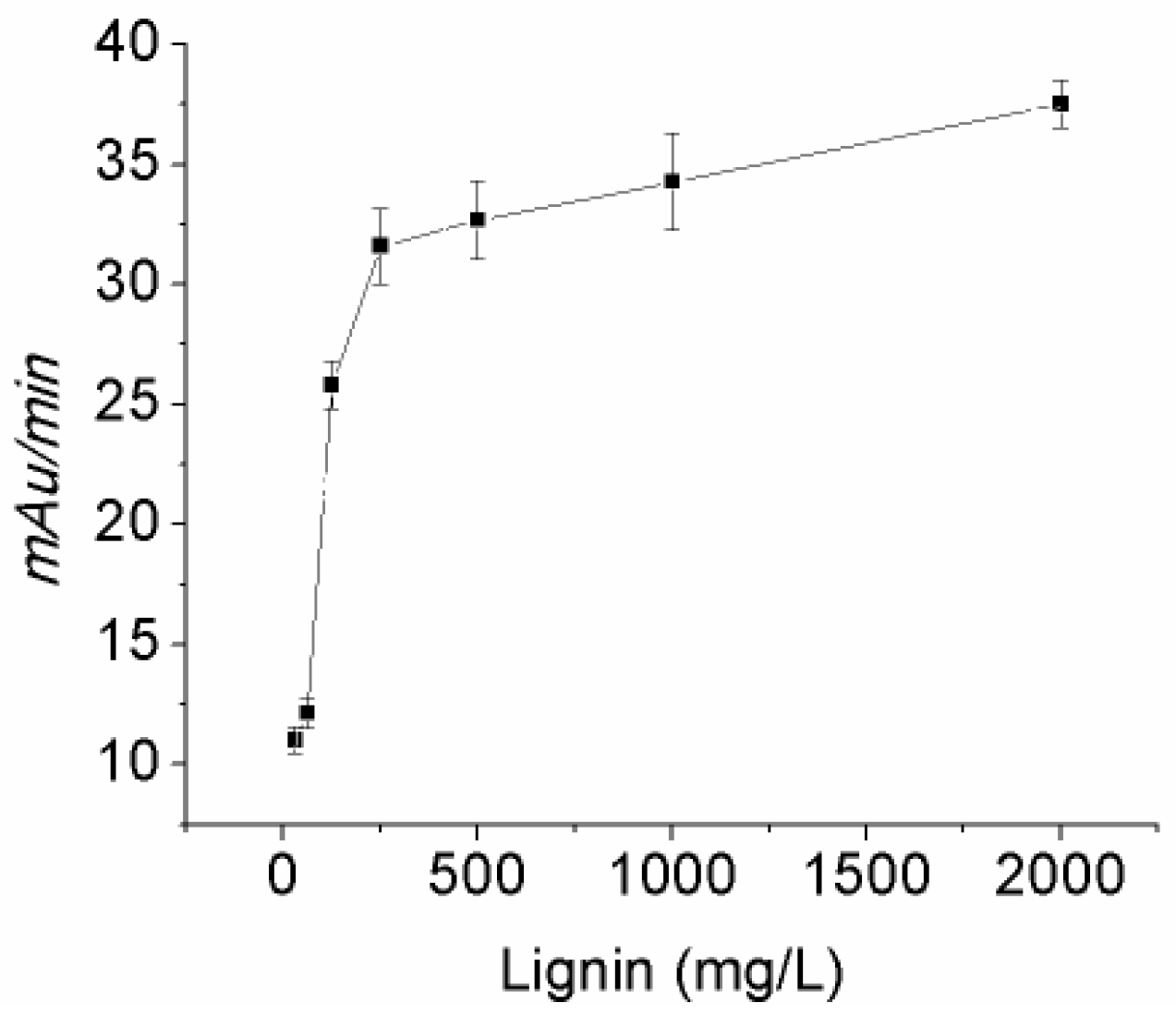
2.2. Biochemical Characterization
3. Discussion
4. Materials and Methods
4.1. Reagents and Enzymes
4.2. Strains, Plasmids, and Growth Conditions
4.3. Sequence Analysis
4.4. Expression and Purification
4.5. Steady-State Kinetic Analyses
4.6. Immobilization Study
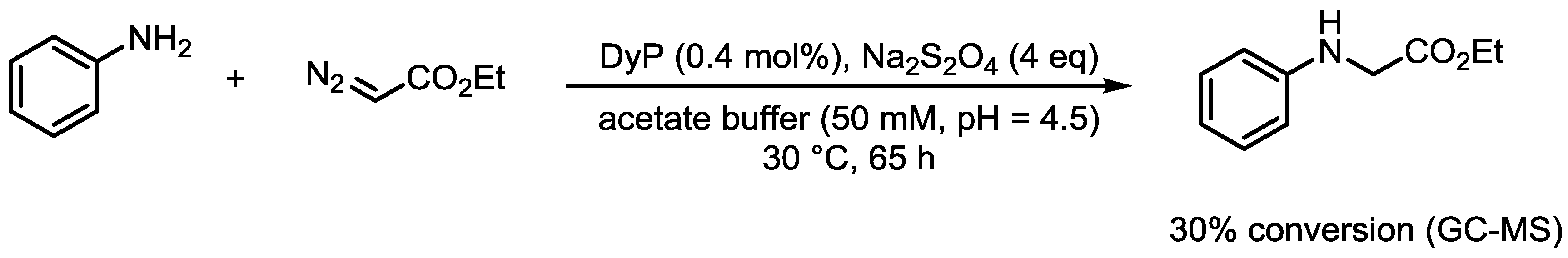
4.7. Procedure for the DyP-Catalyzed Reaction Between Aniline and Ethyl Diazoacetate
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Fraaije, M.W.; Bloois, E.V. DyP-type peroxidases: A promising and versatile class of enzymes. Enzym. Eng. 2012, 1, 1–3. [Google Scholar] [CrossRef]
- Colpa, D.I.; Fraaije, M.W.; van Bloois, E. DyP-type peroxidases: A promising and versatile class of enzymes. J. Ind. Microbiol. Biotechnol. 2014, 41, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lončar, N.; Colpa, D.I.; Fraaije, M.W. Exploring the biocatalytic potential of a DyP-type peroxidase by profiling the substrate acceptance of Thermobifida fusca DyP peroxidase. Tetrahedron 2016, 72, 7276–7281. [Google Scholar] [CrossRef]
- Liers, C.; Bobeth, C.; Pecyna, M.; Ullrich, R.; Hofrichter, M. DyP-like peroxidases of the jelly fungus Auricularia auricula-judae oxidize nonphenolic lignin model compounds and high-redox potential dyes. Appl. Microbiol. Biotechnol. 2010, 85, 1869–1879. [Google Scholar] [CrossRef]
- Linde, D.; Ruiz-Duenas, F.J.; Fernandez-Fueyo, E.; Guallar, V.; Hammel, K.E.; Pogni, R.; Martinez, A.T. Basidiomycete DyPs: Genomic diversity, structural-functional aspects, reaction mechanism and environmental significance. Arch. Biochem. Biophys. 2015, 574, 66–74. [Google Scholar] [CrossRef]
- Salvachua, D.; Prieto, A.; Martinez, A.T.; Martinez, M.J. Characterization of a novel dye-decolorizing peroxidase (DyP)-type enzyme from Irpex lacteus and its application in enzymatic hydrolysis of wheat straw. Appl. Environ. Microbiol. 2013, 79, 4316–4324. [Google Scholar] [CrossRef] [PubMed]
- Scheibner, M.; Hulsdau, B.; Zelena, K.; Nimtz, M.; de Boer, L.; Berger, R.G.; Zorn, H. Novel peroxidases of Marasmius scorodonius degrade beta-carotene. Appl. Microbiol. Biotechnol. 2008, 77, 1241–1250. [Google Scholar] [CrossRef]
- van Bloois, E.; Torres Pazmino, D.E.; Winter, R.T.; Fraaije, M.W. A robust and extracellular heme-containing peroxidase from Thermobifida fusca as prototype of a bacterial peroxidase superfamily. Appl. Microbiol. Biotechnol. 2010, 86, 1419–1430. [Google Scholar] [CrossRef]
- Habib, M.H.; Rozeboom, H.J.; Fraaije, M.W. Characterization of a new DyP-Peroxidase from the alkaliphilic cellulomonad, Cellulomonas bogoriensis. Molecules 2019, 24. [Google Scholar] [CrossRef]
- Colpa, D.I.; Lončar, N.; Schmidt, M.; Fraaije, M.W. Creating Oxidase-Peroxidase Fusion Enzymes as a Toolbox for Cascade Reactions. ChemBioChem 2017, 18, 2226–2230. [Google Scholar] [CrossRef]
- Habib, M.H.M.; Deuss, P.J.; Lončar, N.; Trajkovic, M.; Fraaije, M.W. A biocatalytic one-pot approach for the preparation of lignin oligomers using an oxidase/peroxidase cascade enzyme system. Adv. Synth. Catal. 2017, 359, 3354–3361. [Google Scholar] [CrossRef]
- Yu, W.; Liu, W.; Huang, H.; Zheng, F.; Wang, X.; Wu, Y.; Li, K.; Xie, X.; Jin, Y. Application of a novel alkali-tolerant thermostable DyP-type peroxidase from Saccharomonospora viridis DSM 43017 in biobleaching of eucalyptus kraft pulp. PLoS ONE 2014, 9, e110319. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Guo, D.; Zhou, S.; Yu, X.; Hou, G.; Li, R.; Zhao, B. Cloning and expression of a toxin gene from Pseudomonas fluorescens GcM5-1A. Arch. Microbiol. 2010, 192, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Coelho, P.S.; Brustad, E.M.; Kannan, A.; Arnold, F.H. Olefin cyclopropanation via carbene transfer catalyzed by engineered cytochrome P450 enzymes. Science 2013, 339, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.J.; Peck, N.E.; Renata, H.; Arnold, F.H. Cytochrome P450-catalyzed insertion of carbenoids into N-H bonds. Chem. Sci. 2014, 5, 598–601. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, V.; Fasan, R. Myoglobin-Catalyzed Olefination of Aldehydes. Angew. Chem. Int. Ed. 2016, 55, 2512–2516. [Google Scholar] [CrossRef] [PubMed]
- Sreenilayam, G.; Fasan, R. Myoglobin-catalyzed intermolecular carbene N-H insertion with arylamine substrates. Chem. Commun. 2015, 51, 1532–1534. [Google Scholar] [CrossRef] [Green Version]
- Weissenborn, M.J.; Löw, S.A.; Borlinghaus, N.; Kuhn, M.; Kummer, S.; Rami, F.; Plietker, B.; Hauer, B. Enzyme-catalyzed carbonyl olefination by the E. coli protein YfeX in the absence of phosphines. ChemCatChem 2016, 8, 1636–1640. [Google Scholar] [CrossRef]
- Letoffe, S.; Heuck, G.; Delepelaire, P.; Lange, N.; Wandersman, C. Bacteria capture iron from heme by keeping tetrapyrrol skeleton intact. Proc. Natl. Acad. Sci. USA 2009, 106, 11719–11724. [Google Scholar] [CrossRef] [Green Version]
- Dailey, H.A.; Septer, A.N.; Daugherty, L.; Thames, D.; Gerdes, S.; Stabb, E.V.; Dunn, A.K.; Dailey, T.A.; Phillips, J.D. The Escherichia coli protein YfeX functions as a porphyrinogen oxidase, not a heme dechelatase. MBio 2011, 2, e00248-e11. [Google Scholar] [CrossRef]
- Liu, X.; Yuan, Z.; Wang, J.; Cui, Y.; Liu, S.; Ma, Y.; Gu, L.; Xu, S. Crystal structure and biochemical features of dye-decolorizing peroxidase YfeX from Escherichia coli O157 Asp(143) and Arg(232) play divergent roles toward different substrates. Biochem. Biophys. Res. Commun. 2017, 484, 40–44. [Google Scholar] [CrossRef]
- Hock, K.J.; Knorrscheidt, A.; Hommelsheim, R.; Ho, J.; Weissenborn, M.J.; Koenigs, R.M. Tryptamine synthesis by iron porphyrin catalyzed C-H functionalization of indoles with diazoacetonitrile. Angew. Chem. Int. Ed. 2019, 58, 3630–3634. [Google Scholar] [CrossRef]
- Santos, A.; Mendes, S.; Brissos, V.; Martins, L.O. New dye-decolorizing peroxidases from Bacillus subtilis and Pseudomonas putida MET94: Towards biotechnological applications. Appl. Microbiol. Biotechnol. 2014, 98, 2053–2065. [Google Scholar] [CrossRef]
- Yoshida, T.; Sugano, Y. A structural and functional perspective of DyP-type peroxidase family. Arch. Biochem. Biophys. 2015, 574, 49–55. [Google Scholar] [CrossRef]
- Sugano, Y.; Muramatsu, R.; Ichiyanagi, A.; Sato, T.; Shoda, M. DyP, a unique dye-decolorizing peroxidase, represents a novel heme peroxidase family: ASP171 replaces the distal histidine of classical peroxidases. J. Biol. Chem. 2007, 282, 36652–36658. [Google Scholar] [CrossRef]
- Brown, M.E.; Barros, T.; Chang, M.C.Y. Identification and characterization of a multifunctional dye peroxidase from a lignin-reactive bacterium. ACS Chem. Biol. 2012, 7, 2074–2081. [Google Scholar] [CrossRef]
- Ahmad, M.; Roberts, J.N.; Hardiman, E.M.; Singh, R.; Eltis, L.D.; Bugg, T.D. Identification of DypB from Rhodococcus jostii RHA1 as a lignin peroxidase. Biochemistry 2011, 50, 5096–5107. [Google Scholar] [CrossRef]
- Rahmanpour, R.; Bugg, T.D. Characterisation of Dyp-type peroxidases from Pseudomonas fluorescens Pf-5: Oxidation of Mn(II) and polymeric lignin by Dyp1B. Arch. Biochem. Biophys. 2015, 574, 93–98. [Google Scholar] [CrossRef]
- Pesic, M.; Lopez, C.; Lopez-Santin, J.; Alvaro, G. From amino alcohol to aminopolyol: One-pot multienzyme oxidation and aldol addition. Appl. Microbiol. Biotechnol. 2013, 97, 7173–7183. [Google Scholar] [CrossRef]
- de Gonzalo, G.; Colpa, D.I.; Habib, M.H.; Fraaije, M.W. Bacterial enzymes involved in lignin degradation. J. Biotechnol. 2016, 236, 110–119. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Yue, F.; Cui, Y.; Xu, Y.; Shan, Y.; Liu, B.; Zhou, Y.; Lu, X. Biodegradation of lignin by Pseudomonas sp. Q18 and the characterization of a novel bacterial DyP-type peroxidase. J. Ind. Microbiol. Biotechnol. 2018, 45, 913–927. [Google Scholar] [CrossRef] [PubMed]
- Habib, M.; Trajkovic, M.; Fraaije, M.W. The biocatalytic synthesis of syringaresinol from 2,6-dimethoxy-4-allylphenol in one-pot using a tailored oxidase/peroxidase system. ACS Catal. 2018, 8, 5549–5552. [Google Scholar] [CrossRef] [PubMed]
- Wasak, A.; Drozd, R.; Struk, Ł.; Grygorcewicz, B. Entrapment of DyP-type peroxidase from Pseudomonas fluorescens Pf-5 into Ca-alginate magnetic beads. Biotechnol. Appl. Biochem. 2018, 65, 238–245. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | λ (nm) | ε (mM−1cm−1) | Km (mM) | kcat (s−1) |
|---|---|---|---|---|
| ABTS | 414 | 36.6 | 0.22 ± 0.04 | 102 ± 6.2 |
| o-Dianisidine | 460 | 11.3 | 0.003 ± 0.001 | 14.7 ± 1.7 |
| Pyrogallol | 430 | 2.47 | 1.45 ± 0.63 | 1.98 ± 0.03 |
| Reactive blue 4 | 610 | 4.2 | 0.010 ± 0.004 | 1.54 ± 0.13 |
| Reactive black 5 | 597 | 37 | 0.006 ± 0.002 | 0.04 ± 0.01 |
| H2O2 a | 240 | 0.0394 | 0.52 ± 0.12 | 23 ± 0.84 |
| t-BuOOH a | n.a. | n.a. | 415 ± 144 | 97 ± 28 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lončar, N.; Drašković, N.; Božić, N.; Romero, E.; Simić, S.; Opsenica, I.; Vujčić, Z.; Fraaije, M.W. Expression and Characterization of a Dye-Decolorizing Peroxidase from Pseudomonas Fluorescens Pf0-1. Catalysts 2019, 9, 463. https://doi.org/10.3390/catal9050463
Lončar N, Drašković N, Božić N, Romero E, Simić S, Opsenica I, Vujčić Z, Fraaije MW. Expression and Characterization of a Dye-Decolorizing Peroxidase from Pseudomonas Fluorescens Pf0-1. Catalysts. 2019; 9(5):463. https://doi.org/10.3390/catal9050463
Chicago/Turabian StyleLončar, Nikola, Natalija Drašković, Nataša Božić, Elvira Romero, Stefan Simić, Igor Opsenica, Zoran Vujčić, and Marco W. Fraaije. 2019. "Expression and Characterization of a Dye-Decolorizing Peroxidase from Pseudomonas Fluorescens Pf0-1" Catalysts 9, no. 5: 463. https://doi.org/10.3390/catal9050463