High Cell Density Conversion of Hydrolysed Waste Cooking Oil Fatty Acids Into Medium Chain Length Polyhydroxyalkanoate Using Pseudomonas putida KT2440

 , and
, and
Abstract
:1. Introduction
2. Results
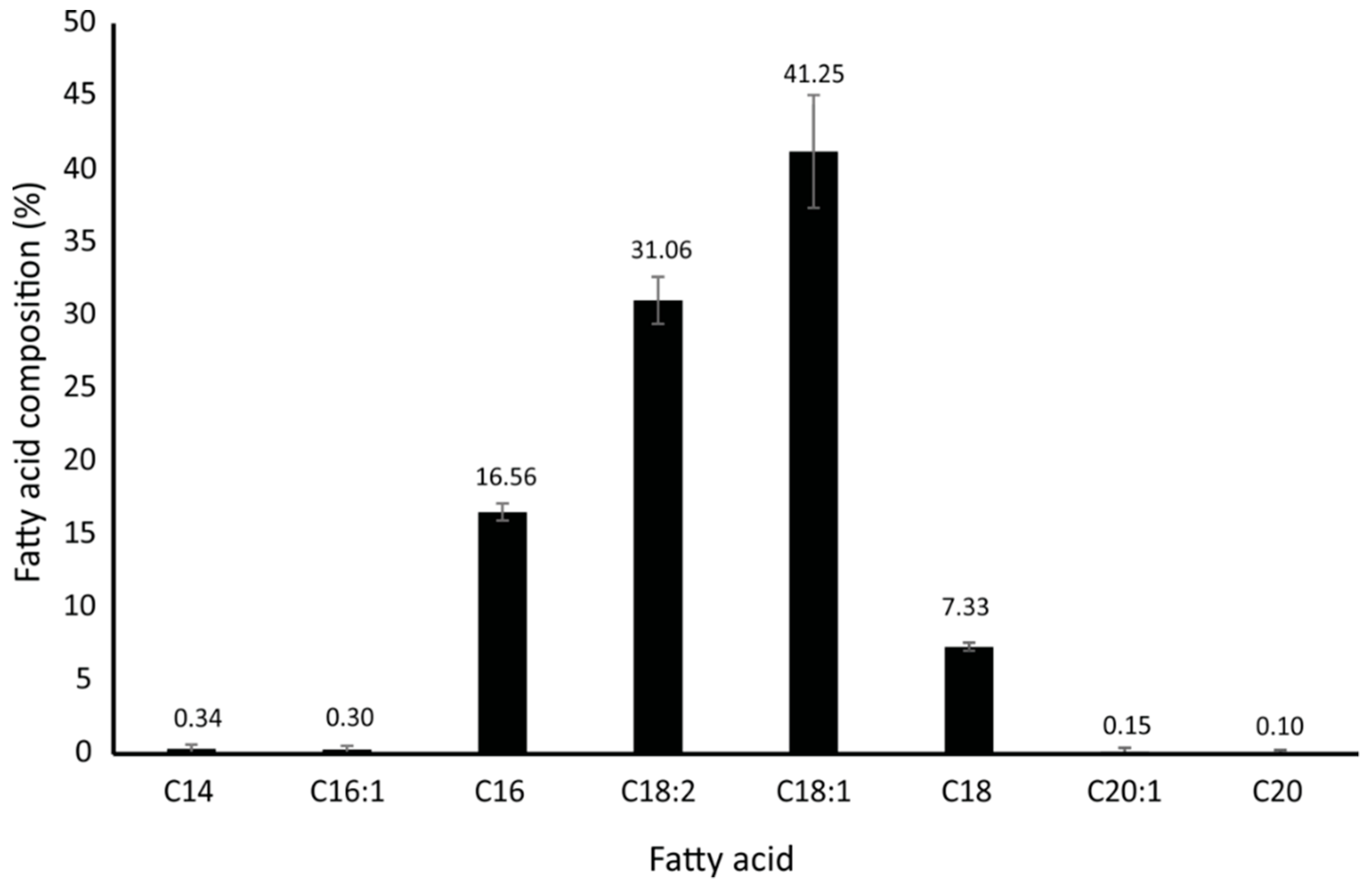
2.1. Fatty Acid Composition of Hydrolysed Waste Cooking Oil (HWCO)
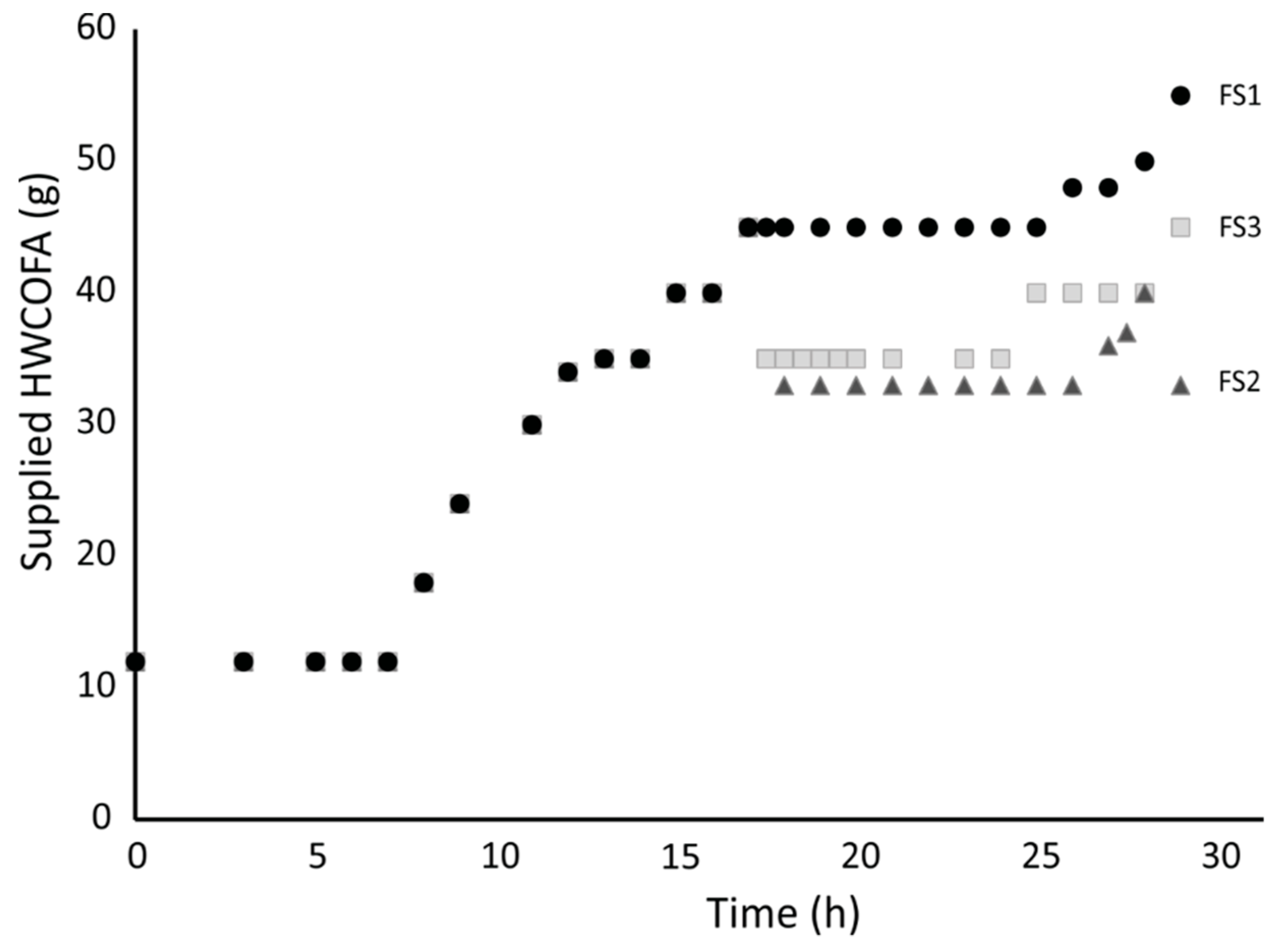
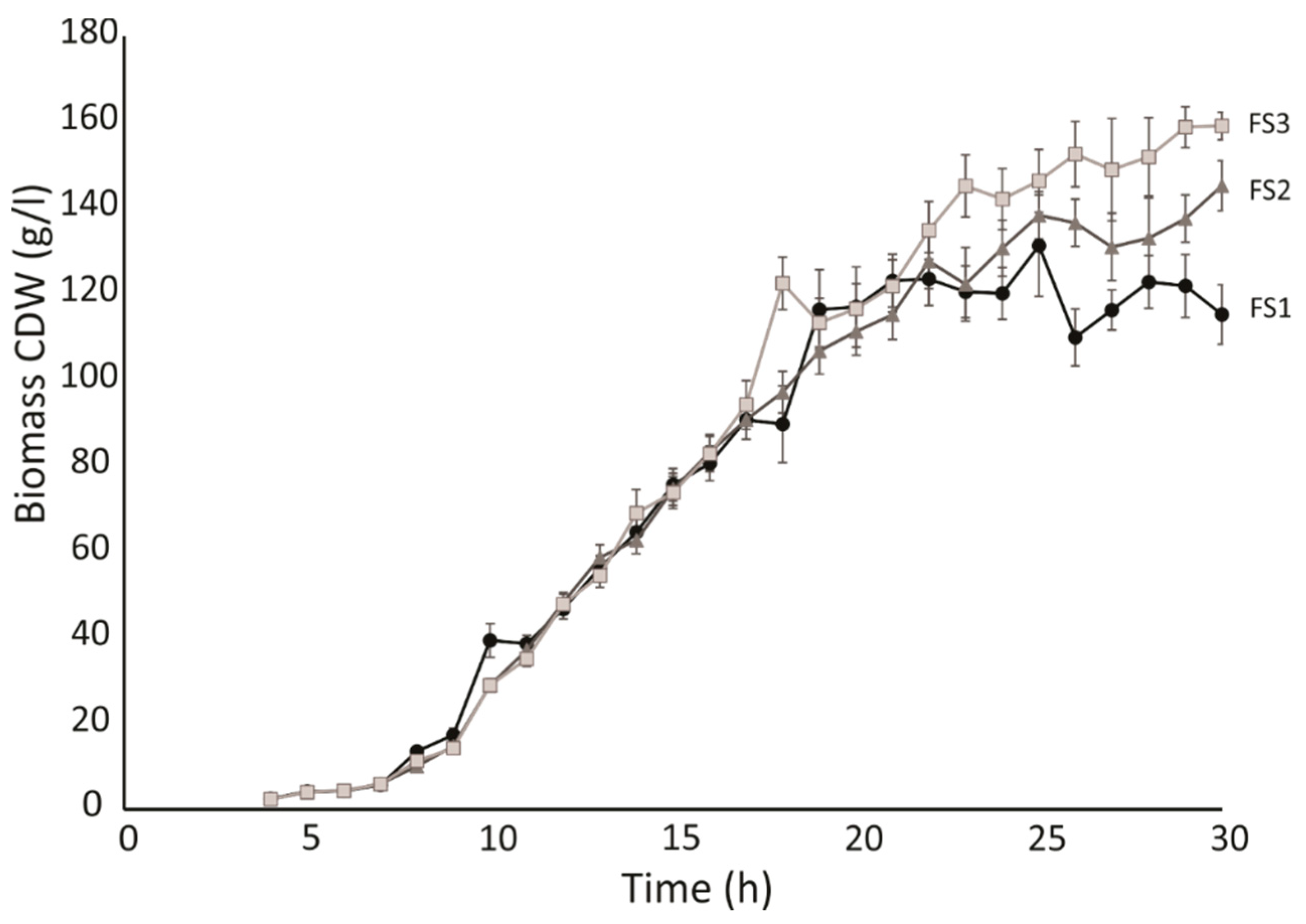
2.2. Bioprocess Development Using HWCOFA Mixture as the Carbon and Energy Source
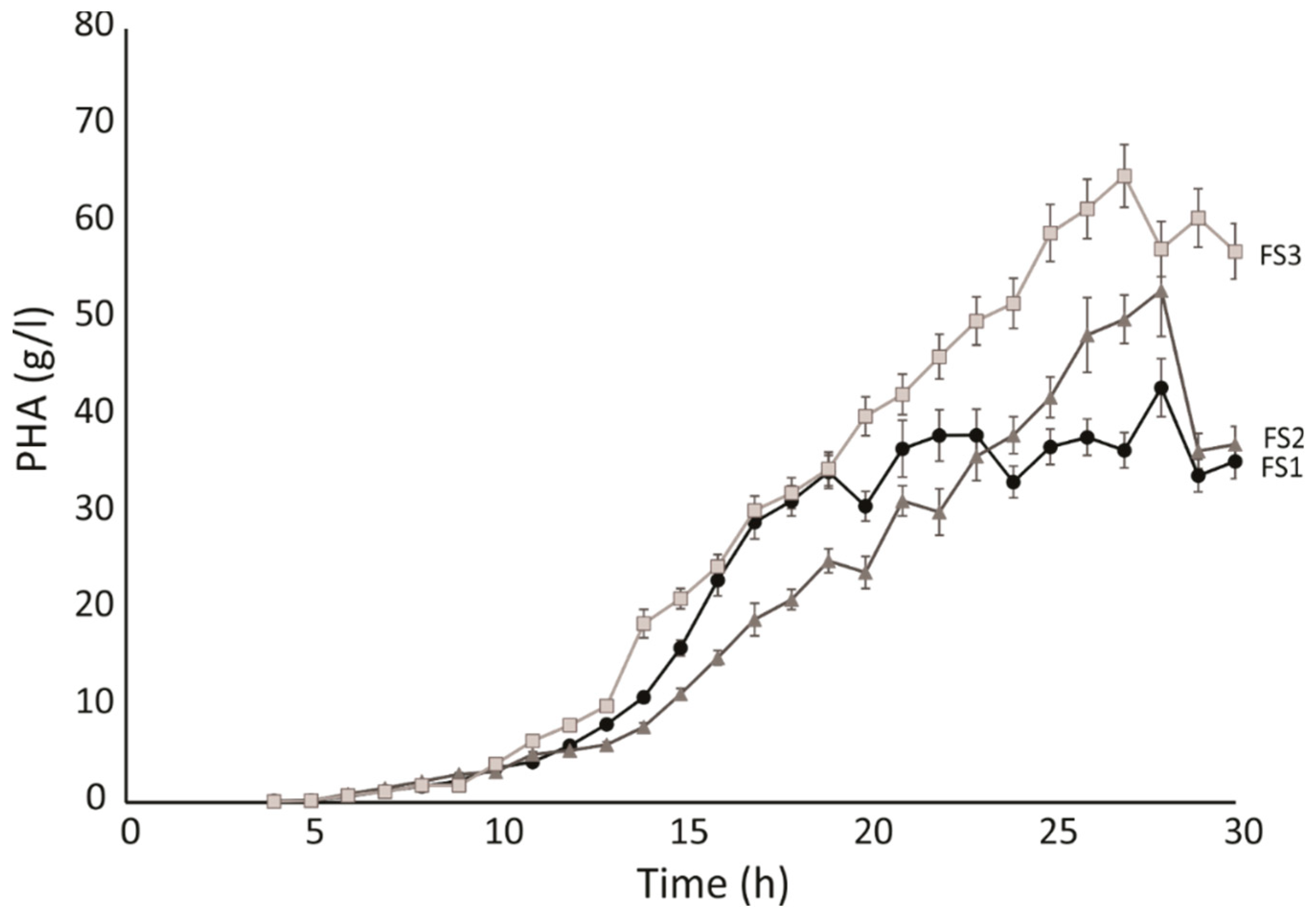
2.3. Polymer Analysis
3. Discussion
4. Materials and Methods
4.1. Bacterial Growth Medium & Strain Maintenance
4.2. Waste Cooking Oil (WCO) Hydrolysis
4.3. Fermentation Conditions
4.4. Analysis of Fatty Acids in Hydrolysed Waste Cooking Oil
4.5. PHA Content and Monomer Composition Determination
4.6. Nutrient and Biomass Analysis
4.7. Polymer Isolation
4.8. PHA Characterisation
4.8.1. Gel Permeation Chromatography (GPC)
4.8.2. Differential Scanning Calorimetry (DSC)
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Keshavarz, T.; Roy, I. Polyhydroxyalkanoates: bioplastics with a green agenda. Curr. Opin. Microbiol. 2010, 13, 321–326. [Google Scholar] [CrossRef]
- Plastics Europe. Plastics—The Facts 2017; 2017. Available online: https://www.plasticseurope.org/en/resources/publications/274-plastics-facts-2017 (accessed on 23 April 2019).
- Narancic, T.; O’Connor, K.E. Plastic waste as a global challenge: are biodegradable plastics the answer to the plastic waste problem. Microbiology 2019, 165, 129–137. [Google Scholar] [CrossRef]
- System Initiative on Environment and Natural Resource Security. The New Plastics Economy: Catalysing action. World Economic Forum: 2017. Available online: https://www.ellenmacarthurfoundation.org/publications/new-plastics-economy-catalysing-action.
- Hanke, G. Marine Beach Litter in Europe—Top Items; Joint Research Centre, European Commission, 2016; Available online: https://mcc.jrc.ec.europa.eu/documents/Marine_Litter/MarineLitterTOPitems_final_24.1.2017.pdf.
- Rochman, C.M.; Browne, M.A.; Halpern, B.S.; Hentschel, B.T.; Hoh, E.; Karapanagioti, H.K.; Rios-Mendoza, L.M.; Takada, H.; Teh, S.; Thompson, R.C. Classify plastic waste as hazardous. Nature 2013, 494, 169–171. [Google Scholar] [CrossRef] [PubMed]
- Rochman, C.M.; Hoh, E.; Kurobe, T.; Teh, S.J. Ingested plastic transfers hazardous chemicals to fish and induces hepatic stress. Sci. Rep. 2013, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, C.; Van Sebille, E.; Hardesty, B.D. Threat of plastic pollution to seabirds is global, pervasive, and increasing. Proc. Natl. Acad. Sci. USA 2015, 112, 11899–11904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mecking, S. Nature or petrochemistry? Biologically degradable materials. Angew. Chem. Int. Ed. 2004, 43, 1078–1085. [Google Scholar] [CrossRef]
- Babu, R.P.; O’Connor, K.; Seeram, R. Current progress on bio-based polymers and their future trends. Prog. Biomater. 2013, 2, 8. [Google Scholar] [CrossRef] [Green Version]
- Mohanty, A.K.; Misra, M.; Drzal, L.T. Sustainable bio-composites from renewable resources: Opportunities and challenges in the green materials world. J. Polym. Environ. 2002, 10, 19–26. [Google Scholar] [CrossRef]
- Harmsen, P.F.H.; Hackmann, M.M.; Bos, H.L. Green building blocks for bio-based plastics. Biofuel Bioprod. Bior. 2014, 8, 306–324. [Google Scholar] [CrossRef]
- Isikgor, F.H.; Becer, C.R. Lignocellulosic biomass: a sustainable platform for the production of bio-based chemicals and polymers. Polym. Chem. 2015, 6, 4497–4559. [Google Scholar] [CrossRef] [Green Version]
- Sudesh, K.; Abe, H.; Doi, Y. Synthesis, structure and properties of polyhydroxyalkanoates: Biological polyesters. Prog. Polym. Sci. 2000, 25, 1503–1555. [Google Scholar] [CrossRef]
- Malacara, C.F.P.; Romero, A.G.; Ponce, M.M.; Marenco, T.C. Approaches for the synthesis of tailor-made polyhydroxyalkanoates. In Microbial Factories: Biodiversity, Biopolymers, Bioactive Molecules: Volume 2; Kalia, V.C., Ed.; Springer India: New Delhi, India; pp. 11–28. [CrossRef]
- Kang, H.O.; Chung, C.W.; Kim, H.W.; Kim, Y.B.; Rhee, Y.H. Cometabolic biosynthesis of copolyesters consisting of 3-hydroxyvalerate and medium-chain-length 3-hydroxyalkanoates by Pseudomonas sp DSY-82. Anton. Leeuw. Int. J. G. 2001, 80, 185–191. [Google Scholar] [CrossRef]
- Lenz, R.W.; Kim, Y.B.; Fuller, R.C. Production of unusual bacterial polyesters by Pseudomonas oleovorans through cometabolism. Fems Microbiol. Lett. 1992, 103, 207–214. [Google Scholar] [CrossRef]
- Ashby, R.D.; Foglia, T.A. Poly(hydroxyalkanoate) biosynthesis from triglyceride substrates. Appl. Microbiol. Biot. 1998, 49, 431–437. [Google Scholar] [CrossRef]
- Li, Z.B.; Loh, X.J. Water soluble polyhydroxyalkanoates: future materials for therapeutic applications. Chem. Soc. Rev. 2015, 44, 2865–2879. [Google Scholar] [CrossRef] [PubMed]
- Vastano, M.; Pellis, A.; Immirzi, B.; Dal Poggetto, G.; Malinconico, M.; Sannia, G.; Guebitz, G.M.; Pezzella, C. Enzymatic production of clickable and PEGylated recombinant polyhydroxyalkanoates. Green. Chem. 2017, 19, 5494–5504. [Google Scholar] [CrossRef]
- Walsh, M.; O’Connor, K.; Babu, R.; Woods, T.; Kenny, S. Plant oils and products of their hydrolysis as substrates for polyhydroxyalkanoate synthesis. Chem. Biochem. Eng. Q. 2015, 29, 123–133. [Google Scholar] [CrossRef]
- Sabapathy, P.C.; Devaraj, S.; Kathirvel, P. Parthenium hysterophorus: low cost substrate for the production of polyhydroxyalkanoates. Curr. Sci. 2017, 112, 2106–2111. [Google Scholar] [CrossRef]
- Poblete-Castro, I.; Binger, D.; Oehlert, R.; Rohde, M. Comparison of mcl-Poly(3-hydroxyalkanoates) synthesis by different Pseudomonas putida strains from crude glycerol: citrate accumulates at high titer under PHA-producing conditions. BMC Biotechnol. 2014, 14. [Google Scholar] [CrossRef]
- Ward, P.G.; Goff, M.; Donner, M.; Kaminsky, W.; O’Connor, K.E. A two step chemo-biotechnological conversion of polystyrene to a biodegradable thermoplastic. Environ. Sci. Technol. 2006, 40, 2433–2437. [Google Scholar] [CrossRef]
- Maddikeri, G.L.; Pandit, A.B.; Gogate, P.R. Intensification approaches for biodiesel synthesis from waste cooking oil: A review. Ind. Eng. Chem. Res. 2012, 51, 14610–14628. [Google Scholar] [CrossRef]
- Sanchez, R.J.; Schripsema, J.; da Silva, L.F.; Taciro, M.K.; Pradella, J.G.C.; Gomez, J.G.C. Medium-chain-length polyhydroxyalkanoic acids (PHA(mcl)) produced by Pseudomonas putida IPT 046 from renewable sources. Eur. Polym. J. 2003, 39, 1385–1394. [Google Scholar] [CrossRef]
- Le Meur, S.; Zinn, M.; Egli, T.; Thony-Meyer, L.; Ren, Q. Production of medium-chain-length polyhydroxyalkanoates by sequential feeding of xylose and octanoic acid in engineered Pseudomonas putida KT2440. BMC Biotechnol. 2012, 12, 1–12. [Google Scholar] [CrossRef]
- Sun, Z.; Ramsay, J.A.; Guay, M.; Ramsay, B. Increasing the yield of MCL-PHA from nonanoic acid by co-feeding glucose during the PHA accumulation stage in two-stage fed-batch fermentations of Pseudomonas putida KT2440. J. Biotechnol. 2007, 132, 280–282. [Google Scholar] [CrossRef]
- Kim, B.S. Production of medium chain length polyhydroxyalkanoates by fed-batch culture of Pseudomonas oleovorans. Biotechnol. Lett. 2002, 24, 125–130. [Google Scholar] [CrossRef]
- Kellerhals, M.B.; Kessler, B.; Witholt, B.; Tchouboukov, A.; Brandl, H. Renewable long-chain fatty acids for production of biodegradable medium-chain-length polyhydroxyalkanoates (mcl-PHAs) at laboratory and pilot plant scales. Macromolecules 2000, 33, 4690–4698. [Google Scholar] [CrossRef]
- Annuar, M.S.M.; Tan, I.K.P.; Ibrahim, S.; Ramachandran, K.B. Production of medium-chain-length poly(3-hydroxyalkanoates) from crude fatty acids mixture by Pseudomonas putida. Food Bioprod Process 2007, 85, 104–119. [Google Scholar] [CrossRef]
- Fernández, D.; Rodríguez, E.; Bassas, M.; Viñas, M.; Solanas, A.M.; Llorens, J.; Marqués, A.M.; Manresa, A. Agro-industrial oily wastes as substrates for PHA production by the new strain Pseudomonas aeruginosa NCIB 40045: Effect of culture conditions. Biochem. Eng. J. 2005, 26, 159–167. [Google Scholar] [CrossRef]
- Mozejko, J.; Wilke, A.; Przybylek, G.; Ciesielski, S. Mcl-PHAs produced by Pseudomonas sp Gl01 using fed-batch cultivation with waste rapeseed oil as carbon source. J. Microbiol. Biotechn. 2012, 22, 371–377. [Google Scholar] [CrossRef]
- Tan, I.K.P.; Kumar, K.S.; Theanmalar, M.; Gan, S.N.; Gordon, B. Saponified palm kernel oil and its major free fatty acids as carbon substrates for the production of polyhydroxyalkanoates in Pseudomonas putida PGA1. Appl. Microbiol. Biot. 1997, 47, 207–211. [Google Scholar] [CrossRef]
- Belda, E.; van Heck, R.G.A.; Lopez-Sanchez, M.J.; Cruveiller, S.; Barbe, V.; Fraser, C.; Klenk, H.P.; Petersen, J.; Morgat, A.; Nikel, P.I.; et al. The revisited genome of Pseudomonas putida KT2440 enlightens its value as a robust metabolic chassis. Environ. Microbiol. 2016, 18, 3403–3424. [Google Scholar] [CrossRef] [PubMed]
- Leung, D.Y.C.; Guo, Y. Transesterification of neat and used frying oil: Optimization for biodiesel production. Fuel Process Technol. 2006, 87, 883–890. [Google Scholar] [CrossRef]
- Cruz, M.V.; Sarraguca, M.C.; Freitas, F.; Lopes, J.A.; Reis, M.A.M. Online monitoring of P(3HB) produced from used cooking oil with near-infrared spectroscopy. J. Biotechnol. 2015, 194, 1–9. [Google Scholar] [CrossRef]
- Martino, L.; Cruz, M.V.; Scoma, A.; Freitas, F.; Bertin, L.; Scandola, M.; Reis, M.A.M. Recovery of amorphous polyhydroxybutyrate granules from Cupriavidus necator cells grown on used cooking oil. Int. J. Biol. Macromol. 2014, 71, 117–123. [Google Scholar] [CrossRef]
- Maclean, H.; Sun, Z.Y.; Ramsay, J.; Ramsay, B. Decaying exponential feeding of nonanoic acid for the production of medium-chain-length poly(3-hydroxyalkanoates) by Pseudomonas putida KT2440. Can. J. Chem. 2008, 86, 564–569. [Google Scholar] [CrossRef]
- Lemos, P.C.; Serafim, L.S.; Reis, M.A.M. Synthesis of polyhydroxyalkanoates from different short-chain fatty acids by mixed cultures submitted to aerobic dynamic feeding. J. Biotechnol. 2006, 122, 226–238. [Google Scholar] [CrossRef] [PubMed]
- Zhila, N.; Shishatskaya, E. Properties of PHA bi-, ter-, and quarter-polymers containing 4-hydroxybutyrate monomer units. Int. J. Biol. Macromol. 2018, 111, 1019–1026. [Google Scholar] [CrossRef] [Green Version]
- Nantha Gopal, K.; Pal, A.; Sharma, S.; Samanchi, C.; Sathyanarayanan, K.; Elango, T. Investigation of emissions and combustion characteristics of a CI engine fueled with waste cooking oil methyl ester and diesel blends. Alex. Eng. J. 2014, 53, 281–287. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, I.; Kagotani, K.; Kimura, Y. Microbial production of poly(hydroxyalkanoate)s from waste edible oils. Green Chem. 2003, 5, 545–548. [Google Scholar] [CrossRef]
- Giridhar, R.; Srivastava, A.K. Fed-batch sorbose fermentation using pulse and multiple feeding strategies for productivity improvement. Biotechnol. Bioprocess Eng. 2000, 5, 340–344. [Google Scholar] [CrossRef]
- Lie, E.; Molin, G. Hydrolysis and esterification with immobilized lipase on hydrophobic and hydrophilic zeolites. J. Chem. Technol. Biot. 1991, 50, 549–553. [Google Scholar] [CrossRef]
- Lee, S.Y.; Wong, H.H.; Choi, J.I.; Lee, S.H.; Lee, S.C.; Han, C.S. Production of medium-chain-length polyhydroxyalkanoates by high-cell-density cultivation of Pseudomonas putida under phosphorus limitation. Biotechnol. Bioeng. 2000, 68, 466–470. [Google Scholar] [CrossRef]
- Lee, S.H.; Oh, D.H.; Ahn, W.S.; Lee, Y.; Choi, J.I.; Lee, S.Y. Production of poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) by high-cell-density cultivation of Aeromonas hydrophila. Biotechnol. Bioeng. 2000, 67, 240–244. [Google Scholar] [CrossRef]
- Marsudi, S.; Unno, H.; Hori, K. Palm oil utilization for the simultaneous production of polyhydroxyalkanoates and rhamnolipids by Pseudomonas aeruginosa. Appl. Microbiol. Biot. 2008, 78, 955–961. [Google Scholar] [CrossRef] [PubMed]
- Verlinden, R.A.J.; Hill, D.J.; Kenward, M.A.; Williams, C.D.; Piotrowska-Seget, Z.; Radecka, I.K. Production of polyhydroxyalkanoates from waste frying oil by Cupriavidus necator. AMB Exp. 2011, 1, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yun, H.S.; Kim, D.Y.; Chung, C.W.; Kim, H.W.; Yang, K.Y.; Rhee, Y.H. Characterization of a tacky poly(3-hydroxyalkanoate) produced by Pseudomonas chlororaphis HS21 from palm kernel oil. J. Microbiol. Biotechn. 2003, 13, 64–69. [Google Scholar]
- Mozejko, J.; Ciesielski, S. Saponified waste palm oil as an attractive renewable resource for mcl-polyhydroxyalkanoate synthesis. J. Biosci. Bioeng. 2013, 116, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Haba, E.; Vidal-Mas, J.; Bassas, M.; Espuny, M.J.; Llorens, J.; Manresa, A. Poly 3-(hydroxyalkanoates) produced from oily substrates by Pseudomonas aeruginosa 47T2 (NCBIM 40044): Effect of nutrients and incubation temperature on polymer composition. Biochem. Eng. J. 2007, 35, 99–106. [Google Scholar] [CrossRef]
- Cromwick, A.M.; Foglia, T.; Lenz, R.W. The microbial production of poly(hydroxyalkanoates) from tallow. Appl. Microbiol. Biot. 1996, 46, 464–469. [Google Scholar] [CrossRef]
- Tobin, K.M.; O’Connor, K.E. Polyhydroxyalkanoate accumulating diversity of Pseudomonas species utilising aromatic hydrocarbons. Fems Microbiol. Lett. 2005, 253, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Valentino, F.; Karabegouic, L.; Majone, M.; Morgan-Sagastume, F.; Werker, A. Polyhydroxyalkanoate (PHA) storage within a mixed-culture biomass with simultaneous growth as a function of accumulation substrate nitrogen and phosphorus levels. Water Res. 2015, 77, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Muhr, A.; Rechberger, E.M.; Salerno, A.; Reiterer, A.; Malli, K.; Strohmeier, K.; Schober, S.; Mittelbach, M.; Koller, M. Novel description of mcl-PHA biosynthesis by Pseudomonas chlororaphis from animal-derived waste. J. Biotechnol. 2013, 165, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Gumel, A.M.; Annuar, M.S.M.; Heidelberg, T. Growth kinetics, effect of carbon substrate in biosynthesis of mcl-PHA by Pseudomonas putida Bet001. Braz. J. Microbiol. 2014, 45, 427–438. [Google Scholar] [CrossRef] [Green Version]
- Schlegel, H.G.; Kaltwasser, H.; Gottschalk, G. A submersion method for culture of hydrogen-oxidizing bacteria: growth physiological studies. Arch. Mikrobiol. 1961, 38, 209–222. [Google Scholar] [CrossRef]
- Brandl, H.; Gross, R.A.; Lenz, R.W.; Fuller, R.C. Pseudomonas oleovorans as a source of poly(beta-hydroxyalkanoates) for potential applications as biodegradable polyesters. Appl. Environ. Microb. 1988, 54, 1977–1982. [Google Scholar]
- Lageveen, R.G.; Huisman, G.W.; Preusting, H.; Ketelaar, P.; Eggink, G.; Witholt, B. Formation of polyesters by Pseudomonas oleovorans—Effect of substrates on formation and composition of poly-(R)-3-hydroxyalkanoates and poly-(R)-3-hydroxyalkenoates. Appl. Environ. Microb. 1988, 54, 2924–2932. [Google Scholar]
- Scheiner, D. Determination of ammonia and Kjeldahl nitrogen by indophenol method. Water Res. 1976, 10, 31–36. [Google Scholar] [CrossRef]
- Elbahloul, Y.; Steinbuhel, A. Large-scale production of poly(3-hydroxyoctanoic acid) by Pseudomonas putida GPo1 and a simplified downstream process. Appl. Environ. Microb. 2009, 75, 643–651. [Google Scholar] [CrossRef]
- Kenny, S.T.; Runic, J.N.; Kaminsky, W.; Woods, T.; Babu, R.P.; Keely, C.M.; Blau, W.; O’Connor, K.E. Up-Cycling of PET (Polyethylene Terephthalate) to the Biodegradable Plastic PHA (Polyhydroxyalkanoate). Environ. Sci. Technol. 2008, 42, 7696–7701. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Feed Strategy | Initial Volume (l) | Total Substrate Used (g) | Final CDW (g/l) | Final Volume (l) | Total Biomass (g) |
|---|---|---|---|---|---|---|
| HWCOFAs * | 1 | 3 | 967 | 115.1 | 4.2 | 483.4 |
| Final PHA (%CDW) | Final PHA (g/l) | PHA productivity (g/l/h) | Total PHA produced (g) | Biomass Yield (g/g) | PHA Yield (g/g) | |
| 30.6 | 35.2 | 1.17 | 147.9 | 0.50 | 0.15 | |
| HWCOFAs | Feed strategy | Initial Volume (l) | Total substrate used (g) | Final CDW (g/l) | Final Volume (l) | Total Biomass (g) |
| 2 | 3 | 804 | 145.2 | 4.1 | 595.3 | |
| Final PHA (%CDW) | Final PHA (g/l) | PHA productivity (g/l/h) | Total PHA produced (g) | Biomass Yield (g/g) | PHA Yield (g/g) | |
| 25.5 | 37.0 | 1.23 | 151.8 | 0.74 | 0.19 | |
| HWCOFAs | Feed strategy | Initial Volume (l) | Total substrate used (g) | Final CDW (g/l) | Final Volume (l) | Total Biomass (g) |
| 3 | 3 | 881 | 159.4 | 4.2 | 669.5 | |
| Final PHA (%CDW) | Final PHA (g/l) | PHA productivity (g/l/h) | Total PHA produced (g) | Biomass Yield (g/g) | PHA Yield (g/g) | |
| 36.4 | 58.0 | 1.93 | 243.7 | 0.76 | 0.28 |
| Substrate Conditions | Td (°C) | Tg (°C) | Tm (°C) | Mn | Mw |
|---|---|---|---|---|---|
| HWCOFAs | 270.61 ± 1.6 | -56.1 ± 0.5 | 20.7 ± 0.5 | 22954 ± 975 | 45317 ± 62 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruiz, C.; Kenny, S.T.; Babu P, R.; Walsh, M.; Narancic, T.; O’Connor, K.E. High Cell Density Conversion of Hydrolysed Waste Cooking Oil Fatty Acids Into Medium Chain Length Polyhydroxyalkanoate Using Pseudomonas putida KT2440. Catalysts 2019, 9, 468. https://doi.org/10.3390/catal9050468
Ruiz C, Kenny ST, Babu P R, Walsh M, Narancic T, O’Connor KE. High Cell Density Conversion of Hydrolysed Waste Cooking Oil Fatty Acids Into Medium Chain Length Polyhydroxyalkanoate Using Pseudomonas putida KT2440. Catalysts. 2019; 9(5):468. https://doi.org/10.3390/catal9050468
Chicago/Turabian StyleRuiz, Carolina, Shane T. Kenny, Ramesh Babu P, Meg Walsh, Tanja Narancic, and Kevin E. O’Connor. 2019. "High Cell Density Conversion of Hydrolysed Waste Cooking Oil Fatty Acids Into Medium Chain Length Polyhydroxyalkanoate Using Pseudomonas putida KT2440" Catalysts 9, no. 5: 468. https://doi.org/10.3390/catal9050468