
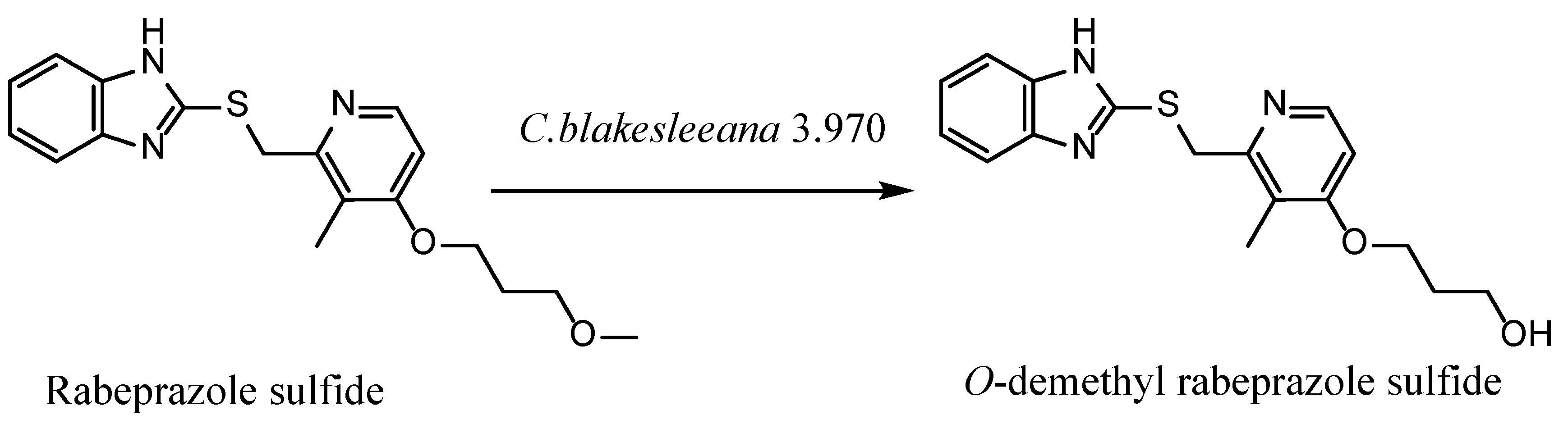
An O-Demethylation Metabolite of Rabeprazole Sulfide by Cunninghamella blakesleeana 3.970 Biotransformation

Abstract
:1. Introduction
2. Results and Discussion
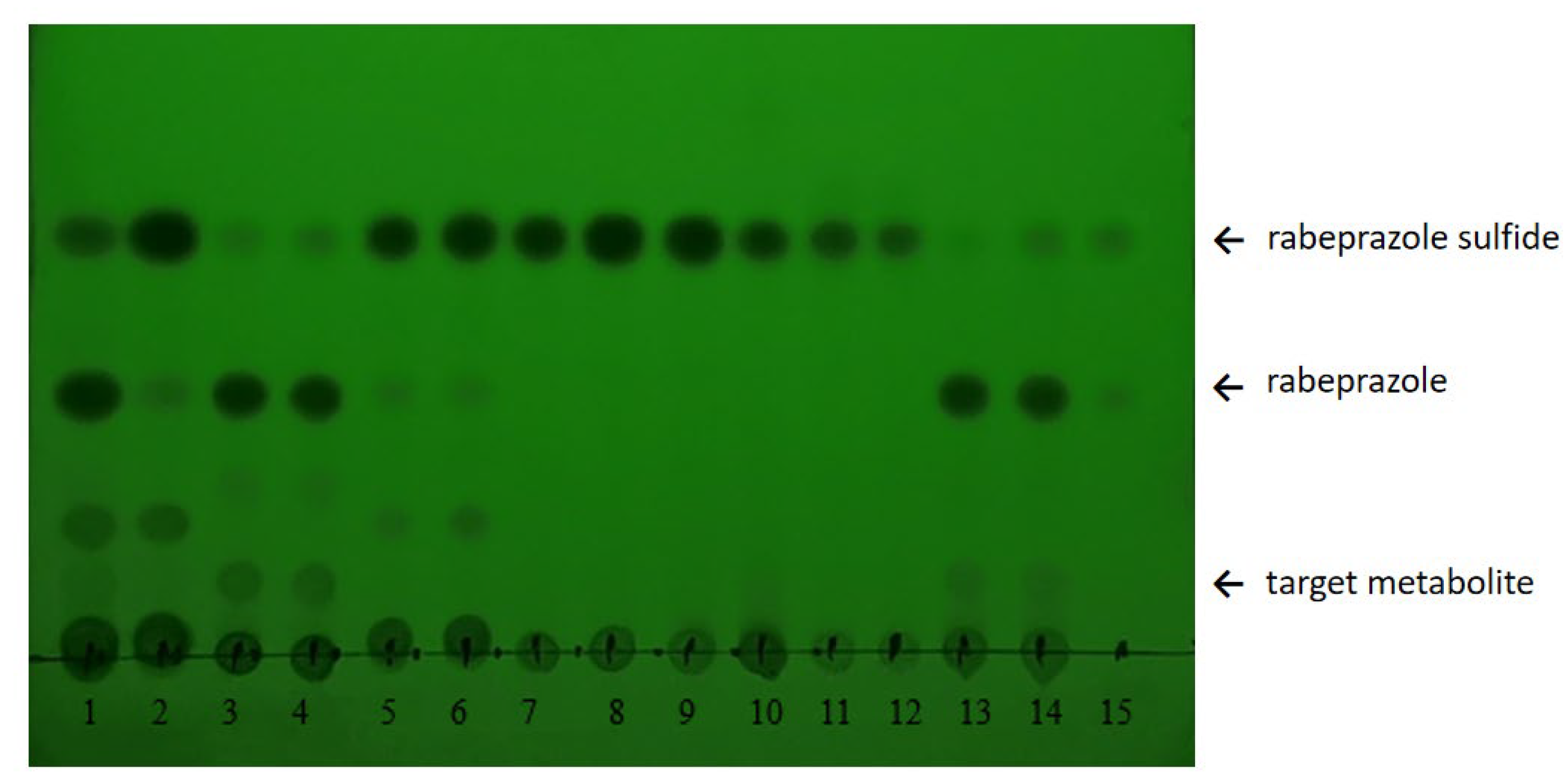
2.1. Whole-Cell Biotransformation Results of Rabeprazole Sulfide
2.2. Optimization of Transformation Media
2.3. Single-Factor Evaluation for Rabeprazole Sulfide Biotransformation
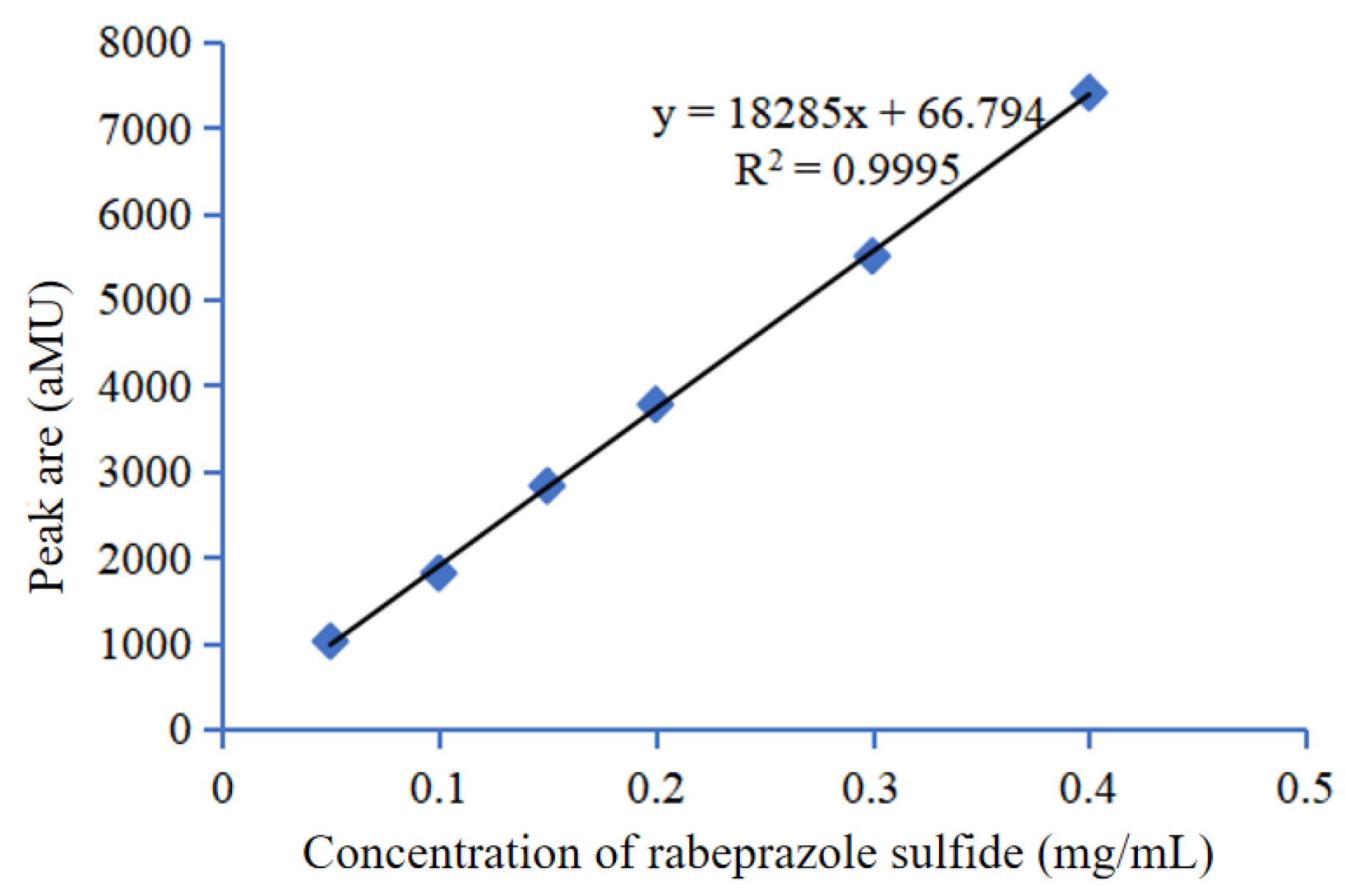
2.3.1. Standard Curve of Rabeprazole Sulfide
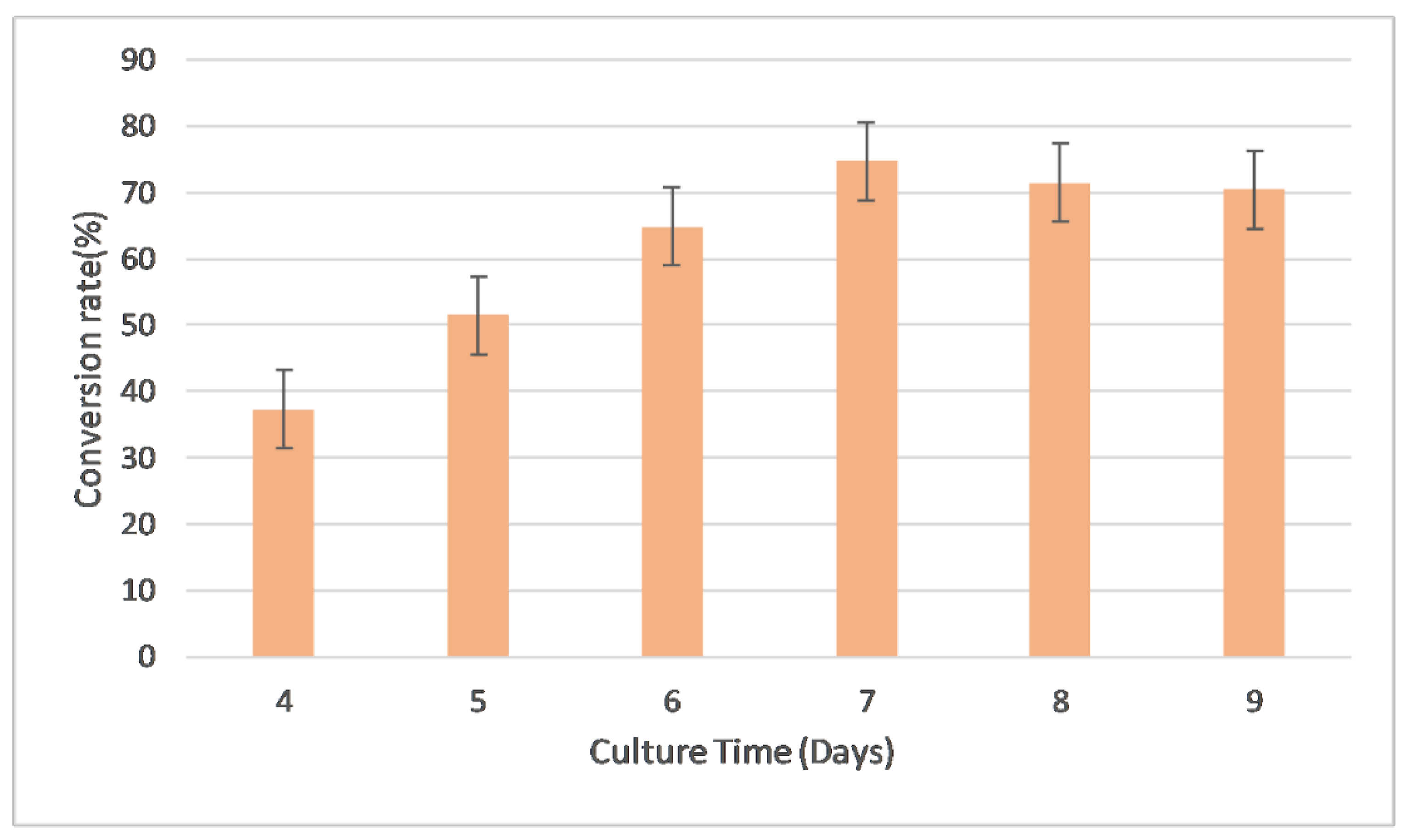
2.3.2. Evaluation of Culture Time
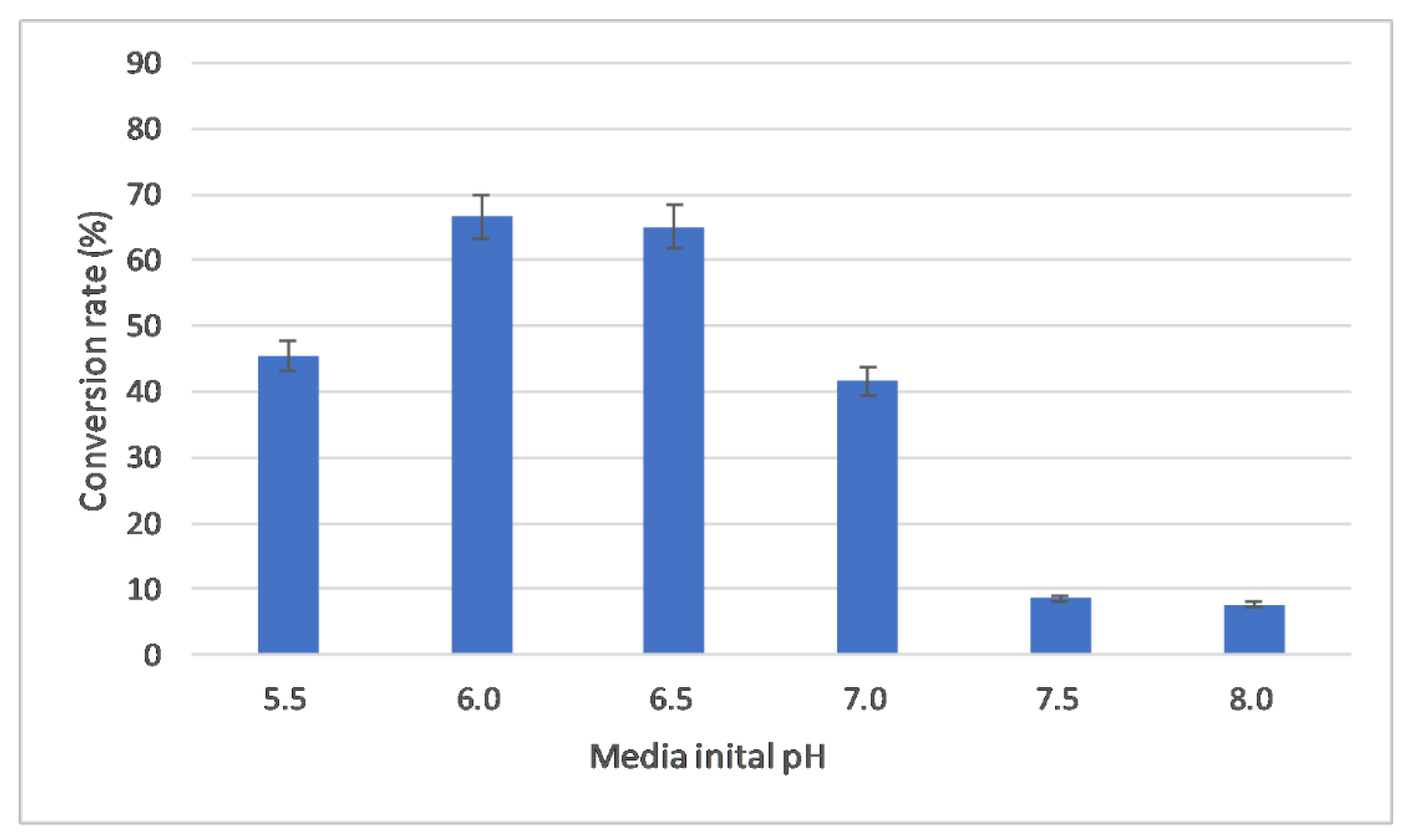
2.3.3. Evaluation of Initial pH
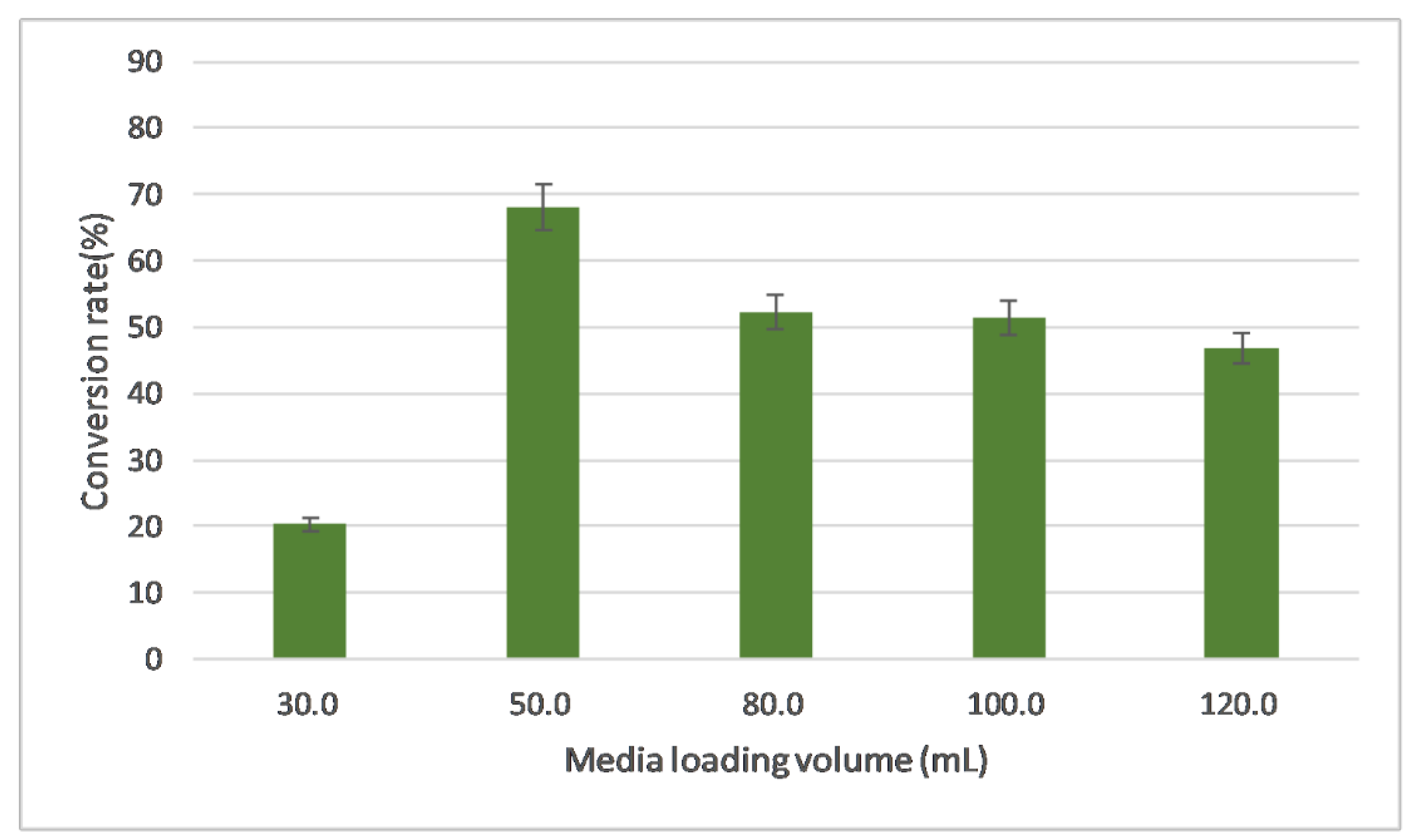
2.3.4. Evaluation of Media Volume
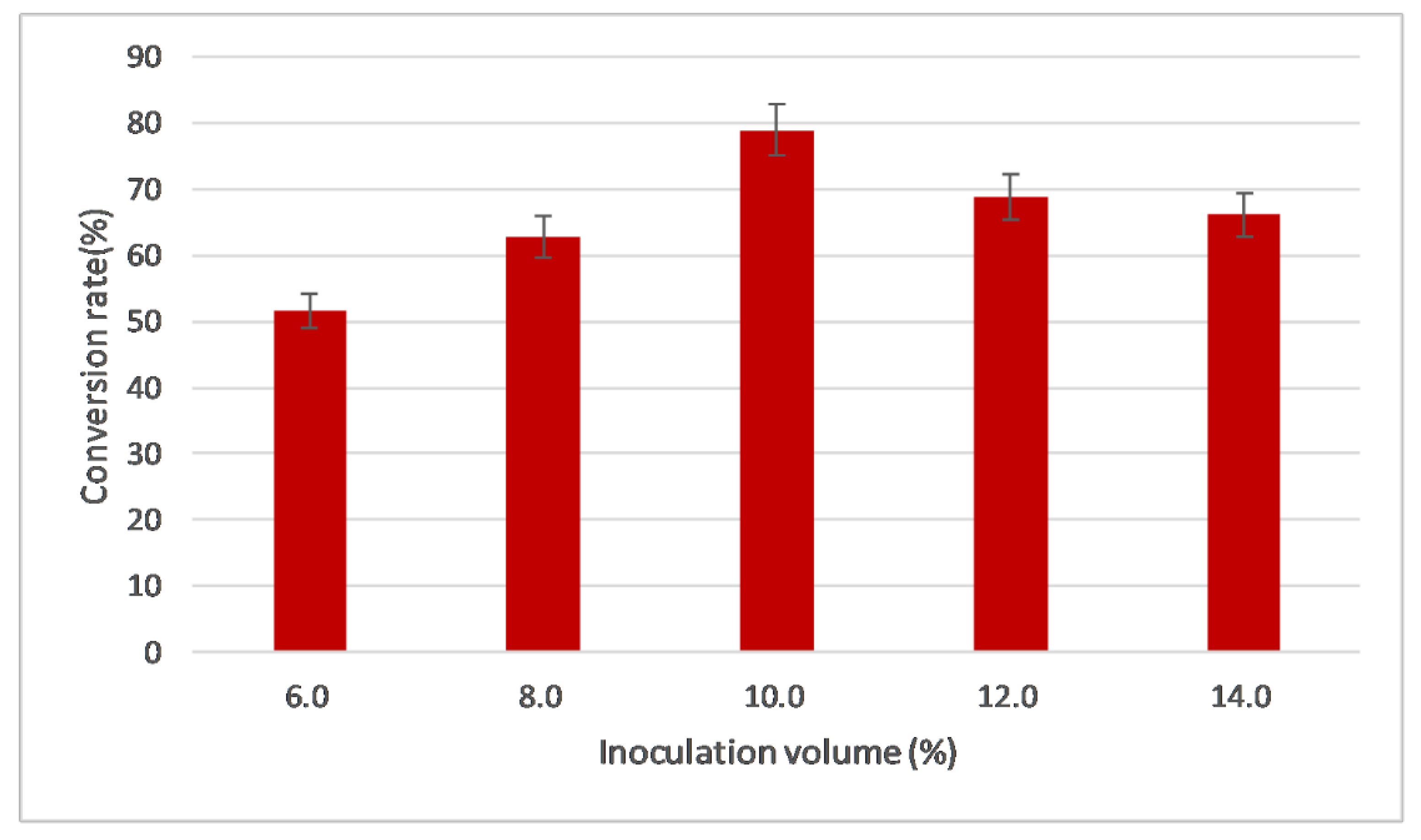
2.3.5. Evaluation of Inoculation Volume
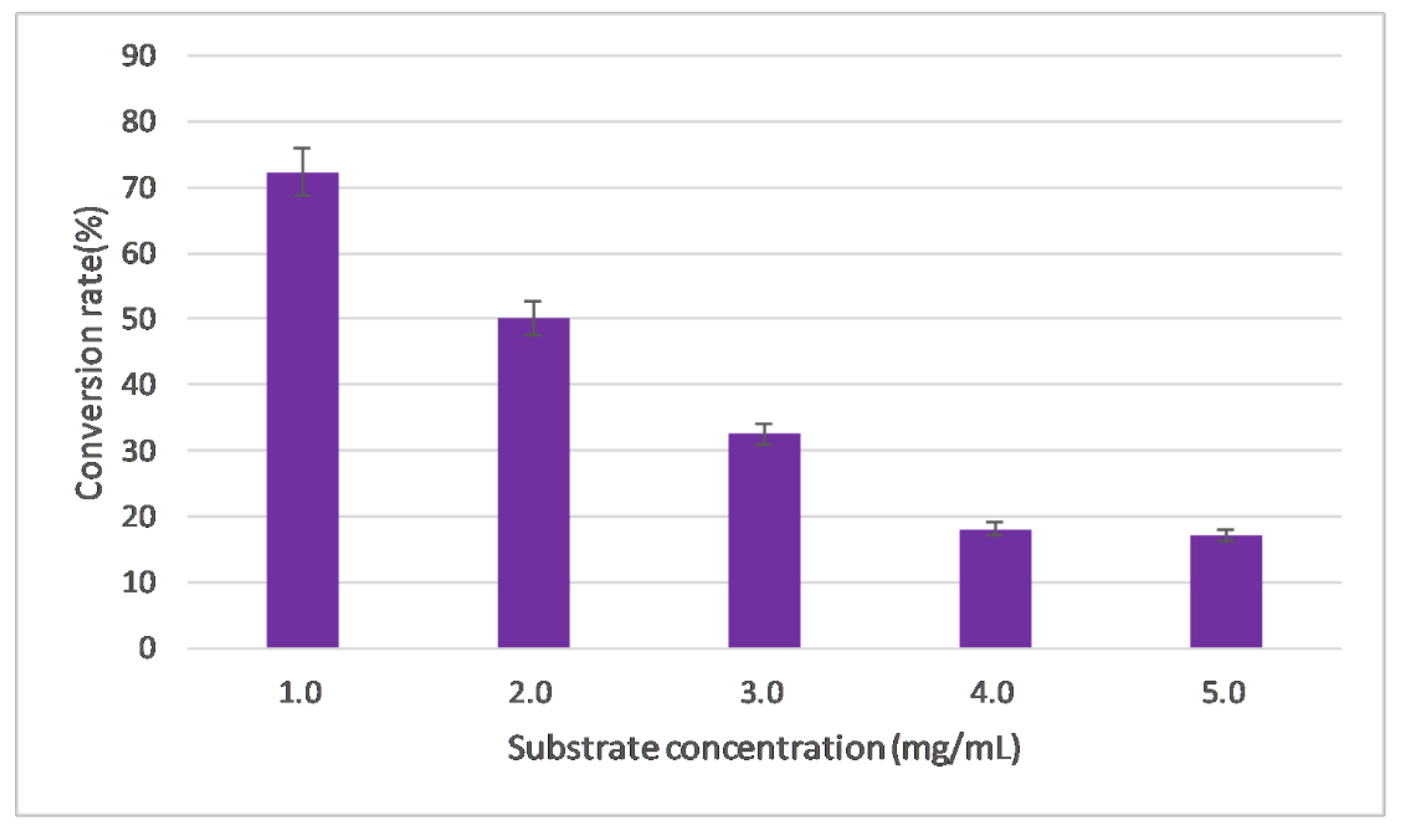
2.3.6. Evaluation of Substrate Concentration
2.4. Orthogonal Optimization
2.5. Structure Identification for the Rabeprazole Sulfide Biotransformation
3. Materials and Methods
3.1. Materials and Strains
3.2. Media and Strain Cultivations
3.3. Thin-Layer Chromatography (TLC)
3.4. Extraction of Metabolites and HPLC Detection
3.5. Isolation and Identification of Major Metabolite
3.6. Determination of the Standard Curve
4. Conclusions and Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- FDA. Approval for Aciphex (Rabeprazole Sodium) Tablets. 2002. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/nda/2002/021456_aciphex.cfm (accessed on 25 April 2003).
- Duan, Z.; Wang, Y.; Zhang, L.; Cao, X.; Fu, L.; Li, Z.; Zhang, J. An application of continuous flow microreactor in the synthesis and extraction of rabeprazole. Int. J. Chem. React. Eng. 2021, 19, 287–294. [Google Scholar] [CrossRef]
- Nishiguchi, S.; Izumi, T.; Kouno, T.; Sukegawa, J.; Ilies, L.; Nakamura, E. Synthesis of Esomeprazole and Related Proton Pump Inhibitors through Iron-Catalyzed Enantioselective Sulfoxidation. ACS Catal. 2018, 8, 9738–9743. [Google Scholar] [CrossRef]
- Lu, C.; Jia, Y.; Song, Y.; Li, X.; Sun, Y.; Zhao, J.; Wang, S.; Shi, L.; Wen, A.; Ding, L. Application of a liquid chromatographic/tandem mass spectrometric method to a urinary excretion study of rabeprazole and two of its metabolites in healthy human urine. J. Chromatogr. B 2015, 988, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Zannikos, P.N.; Doose, D.R.; Leitz, G.J.; Rusch, S.; Gonzalez, M.D.; Solanki, B.; Haddad, I.; Mulberg, A.E. Pharmacokinetics and Tolerability of Rabeprazole in Children 1 to 11 Years Old with Gastroesophageal Reflux Disease. J. Pediatr. Gastroenterol. Nutr. 2011, 52, 691–701. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.; Park, M.-J.; Kim, A.; Lee, B.-J. In vitro metabolic stability of moisture-sensitive rabeprazole in human liver microsomes and its modulation by pharmaceutical excipients. Arch. Pharmacal Res. 2008, 31, 406–413. [Google Scholar] [CrossRef]
- Miura, M.; Satoh, S.; Tada, H.; Habuchi, T.; Suzuki, T. Stereoselective metabolism of rabeprazole-thioether to rabeprazole by human liver microsomes. Eur. J. Clin. Pharmacol. 2005, 62, 113–117. [Google Scholar] [CrossRef]
- Sharara, A.I. Rabeprazole: The role of proton pump inhibitors in Helicobacter pylori eradication. Expert Rev. Anti-Infect. Ther. 2005, 3, 863–870. [Google Scholar] [CrossRef]
- Lun, J.; Ma, S.; Xue, M.; Zhao, P.; Song, Y.; Guo, X. Simultaneous enantiomeric analysis of five proton-pump inhibitors in soil and sediment using a modified QuEChERS method and chiral high performance liquid chromatography coupled with tandem mass spectrometry. Microchem. J. 2020, 160, 105625. [Google Scholar] [CrossRef]
- Marelli, S.; Pace, F. Rabeprazole for the treatment of acid-related disorders. Expert Rev. Gastroenterol. Hepatol. 2012, 6, 423–435. [Google Scholar] [CrossRef]
- Yousuf, M.; Jamil, W.; Mammadova, M.Y.A.K. Microbial Bioconversion: A Regio-specific Method for Novel Drug Design and Toxicological Study of Metabolites. Curr. Pharm. Biotechnol. 2019, 20, 1156–1162. [Google Scholar] [CrossRef]
- Asha, S.; Vidyavathi, M. Cunninghamella—A microbial model for drug metabolism studies—A review. Biotechnol. Adv. 2009, 27, 16–29. [Google Scholar] [CrossRef]
- Murphy, C.D. Drug metabolism in microorganisms. Biotechnol. Lett. 2015, 37, 19–28. [Google Scholar] [CrossRef]
- Keum, Y.-S.; Lee, Y.-H.; Kim, J.-H. Metabolism of methoxychlor by Cunninghamella elegans ATCC36112. J. Agric Food Chem. 2009, 57, 7931–7937. [Google Scholar] [CrossRef]
- Choudhary, M.I.; Khan, N.T.; Musharraf, S.G.; Anjum, S.; Rahman, A.U. Biotransformation of adrenosterone by filamentous fungus, Cunninghamella elegans. Steroids 2007, 72, 923–929. [Google Scholar] [CrossRef]
- Hanson, R.L.; Matson, J.A.; Brzozowski, D.B.; LaPorte, T.L.; Springer, D.M.; Patel, R.N. Hydroxylation of Mutilin by Streptomyces griseus and Cunninghamella echinulata. Org. Process Res. Dev. 2002, 6, 482–487. [Google Scholar] [CrossRef]
- Ibrahim, A.; Khalifa, S.I.; Khafagi, I.; Youssef, D.T.; Khan, S.; Mesbah, M.; Khan, I. Microbial Metabolism of Biologically Active Secondary Metabolites from Nerium oleander L. Chem. Pharm. Bull. 2008, 56, 1253–1258. [Google Scholar] [CrossRef] [Green Version]
- Kouzi, S.A.; Chatterjee, P.; Pezzuto, J.M.; Hamann, M.T. Microbial Transformations of the Antimelanoma Agent Betulinic Acid. J. Nat. Prod. 2000, 63, 1653–1657. [Google Scholar] [CrossRef]
- Ning, L.; Zhan, J.; Qu, G.; Zhong, L.; Guo, H.; Bi, K.; Guo, D. Biotransformation of triptolide by Cunninghamella blakesleana. Tetrahedron 2003, 59, 4209–4213. [Google Scholar] [CrossRef]
- Zhang, D.; Evans, F.E.; Freeman, J.P.; Duhart, B.; Cerniglia, C.E. Biotransformation of amitriptyline by Cunninghamella elegans. Drug Metab. Dispos. 1995, 23, 1417–1425. [Google Scholar]
- Wu, Y.; Lu, Y.; Yi, Y.; Wang, A.; Wang, W.; Yang, M.; Fan, B.; Chen, G. Biotransformation of asiatic acid by Cunninghamella echinulata and Circinella muscae to discover anti-neuroinflammatory derivatives. Nat. Prod. Res. 2022, 36, 1–6. [Google Scholar] [CrossRef]
- Khan, M.F.; Murphy, C.D. Cytochrome P450 5208A3 is a promiscuous xenobiotic biotransforming enzyme in Cunninghamella elegans. Enzym. Microb. Technol. 2022, 161, 110102. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Zhao, Y.; Gao, X.; Zhang, D.; Ma, Y.; Yang, L.; Sun, P. A Novel Antimalarial Metabolite in Erythrocyte from the Hydroxylation of Dihydroartemisinin by Cunninghamella elegans. Front. Chem. 2022, 10, 850133. [Google Scholar] [CrossRef] [PubMed]
- Sousa, I.P.; de SousaTeixeira, M.V.; Freitas, J.A.; Ferreira, A.G.; Pires, L.M.; Santos, R.A.; Heleno, V.C.G.; Furtado, N.A.J.C. Production of More Potent Anti- Candida Labdane Diterpenes by Biotransformation Using Cunninghamella elegans. Chem. Biodivers. 2022, 19, e202100757. [Google Scholar] [CrossRef] [PubMed]
- Barrero, A.F.; Oltra, J.E.; Raslan, A.D.S.; Saúde, D.A. Microbial Transformation of Sesquiterpene Lactones by the Fungi Cunninghamella echinulata and Rhizopus oryzae. J. Nat. Prod. 1999, 62, 726–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Hansen, E.B., Jr.; Deck, J.; Heinze, T.M.; Henderson, A.; Korfmacher, W.A.; Cerniglia, C.E. Fungal transformations of antihistamines: Metabolism of cyproheptadine hydrochloride by Cunninghamella elegans. Xenobiotica 1997, 27, 301–315. [Google Scholar] [CrossRef]
- Zhou, L.; Xu, W.; Chen, Y.; Zhao, J.; Yu, N.; Fu, B.; You, S. Stereoselective epoxidation of curcumol and curdione by Cunninghamella elegans AS 3.2028. Catal. Commun. 2012, 28, 191–195. [Google Scholar] [CrossRef]
- Grafinger, K.E.; Wilke, A.; König, S.; Weinmann, W. Investigating the ability of the microbial model Cunninghamella elegans for the metabolism of synthetic tryptamines. Drug Test. Anal. 2018, 11, 721–729. [Google Scholar] [CrossRef]
- Nykodemová, J.; Šuláková, A.; Palivec, P.; Češková, H.; Rimpelová, S.; Šíchová, K.; Leonhardt, T.; Jurásek, B.; Hájková, K.; Páleníček, T.; et al. 2C-B-Fly-NBOMe Metabolites in Rat Urine, Human Liver Microsomes and C. elegans: Confirmation with Synthesized Analytical Standards. Metabolites 2021, 11, 775. [Google Scholar] [CrossRef]
- Xie, Z.Y.; Huang, H.H.; Zhong, D.F. Biotransformation of pantoprazole by the fungus Cunninghamella blakesleeana. Xenobiotica 2005, 35, 467–477. [Google Scholar] [CrossRef]
- Sutherland, J.B.; Freeman, J.P.; Heinze, T.M.; Moody, J.D.; Parshikov, I.A.; Williams, A.J.; Zhang, D. Oxidation of phenothiazine and phenoxazine by Cunninghamella elegans. Xenobiotica 2001, 31, 799–809. [Google Scholar] [CrossRef]
- Zhang, D.; Freeman, J.P.; Sutherland, J.B.; Walker, A.E.; Yang, Y.; Cerniglia, C.E. Biotransformation of chlorpromazine and methdilazine by Cunninghamella elegans. Appl. Environ. Microbiol. 1996, 62, 798–803. [Google Scholar] [CrossRef]
- Bachmann, F.; zu Schwabedissen, H.E.M.; Duthaler, U.; Krähenbühl, S. Cytochrome P450 1A2 is the most important enzyme for hepatic metabolism of the metamizole metabolite 4-methylaminoantipyrine. Br. J. Clin. Pharmacol. 2021, 88, 1885–1896. [Google Scholar] [CrossRef]
- Ren, H.; Dhanaraj, P.; Enoch, I.V.; Paulraj, M.S.; Paulraj, M.S. Synthesis and Biological Evaluation of 4-Aminoantipyrine Analogues. Med. Chem. 2022, 18, 26–35. [Google Scholar] [CrossRef]
- Eliwa, D.; Albadry, M.A.; Ibrahim AR, S.; Kabbash, A.; Meepagala, K.; Khan, I.A.; El-Aasr, M.; Ross, S.A. Biotransformation of papaverine and in silico docking studies of the metabolites on human phosphodiesterase 10a. Phytochemistry 2021, 183, 112598. [Google Scholar] [CrossRef]
- Yang, W.; Jiang, T.; Acosta, D.; Davis, P.J. Microbial models of mammalian metabolism: Involvement of cytochrome P450 in the N-demethylation of N-methylcarbazole by Cunninghamella echinulata. Xenobiotica 1993, 23, 973–982. [Google Scholar] [CrossRef]
- Hansen, E.B., Jr.; Cho, B.P.; Korfmacher, W.A.; Cerniglia, C.E. Fungal transformations of antihistamines: Metabolism of brompheniramine, chlorpheniramine, and pheniramine to N-oxide and N-demethylated metabolites by the fungus Cunninghamella elegans. Xenobiotica 1995, 25, 1081–1092. [Google Scholar] [CrossRef]
- Fan, H.X.; Zhou, Z.Q.; Peng, J.; Wu, B.J.; Chen, H.R.; Bao, X.F.; Mu, Z.Q.; Jiao, W.H.; Yao, X.S.; Gao, H. A microbial model of mammalian metabolism: Biotransformation of 4,5-dimethoxyl-canthin-6-one using Cunninghamella blakesleeana CGMCC 3.970. Xenobiotica 2017, 47, 284–289. [Google Scholar] [CrossRef]
- Ibrahim, A.-R.S.; Galal, A.M.; Ahmed, M.S.; Mossa, G.S. O-Demethylation and Sulfation of 7-Methoxylated Flavanones by Cunninghamella elegans. Chem. Pharm. Bull. 2003, 51, 203–206. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Wang, Y.; Jiang, Y.; Hu, M.; Li, S.; Zhai, Q. Biocatalytic synthesis of C3 chiral building blocks by chloroperoxidase-catalyzed enantioselective halo-hydroxylation and epoxidation in the presence of ionic liquids. Biotechnol. Prog. 2015, 31, 724–729. [Google Scholar] [CrossRef]
- Morozov, A.N.; Chatfield, D.C. Chloroperoxidase-catalyzed epoxidation of cis-beta-methylstyrene: Distal pocket flexibility tunes catalytic reactivity. J. Phys. Chem. B 2012, 116, 12905–12914. [Google Scholar] [CrossRef] [Green Version]
- de Hoog, H.M.; Nallani, M.; Cornelissen, J.J.L.M.; Rowan, A.E.; Nolte, R.J.M.; Arends, I.W.C.E. Biocatalytic oxidation by chloroperoxidase from Caldariomyces fumago in polymersome nanoreactors. Org. Biomol. Chem. 2009, 7, 4604–4610. [Google Scholar] [CrossRef] [PubMed]
- Kellner, H.; Pecyna, M.J.; Buchhaupt, M.; Ullrich, R.; Hofrichter, M. Draft Genome Sequence of the Chloroperoxidase-Producing Fungus Caldariomyces fumago Woronichin DSM1256. Genome Announc. 2016, 4, e00774-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, X.; Li, H.; Jiang, Y.; Hu, M.; Li, S.; Zhai, Q. Rapid and efficient degradation of bisphenol A by chloroperoxidase from Caldariomyces fumago: Product analysis and ecotoxicity evaluation of the degraded solution. Biotechnol. Lett. 2016, 38, 1483–1491. [Google Scholar] [CrossRef] [PubMed]
- Buchhaupt, M.; Hüttmann, S.; Sachs, C.C.; Bormann, S.; Hannappel, A.; Schrader, J. Caldariomyces fumago DSM1256 Contains Two Chloroperoxidase Genes, Both Encoding Secreted and Active Enzymes. J. Mol. Microbiol. Biotechnol. 2015, 25, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Buchhaupt, M.; Hüttmann, S.; Schrader, J. White Mutants of Chloroperoxidase-Secreting Caldariomyces fumago as Superior Production Strains, Revealing an Interaction between Pigmentation and Enzyme Secretion. Appl. Environ. Microbiol. 2012, 78, 5923–5925. [Google Scholar] [CrossRef] [Green Version]
- Buchhaupt, M.; Ehrich, K.; Hüttmann, S.; Guder, J.; Schrader, J. Over-expression of chloroperoxidase in Caldariomyces fumago. Biotechnol. Lett. 2011, 33, 2225–2231. [Google Scholar] [CrossRef]
- Höfler, G.T.; But, A.; Hollmann, F. Haloperoxidases as catalysts in organic synthesis. Org. Biomol. Chem. 2019, 17, 9267–9274. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Huang, H.-H.; Liu, L.; Zhong, D.-F. Transformation of Verapamil by Cunninghamella blakesleeana. Appl. Environ. Microbiol. 2004, 70, 2722–2727. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | A/Culture Time (Days) | B/Inoculation Volume (%) | C/Media Volume (mL) | D/Media Starting pH | E/Substrate Concentration (g/L) |
|---|---|---|---|---|---|
| Level | |||||
| 1 | 5 | 8 | 30 | 5.5 | 1 |
| 2 | 6 | 10 | 50 | 6.0 | 2 |
| 3 | 7 | 12 | 80 | 6.5 | 3 |
| 4 | 8 | 14 | 100 | 7.0 | 4 |
| Factors Experiment No. | A | B | C | D | E | Conversion Rate |
|---|---|---|---|---|---|---|
| 1 | 1 | 1 | 1 | 1 | 1 | 0.352 |
| 2 | 1 | 2 | 2 | 2 | 2 | 0.315 |
| 3 | 1 | 3 | 3 | 3 | 3 | 0.348 |
| 4 | 1 | 4 | 4 | 4 | 4 | 0.201 |
| 5 | 2 | 1 | 2 | 3 | 4 | 0.213 |
| 6 | 2 | 2 | 1 | 4 | 3 | 0.124 |
| 7 | 2 | 3 | 4 | 1 | 2 | 0.336 |
| 8 | 2 | 4 | 3 | 2 | 1 | 0.566 |
| 9 | 3 | 1 | 3 | 4 | 2 | 0.347 |
| 10 | 3 | 2 | 4 | 3 | 1 | 0.171 |
| 11 | 3 | 3 | 1 | 2 | 4 | 0.135 |
| 12 | 3 | 4 | 2 | 1 | 3 | 0.343 |
| 13 | 4 | 1 | 4 | 2 | 3 | 0.724 |
| 14 | 4 | 2 | 3 | 1 | 4 | 0.053 |
| 15 | 4 | 3 | 2 | 4 | 1 | 0.094 |
| 16 | 4 | 4 | 1 | 2 | 3 | 0.033 |
| K1 | 1.216 | 1.636 | 0.644 | 1.084 | 1.184 | |
| K2 | 1.24 | 0.664 | 0.964 | 1.812 | 1.032 | |
| K3 | 0.996 | 0.912 | 1.312 | 0.764 | 1.54 | |
| K4 | 1.304 | 1.144 | 1.432 | 0.764 | 0.604 | |
| k1 | 0.304 | 0.409 | 0.161 | 0.271 | 0.296 | |
| k2 | 0.310 | 0.166 | 0.241 | 0.453 | 0.258 | |
| k3 | 0.249 | 0.228 | 0.328 | 0.191 | 0.385 | |
| k4 | 0.326 | 0.286 | 0.358 | 0.191 | 0.151 | |
| R(Range analysis) | 0.084 | 0.243 | 0.197 | 0.244 | 0.234 | |
| Best combination | A2 | B1 | C4 | D2 | E1 |
| Ingredients | No.1 (g) | No.2 (g) | No.3 (g) | No.4 (g) |
|---|---|---|---|---|
| Sucrose | 30 | 15 | ||
| Yeast extracts | 20 | 0.5 | 2 | |
| Corn steep liquor | 5 | |||
| Peptone | 0.5 | 5 | 5 | |
| Glucose | 4 | 15 | 50 | |
| K2HPO4 | 16 | 0.5 | 1 | 4 |
| KH2PO4 | 2 | 5 | ||
| MgSO4 | 0.5 | 0.5 | 0.2 | |
| FeSO4 | 0.05 | 0.01 | 0.05 | |
| KCl | 0.5 | 0.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, M.; Zhu, H.; Wang, J.; Xu, W.; Xu, W. An O-Demethylation Metabolite of Rabeprazole Sulfide by Cunninghamella blakesleeana 3.970 Biotransformation. Catalysts 2023, 13, 15. https://doi.org/10.3390/catal13010015
Song M, Zhu H, Wang J, Xu W, Xu W. An O-Demethylation Metabolite of Rabeprazole Sulfide by Cunninghamella blakesleeana 3.970 Biotransformation. Catalysts. 2023; 13(1):15. https://doi.org/10.3390/catal13010015
Chicago/Turabian StyleSong, Ming, Hongxiang Zhu, Jian Wang, Weizhuo Xu, and Wei Xu. 2023. "An O-Demethylation Metabolite of Rabeprazole Sulfide by Cunninghamella blakesleeana 3.970 Biotransformation" Catalysts 13, no. 1: 15. https://doi.org/10.3390/catal13010015