
Correlation Relationship between Phase Inversion of Pickering Emulsions and Biocatalytic Activity of Microbial Transformation of Phytosterols

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
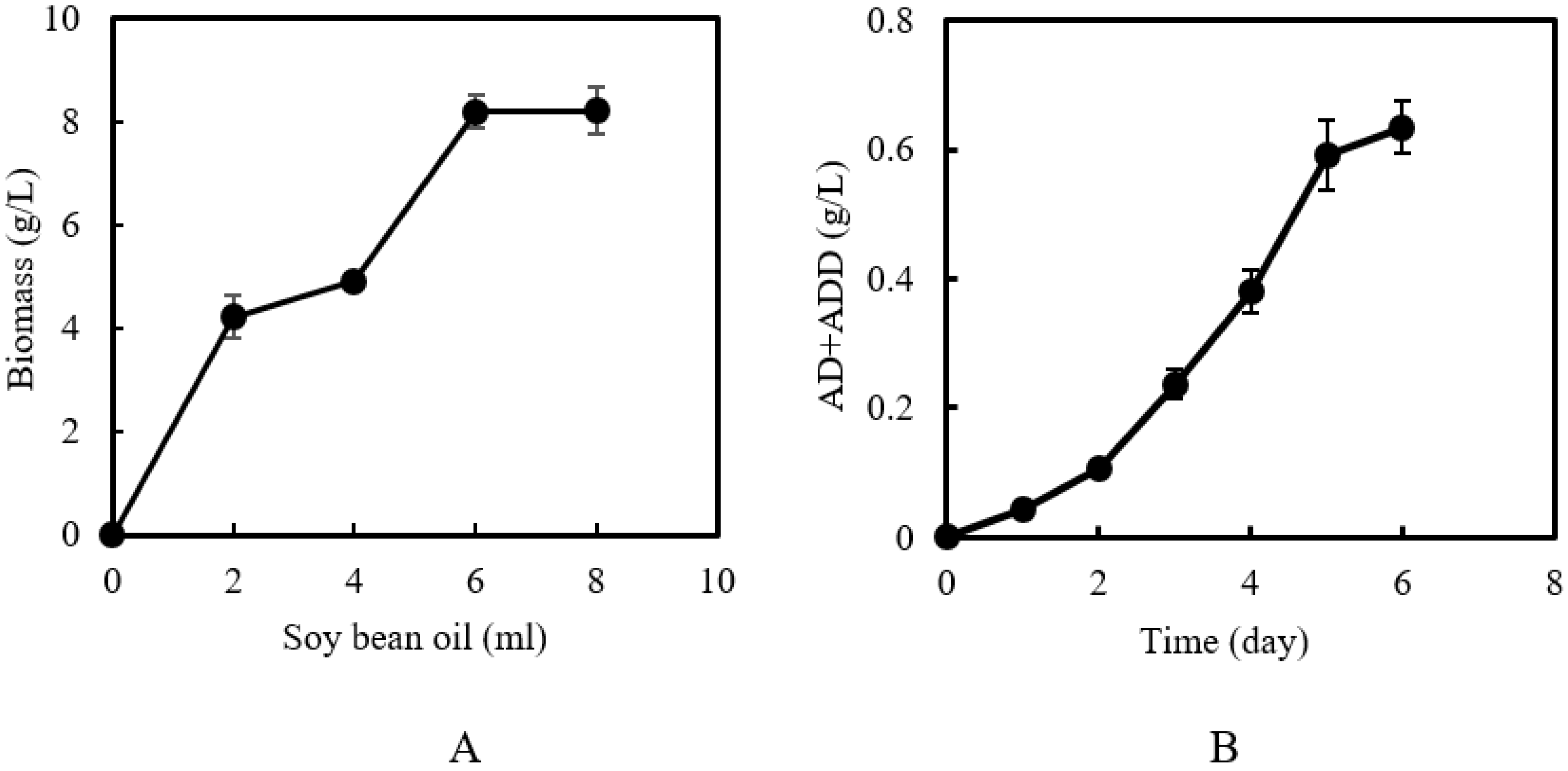
2.1. Plant Oil Acting Both as Co-Substrate and as Solvent
2.2. Effect of Volume Ratio of Oil to Water
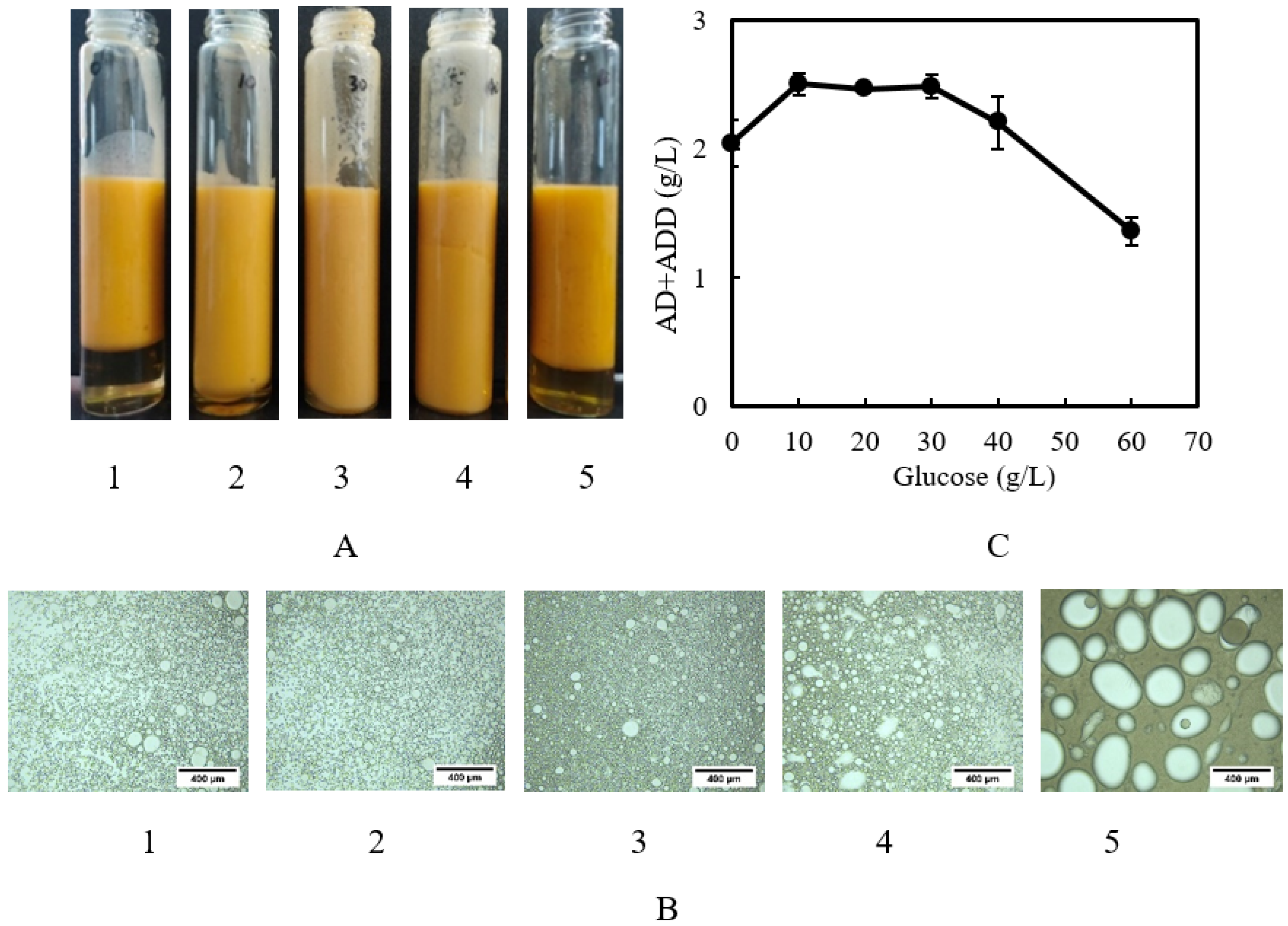
2.3. Influence of Glucose as a Co-Substrate
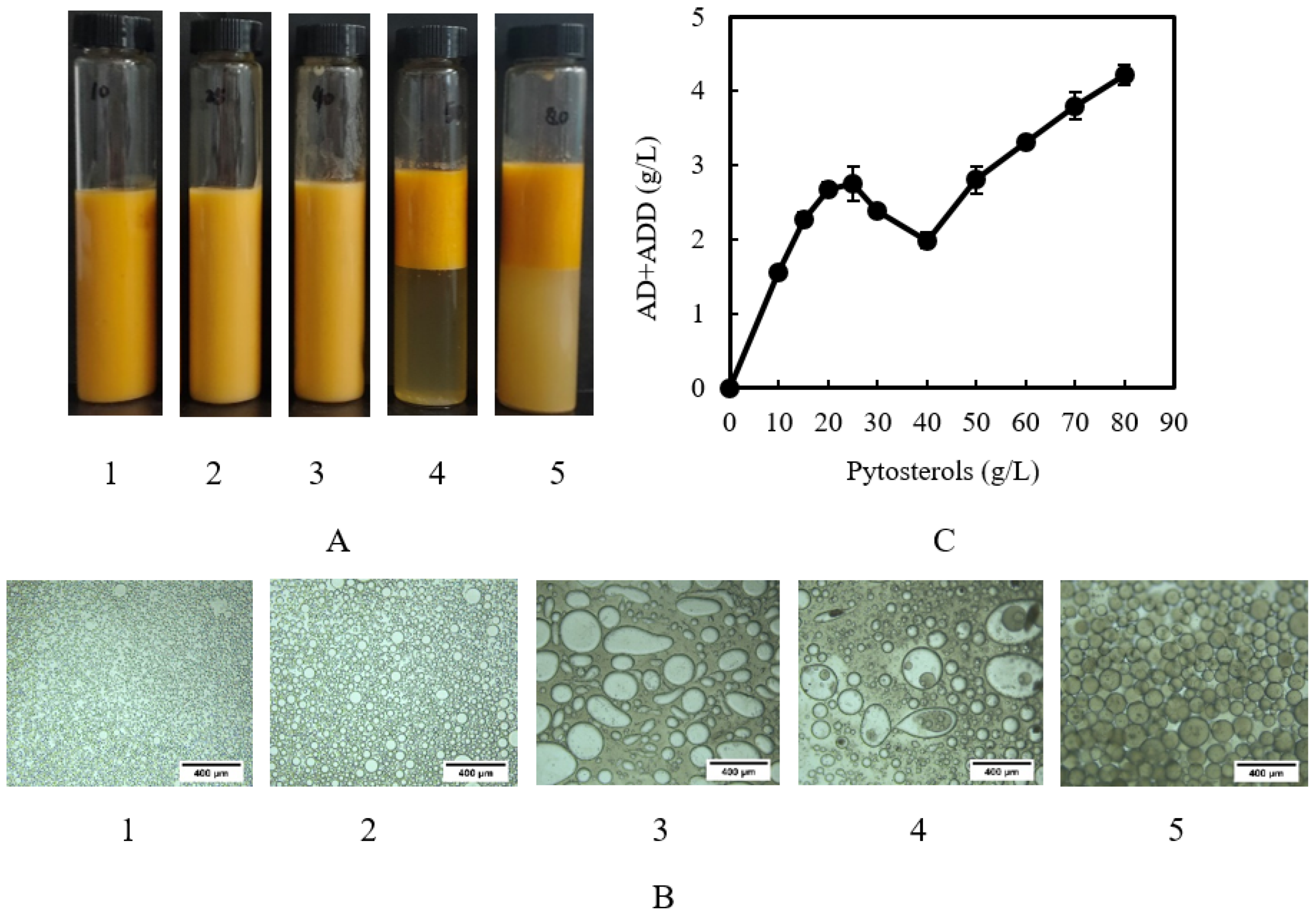
2.4. Microbial Transformation of Crystal Phytosterols
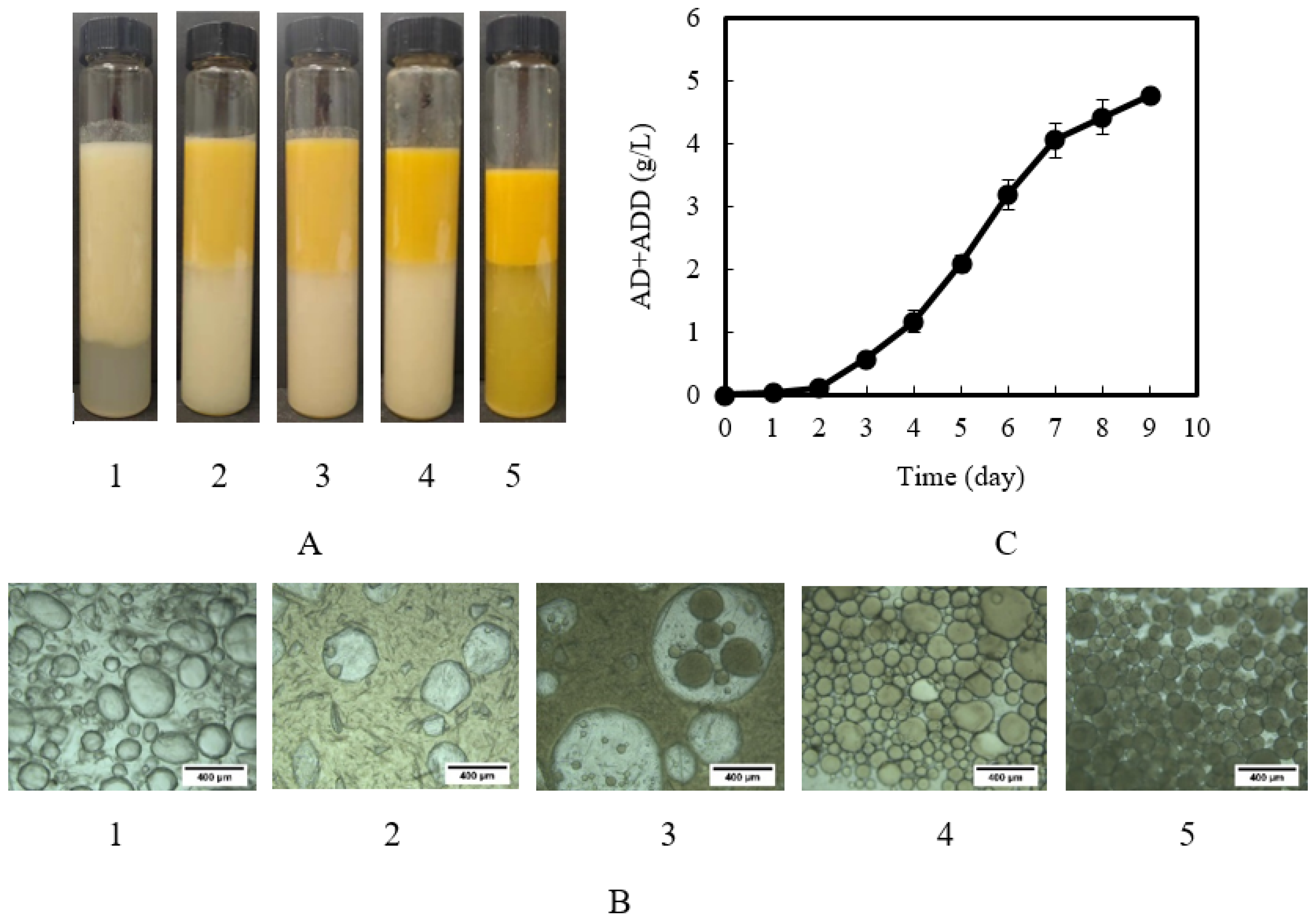
2.5. Time Course of Microbial Transformation
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Microbial Transformation
4.3. Analysis of AD and ADD Concentrations
4.4. Estimation of Microbial Growth
4.5. Investigation of Pickering Emulsions
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Feng, J.; Wu, Q.; Zhu, D.; Ma, Y. Biotransformation enables innovations toward green synthesis of steroidal pharmaceuticals. ChemSusChem 2022, 15, e202102399. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, P.; Cruz, A.; Angelova, B.; Pinheiro, H.M.; Cabral, J.M.S. Microbial conversion of steroid compounds: Recent developments. Enzyme Microb. Technol. 2003, 32, 688–705. [Google Scholar] [CrossRef]
- Liu, N.; Feng, J.; Zhang, R.; Chen, X.; Li, X.; Yao, P.; Wu, Q.; Ma, Y.; Zhu, D. Efficient microbial synthesis of key steroidal intermediates from bio-renewable phytosterols by genetically modified Mycobacterium fortuitum strains. Green Chem. 2019, 21, 4076–4083. [Google Scholar] [CrossRef]
- Lorena, F.-C.; Beatriz, G.; José, L. New insights on steroid biotechnology. Front Microbiol. 2018, 9, 958. [Google Scholar] [CrossRef]
- Phase, N.; Patil, S. Natural oils are better than organic solvents for the conversion of soybean sterols to 17-ketosteroids by Mycobacterium fortuitum. World J. Microbiol. Biotechnol. 1994, 10, 228–229. [Google Scholar] [CrossRef]
- Cruz, A.; Fernandes, P.; Cabral, J.M.S.; Pinheiro, H.M. Whole-cell bioconversion of β-sitosterol in aqueous-organic two-phase systems. J. Mol. Catal. B 2001, 11, 579–585. [Google Scholar] [CrossRef]
- Donova, M.V.; Dovbnya, D.V.; Koshcheyenko, K.A. Modified CDs-mediated enhancement of microbial sterol-side-chain degradation. In Proceedings of the Eighth International Symposium on Cyclodextrin, Budapest, Hungary, 31 March–2 April 1996; pp. 527–530. [Google Scholar]
- Su, L.; Xu, S.; Shen, Y.; Xia, M.; Ren, X.; Wang, L.; Shang, Z.; Wang, M. The sterol carrier hydroxypropyl-β-cyclodextrin enhances the metabolism of phytosterols by Mycobacterium neoaurum. Appl. Environ. Microbiol. 2020, 86, e00441-20. [Google Scholar] [CrossRef]
- Heipieper, H.J.; Neumann, G.; Cornelissen, S.; Meinhardt, F. Solvent-tolerant bacteria for biotransformation in two-phase fermentation systems. Appl. Microbiol. Biotechnol. 2007, 74, 961–973. [Google Scholar] [CrossRef]
- Xiao, X.; He, J.-K.; Guan, Y.-X.; Yao, S.-J. Effect of cholinium amino acids ionic liquids as cosolvents on the bioconversion of phytosterols by Mycobacterium sp. resting cells. ACS Sustain. Chem. Eng. 2020, 8, 17124–17132. [Google Scholar] [CrossRef]
- Yuan, J.-J.; Guan, Y.-X.; Wang, Y.-T.; Wang, H.-Q.; Yao, S.-J. Side-chain cleavage of phytosterols by Mycobacterium sp. MB 3683 in a biphasic ionic liquid/aqueous system. J. Chem. Technol. Biotechnol. 2016, 91, 2631–2637. [Google Scholar] [CrossRef]
- Carvalho, F.; Marques, M.P.C.; de Carvalho, C.C.C.R.; Cabral, J.M.S.; Fernandes, P. Sitosterol bioconversion with resting cells in liquid polymer based systems. Biores. Technol. 2009, 100, 4050–4053. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhao, F.; Chen, D.; Li, D. Biotransformation of phytosterol to adustra-diene-dione by resting cells of Mycobacterium in cloud point system. Process Biochem. 2006, 41, 557–561. [Google Scholar] [CrossRef]
- Wang, Z.; Zhao, F.; Hao, X.; Chen, D.; Li, D. Microbial transformation of hydrophobic compound in cloud point system. J. Mol. Catal. B 2004, 27, 147–153. [Google Scholar] [CrossRef]
- Xie, H.; Zhao, W.; Ali, D.C.; Zhang, X.; Wang, Z. Interfacial biocatalysis in bacteria-stabilized Pickering emulsions for microbial transformation of hydrophobic chemicals. Catal. Sci. Technol. 2021, 11, 2816–2826. [Google Scholar] [CrossRef]
- Zhao, W.; Xie, H.; Zhang, X.; Wang, Z. Crystal substrate inhibition during microbial transformation of phytosterols in Pickering emulsions. Appl. Microbiol. Biotechnol. 2022, 106, 2403–2414. [Google Scholar] [CrossRef]
- Lemos, D.A.; Sonego, J.L.S.; Boschiero, M.V.; Araujo, E.C.C.; Cruz, A.J.G.; Badino, A.C. Selection and application of nontoxic solvents in extractive ethanol fermentation. Biochem. Eng. J. 2017, 127, 128–135. [Google Scholar] [CrossRef]
- Li, N.; Han, Z.; O’Donnell, T.J.; Kurasaki, R.; Kajihara, L.; Williams, P.G.; Tang, Y.; Su, W.W. Production and excretion of astaxanthin by engineered Yarrowia lipolytica using plant oil as both the carbon source and the biocompatible extractant. Appl. Microbiol. Biotechnol. 2020, 104, 6977–6989. [Google Scholar] [CrossRef]
- Li, H.; Fu, Z.; Li, H.; Zhang, X.; Shi, J.; Xu, Z. Enhanced biotransformation of dehydroepiandrosterone to 3β,7α,15α-trihydroxy-5-androsten-17-one with Gibberella intermedia CA3-1 by natural oils addition. J. Ind. Microbiol. Biotechnol. 2014, 41, 1497–1504. [Google Scholar] [CrossRef]
- Stefanov, S.; Yankov, D.; Beschkov, V. Biotransformation of phytosterols to androstenedione in two phase water-oil systems. Chem. Biochem. Eng. Q 2006, 20, 421–427. [Google Scholar]
- Xu, Y.-G.; Guan, Y.-X.; Wang, H.-Q.; Yao, S.-J. Microbial side-chain cleavage of phytosterols by Mycobacteria in vegetable oil/aqueous two-phase system. Appl. Biochem. Biotechnol. 2014, 174, 522–533. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, Y.; Shen, Y.; Zhang, X.; Xu, S.; Shang, Z.; Xia, M.; Wang, M. Efficient production of androstenedione by repeated batch fermentation in waste cooking oil media through regulating NAD+/NADH ratio and strengthening cell vitality of Mycobacterium neoaurum. Bioresour. Technol. 2019, 279, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Binks, B.P.; Fletcher, P.; Holt, B.; Beaussoubre, P.; Wong, K. Phase inversion of particle-stabilised perfume oil-water emulsions: Experiment and theory. Phys. Chem. Chem. Phys. 2010, 12, 11954–11966. [Google Scholar] [CrossRef] [PubMed]
- Lan, M.; Zheng, J.; Huang, C.; Wang, Y.; Hu, W.; Lu, S.; Liu, F.; Ou, S. Water-in-oil Pickering emulsions stabilized by microcrystalline phytosterols in oil: Fabrication mechanism and application as a salt release system. J. Agric. Food Chem. 2022, 70, 5408–5416. [Google Scholar] [CrossRef]
- Binks, B.P.; Lumsdon, S.O. Transitional phase inversion of solid-stabilized emulsions using particle mixtures. Langmuir 2000, 16, 3748–3756. [Google Scholar] [CrossRef]
- Glonke, S.; Sadowski, G.; Brandenbusch, C. Applied catastrophic phase inversion: A continuous non-centrifugal phase separation step in biphasic whole-cell biocatalysis. J. Ind. Microbiol. Biotechnol. 2016, 43, 1527–1535. [Google Scholar] [CrossRef]
- Xie, H.; Zhao, W.; Zhang, X.; Wang, Z. Demulsification of bacteria-stabilized Pickering emulsion using modified silica nanoparticle. ACS Appl. Mater. Interfaces 2022, 14, 24102–24112. [Google Scholar] [CrossRef]
- Jiang, H.; Liu, L.; Li, Y.; Yin, S.; Ngai, T. Inverse Pickering emulsion stabilized by binary particles with contrasting characteristics and functionality for interfacial biocatalysis. ACS Appl. Mater. Interfaces 2020, 12, 4989–4997. [Google Scholar] [CrossRef]
- Nallamilli, T.; Binks, B.P.; Mani, E.; Basavaraj, M.G. Stabilization of Pickering emulsions with oppositely charged latex particles: Influence of various parameters and particle arrangement around droplets. Langmuir 2015, 31, 11200–11208. [Google Scholar] [CrossRef]
- Chen, Z.; Zhao, C.; Ju, E.; Ji, H.; Ren, J.; Binks, B.P.; Qu, X. Design of surface-active artificial enzyme particles to stabilize Pickering emulsions for high-performance biphasic biocatalysis. Adv. Mater. 2016, 28, 1682–1688. [Google Scholar] [CrossRef]
- Meng, T.; Bai, R.; Wang, W.; Yang, X.; Guo, T.; Wang, Y. Enzyme-loaded mesoporous silica particles with tuning wettability as a Pickering catalyst for enhancing biocatalysis. Catalysts 2019, 9, 78. [Google Scholar] [CrossRef] [Green Version]
- Jia, X.; Zhang, S.; Li, J.; Xia, J.; Yao, R.; Zhao, X.; Wu, B.; Bai, F.; Xiao, Y. Engineered bacterial biofloc formation enhancing phenol removal and cell tolerance. Appl. Microbiol. Biotechnol. 2020, 104, 1187–1199. [Google Scholar] [CrossRef] [PubMed]
- Binks, B.P.; Philip, J.; Rodrigues, J.A. Inversion of silica-stabilized emulsions induced by particle concentration. Langmuir 2005, 21, 3296–3302. [Google Scholar] [CrossRef] [PubMed]
- Whitby, C.P.; Parthipan, R. Influence of particle concentration on multiple droplet formation in Pickering emulsions. J. Colloid Interf. Sci. 2019, 554, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Lei, C.; Xie, Y.; Wu, Y.; Li, Y.; Li, B.; Pei, Y.; Liu, S. Properties and stability of water-in-water emulsions stabilized by microfibrillated bacterial cellulose. Food Hydrocoll. 2022, 130, 107698. [Google Scholar] [CrossRef]
- Pera-Titus, M.; Leclercq, L.; Clacens, J.M.; de Campo, F.; Nardello-Rataj, V. Pickering interfacial catalysis for biphasic systems: From emulsion design to green reactions. Angew. Chem. Int. Ed. 2015, 54, 2006–2021. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; van Oers, M.C.M.; Rutjes, F.P.J.T.; van Hest, J.C.M. Polymersome colloidosomes for enzyme catalysis in a biphasic system. Angew. Chem. Int. Ed. 2012, 51, 10746–10750. [Google Scholar] [CrossRef]
- Rodriguez, A.M.B.; Binks, B.P. Catalysis in Pickering emulsions. Soft Matter 2020, 16, 10221–10243. [Google Scholar] [CrossRef]
- Achberger, A.M.; Doyle, S.M.; Mills, M.I.; Holmes, C.P.; Quigg, A.; Sylvan, J.B. Bacteria-oil microaggregates are an important mechanism for hydrocarbon degradation in the marine water column. mSystems 2021, 6, e01105-21. [Google Scholar] [CrossRef]
- Shao, M.; Zhang, X.; Rao, Z.; Xu, M.; Yang, T.; Li, H.; Xu, Z.; Yang, S. A mutant form of 3-ketosteroid-Δ1 -dehydrogenase gives altered androst-1, 4-diene-3, 17-dione/androst-4-ene-3, 17-dione molar ratios in steroid biotransformations by Mycobacterium neoaurum ST-095. J. Ind. Microbiol. Biotechnol. 2016, 43, 691–701. [Google Scholar] [CrossRef]
- Angelova, B.; Fernandes, P.; Cruz, A.; Pinheiro, H.M.; Mutafov, S.; Cabral, J.M.S. Hydroxylation of androstenedione by resting Rhodococcus sp cells in organic media. Enzyme Microb. Technol. 2005, 37, 718–722. [Google Scholar] [CrossRef]
- He, K.; Sun, H.; Song, H. Engineering phytosterol transport system in Mycobacterium sp. strain MS136 enhances production of 9a-hydroxy-4-androstene-3, 17-dione. Biotechnol. Lett. 2018, 40, 673–678. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Yang, J.; He, K.; Wang, Y.-P.; Song, H. Enhancing production of 9a-hydroxy-androst-4-ene-3, 17-dione (9-OH-AD) from phytosterols by metabolic pathway engineering of Mycobacteria. Chem. Eng. Sci. 2021, 230, 116195. [Google Scholar] [CrossRef]
- Song, N.; Wang, A.; Li, J.; Zhu, Z.; Shi, H.; Ma, X.; Sun, D. Study on influencing factors of Pickering emulsions stabilized by hydroxyapatite nanoparticles with nonionic surfactants. Soft Matter 2018, 14, 3889–3901. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, W.; Xie, H.; Zhang, X.; Wang, Z. Correlation Relationship between Phase Inversion of Pickering Emulsions and Biocatalytic Activity of Microbial Transformation of Phytosterols. Catalysts 2023, 13, 72. https://doi.org/10.3390/catal13010072
Zhao W, Xie H, Zhang X, Wang Z. Correlation Relationship between Phase Inversion of Pickering Emulsions and Biocatalytic Activity of Microbial Transformation of Phytosterols. Catalysts. 2023; 13(1):72. https://doi.org/10.3390/catal13010072
Chicago/Turabian StyleZhao, Wenyu, Haisheng Xie, Xuehong Zhang, and Zhilong Wang. 2023. "Correlation Relationship between Phase Inversion of Pickering Emulsions and Biocatalytic Activity of Microbial Transformation of Phytosterols" Catalysts 13, no. 1: 72. https://doi.org/10.3390/catal13010072