1. Introduction
α-Ketoglutaric acid (KGA) is a valuable compound with many demonstrated applications in the cosmetic, pharmaceutical and food industries or as an intermediate in chemical syntheses [
1]. The chemical industry uses KGA as a substrate for synthesis of biopolymers [
2] and heterocyclic compounds [
3]. In food production it is applied as a functional additive in beverages and a nutraceutical in functional food [
4,
5,
6]. In medicine it has been used in the treatment of various diseases and for the synthesis of pharmaceuticals [
7,
8,
9,
10,
11]. The health benefits of KGA consumption have been demonstrated not only in humans but also in animals; therefore, its preparations are of interest to feed producers [
12,
13,
14,
15].
Although KGA is present in the central metabolism of every living cell, it is an intermediate molecule that is synthesized only in an amount that meets the needs of the cells. Therefore, KGA cannot be obtained from easily accessible sources such as food [
16]. Industrial production of KGA is achieved mainly by a multi-step chemical method that uses diethyl oxalate and diethyl succinate as the substrates [
3,
17]. The process yields 75% efficiency, but it causes environmental hazards and has low economic attractiveness of production because of application of raw materials derived from depleting petrochemical resources, use of harmful reagents and generation of toxic wastes and high by-production of impurities. Moreover, the product obtained by this method may be excluded from use in certain applications (e.g., food production) [
1,
18,
19].
The possibility of converting various carbon sources to KGA using microbial fermentation has attracted scientists for several decades [
1,
4,
19]. A wide variety of microorganisms have been found to be capable of synthesizing KGA. Nevertheless, the limiting factors for the use of most of them on a commercial scale are low titer, low yield, formation of by-products, intolerance to low pH and inhibitor presence and high nutritional requirements. Microorganisms that appear to be particularly effective in the biosynthesis of KGA are yeast, among which
Yarrowia lipolytica has received special attention. In comparison to other yeast species,
Y. lipolytica enables more efficient production of KGA from a very wide range of substrates and the use of simple culture media that meet low nutritional and vitamin supplementation requirements [
1,
19,
20].
Previously we presented the genetically engineered strain of
Y. lipolytica as a prospective producer of KGA from mixed renewable carbon sources (glycerol + rapeseed oil), along with simple technology of product recovery from post-culture broth [
21]. Moreover, the biomass of this strain was proved to be of nutritional and health-beneficial characteristics especially desired in food and feed applications. The aim of the present work was to increase the efficiency of KGA production from mixed glycerol/rapeseed-oil-based media by the transformant strain
Y. lipolytica CBS146773 by optimizing the conditions of biosynthesis.
2. Results and Discussion
In our earlier investigation, the transformant strain
Yarrowia lipolytica CBS146773, formerly named 1.31.GUT1/6.CIT1/3.E34672 [
21], was obtained by genetic engineering. This strain was characterized by overexpression of genes encoding glycerol kinase (
GUT1), citrate synthase (
CIT1) and mitochondrial acid transporter (
YALI0E34672g) and was found to be a good producer of KGA in the process with synergistic co-feeding of glycerol and rapeseed oil [
21]. The use of glycerol and rapeseed oil as substrates for
Y. lipolytica was dictated by ecological and economic reasons. Both substrates are renewable carbon sources suitable for KGA biosynthesis; however, their applications differ in the efficiency of KGA production. The yield of KGA biosynthesis obtained using rapeseed oil might exceed 100%, whereas glycerol application results in by-formation of pyruvic acid (PA), which is linked to the decreased production of KGA and negatively affects the selectivity of the process. However, in practice, the possibility of even partial replacement of the water-insoluble substrate with hydrophilic glycerol can greatly facilitate unit operations during industrial-scale processes [
21,
22]. Since the use of a mixed-substrates feeding strategy has already been demonstrated to be suitable for the process with
Y. lipolytica CBS146773, in this work we focused on the selection of the conditions of the KGA biosynthesis that are aimed at fully exploiting the genetic potential of the selected strain.
2.1. Maintenance of pH
The effect of pH-control strategy on the growth and biosynthesis of KGA by the yeast strain
Y. lipolytica CBS146773 was studied in two aspects: examination of pH value and comparison of different neutralizing agents used for maintaining pH at the appropriate level. The literature includes information about the positive effect of acidic pH on KGA biosynthesis by
Y. lipolytica yeast [
23,
24,
25,
26,
27]. In the reports, the pH level optimal for KGA production varied from 2.79 to 4.5 depending on the strain and substrate used in a specific process. In this study the pH impact was tested in the range of 3.0–4.5, using KOH as neutralizing agent. As presented in
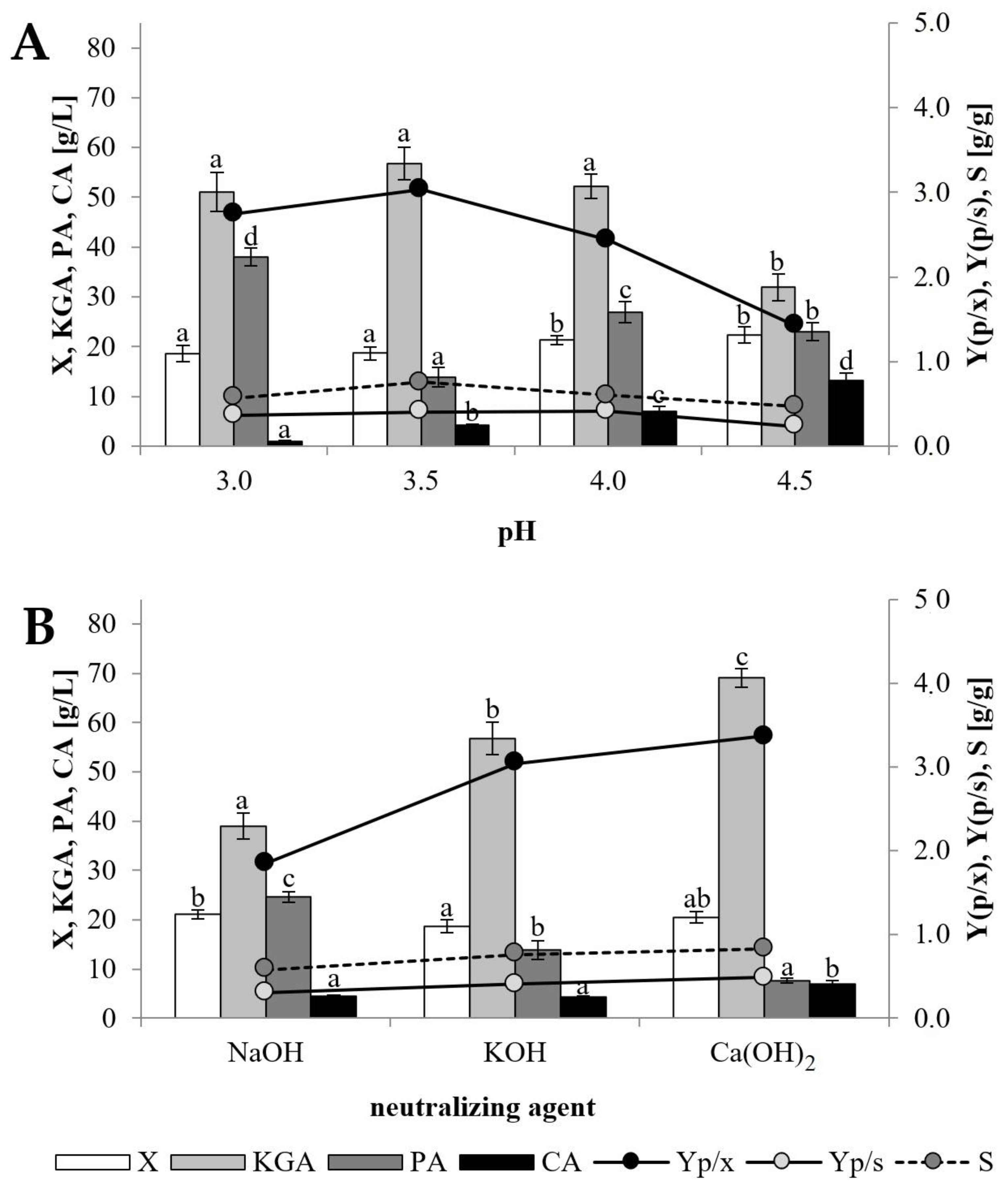
Figure 1A, the yeast strain presented good growth in the examined pH range. However, the increase in pH value resulted in an increased level of biomass from 18.6 g/L at pH 3.0 to 22.4 g/L when pH 4.5 was applied. It was found that the formation of KGA was dependent on pH. The highest KGA production of 56.8 g/L corresponding to the yield (Yp/s) of 0.41 g/g was obtained in the culture conducted at pH 3.5. Simultaneously, in the same conditions, PA content in the post-culture broth was the lowest and reached 13.9 g/L, whereas in other cultures the concentration of the acid was significantly higher (23.0–38.0 g/L). It was also observed that increasing the pH level from 3.0 to 4.5 resulted in increased citric acid (CA) by-product formation from 1.0 to 13.2 g/L. Taking into account overall production of the acid pool, the highest selectivity of the KGA biosynthesis process (0.76 = 76%) was noted when the pH was maintained at 3.5. Some authors reported that not only the pH value applied but also its control strategy, in which the pH level set at the growth phase was lowered and was controlled at the low level to the end of the cultivation process, enhancing KGA biosynthesis [
24,
27]. In the process conducted on glycerol with such a two-step pH control strategy,
Y. lipolytica WSH-Z06 produced 32.5% more KGA (53.4 g/L) than in the process in which a one-step strategy was applied [
27].
In this study the second approach to the pH control strategy was to verify whether a different neutralizing agent may affect the process of KGA formation by the examined transformant yeast strain. For this purpose, biosynthesis conducted at pH 3.5 was maintained by automatic addition of a 20% solution of NaOH, KOH or Ca(OH)
2 (
Figure 1B). In comparison to KOH, application of the two other neutralizing factors resulted in slightly higher biomass growth. The results of this experiment revealed that the type of factor used for neutralization has a significant influence on the efficiency of KGA biosynthesis. When NaOH was used, the yeast produced moderate concentrations of both KGA (39.0 g/L) and PA (24.6 g/L), which, despite the relatively low amount of CA, led to low selectivity of the process (57%). A significant increase in KGA biosynthesis was noted in the culture in which Ca(OH)
2 was applied, as yeast produced 69.1 g/L of KGA with the yield (Yp/s) of 0.49 g/g and selectivity of 83%. These results were in agreement with an earlier investigation that revealed a positive impact of Ca
2+ ions on pyruvate carboxylase [
28]. In the cultures of
Torulopsis glabrata CCTCC M202019 growing on glucose, a lower concentration of KGA (1.3 g/L) was obtained when NaOH was used as a neutralizing agent, whereas application of CaCO
3 increased KGA synthesis (11.5–12.0 g/L) [
28,
29]. This relationship was also confirmed in the culture of
Y. lipolytica WSK-Z06 performed in glycerol media, where the change of the neutralizing agent from NaOH to CaCO
3 resulted in an increase in the KGA:PA ratio from 22.0:36.9 g/L to 40.3:31.8 g/L [
27]. As a consequence of the above described results, all subsequent experiments were carried out at pH 3.5 maintained by addition of Ca(OH)
2.
2.2. Manipulation of Aeration Level
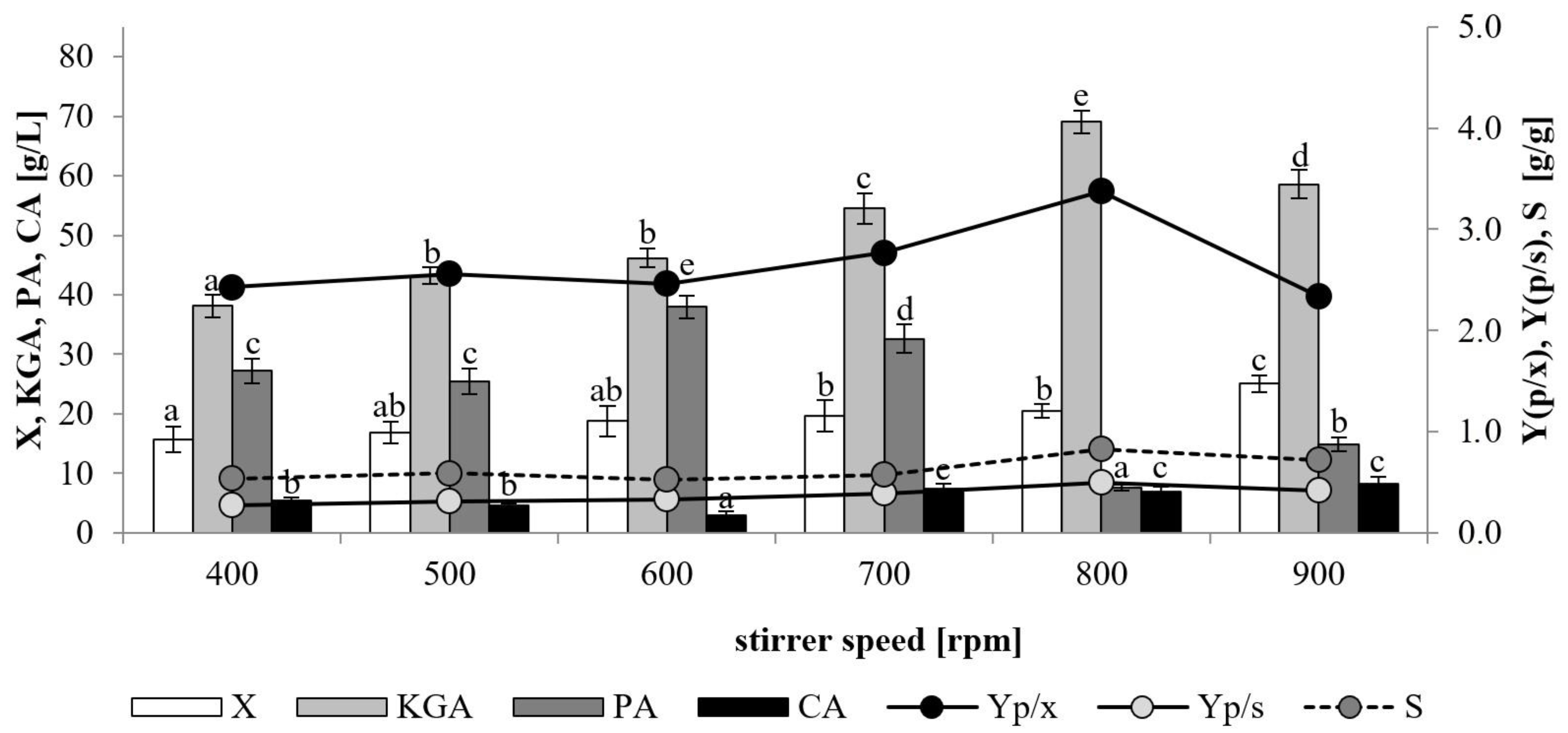
In order to evaluate the impact of dissolved oxygen level cultures were performed at various agitation rates (400–900 rpm) while maintaining a constant air flow of 0.8 vvm. A significant effect of agitation rate on growth of the yeast was observed (
Figure 2). The biomass level reached a maximum of 25.1 g/L at 900 rpm and decreased depending on the decrease in the agitation rate to 15.7 g/L at 400 rpm. In the range of 400–700 rpm biosynthesis of KGA (38.1–54.5 g/L) was accompanied by comparatively high production of PA (25.5–38 g/L). As a result, KGA production yield (Yp/s) and selectivity were at the level of 0.27–0.39 g/g and 53–59%, respectively. The highest concentration of KGA and the best parameters of its biosynthesis were obtained when the agitation rate reached 800 rpm, and a further increase to 900 rpm resulted in a decrease in the KGA biosynthesis efficiency. In these cultures, the selectivity of the process was found to be significantly higher (72–83%) than in the process that was conducted at a lower agitation speed, i.e., 400–700 rpm. The agitation rate is a parameter affecting the amount of oxygen dissolved in the culture broth. In this study, the application of an agitation rate in the range of 400–900 rpm corresponded to 20–60% pO
2, measured in the KGA production phase of the performed cultures. The aeration level has been identified as an important factor influencing KGA biosynthesis by
Y. lipolytica growing on ethanol, rapeseed oil and biodiesel waste [
23,
24,
25]. Similar to the results obtained in our study with the use of glycerol/oil media, the process of KGA production by
Y. lipolytica VKM Y-2412 conducted on biodiesel waste (a substrate containing 70.8% glycerol and 23.9% fatty acids) was also promoted by high aeration [
24]. The increase in aeration from 5% pO
2 to 50% pO
2 enabled an increase in the production of KGA from 56.8 to 80.4 g/L. In contrast, high aeration was not necessary for KGA biosynthesis from ethanol. In comparison to the culture with high aeration (50% pO
2), 1.3-times higher KGA formation (49.0 g/L) was observed when low aeration was applied (5% pO
2) [
25]. As high aeration was found to stimulate KGA biosynthesis by the examined yeast strain, an agitation rate of 800 rpm was applied in all further experiments.
2.3. Availability of Exogenous Vitamins
Yeast reported as producers of KGA have been characterized as auxotrophic for one, two or several vitamins. These vitamins are co-factors of enzymes in the Krebs cycle, and their exogenous level is one of the crucial factors affecting accumulation of KGA in auxotrophic cells [
19].
Y. lipolytica is auxotrophic only to thiamine, limitation of which is known to reduce the activity of α-ketoglutarate dehydrogenase and therefore determines KGA oversynthesis [
24]. It should be noted that auxotrophy only for one vitamin gives an advantage to the process performed by
Y. lipolytica because it requires very precise control of only one vitamin. In order to obtain thiamine limitation in the cultures with
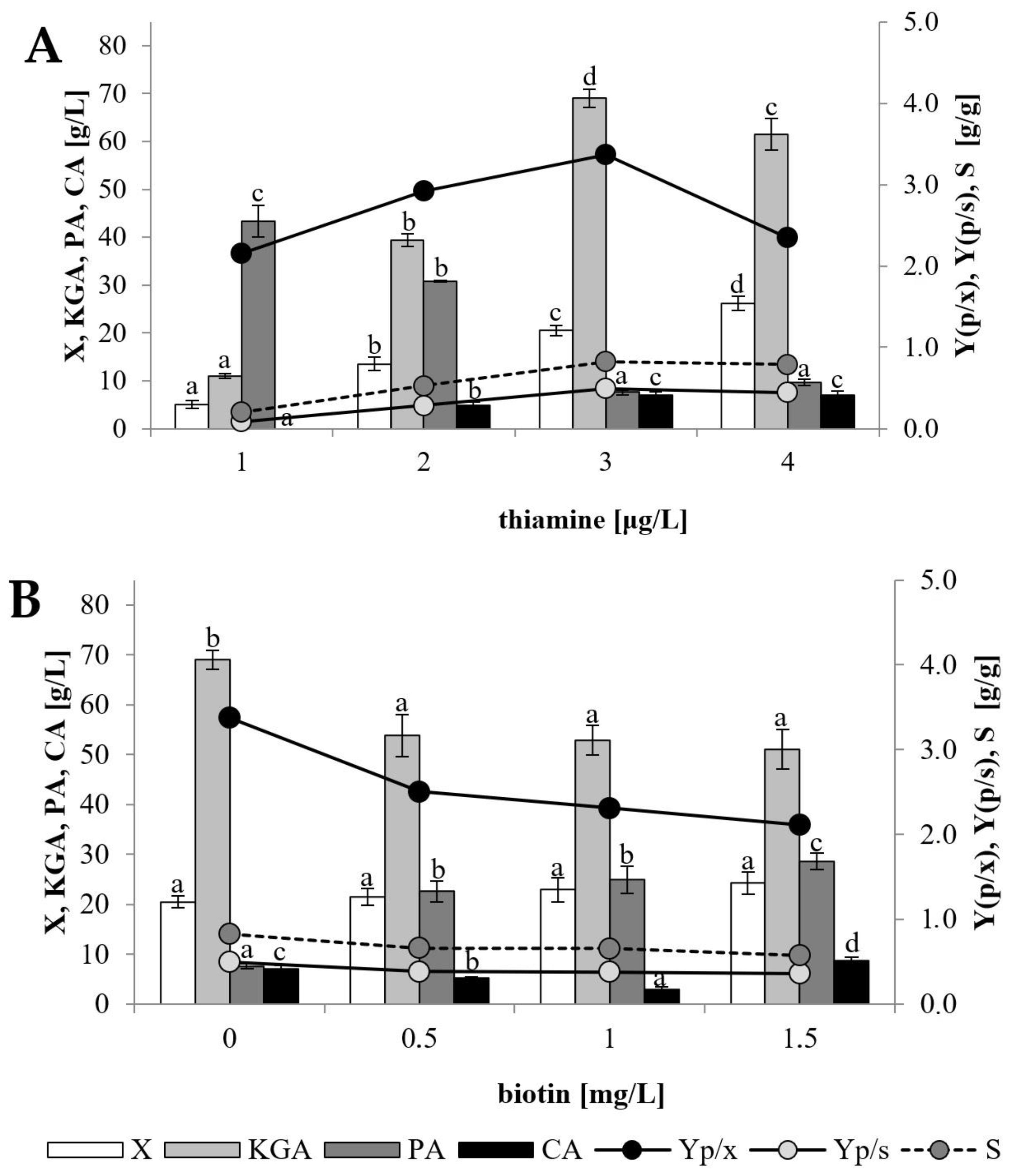
Y. lipolytica CBS146773, the vitamin concentration was applied in the very low range of 1–4 μg/L (
Figure 3A). It was clearly apparent, that thiamine had a significant impact on yeast growth, as the biomass level increased from 5.1 to 26.2 g/L with increased thiamine concentration. Increasing the vitamin addition from 1 to 3 μg/L resulted in rapid changes in both KGA and PA concentrations, but the opposite trend was observed for these acids—an increase from 11.0 to 69.1 g/L in the case of KGA and a decrease from 43.3 to 7.6 g/L in the case of PA. Thus, the selectivity of the process increased from 20% to 83% with the change of thiamine availability from 1 to 3 μg/L. No further improvement in KGA concentration or parameters of its biosynthesis was observed after the addition of 4 μg/L of thiamine. Because of the big differences in yeast growth observed between all the cultures it is worth paying attention to the parameter of yield of KGA calculated with respect to biomass formed (Yp/x). This parameter was the highest (3.37 g/g) when application of 3 μg/L of thiamine resulted in the highest amount of KGA produced. However, its value was very similar in the cultures with thiamine supplementation of 1 and 4 μg/L, where it reached 2.16 and 2.35 g/g, respectively, despite the amount of produced KGA (11.0—61.5 g/L, respectively) differing significantly between these processes. As mentioned above, the appropriate thiamine concentration is a crucial factor for effective KGA biosynthesis by the yeast belonging to the species
Y. lipolytica. The optimal concentration for KGA biosynthesis requires a balance between the amount necessary for growth and the amount determining the decreased activity of α-ketoglutarate dehydrogenase and is a strain-dependent feature. It is reported in the literature that increasing availability of thiamine (up to 200 μg/L) stimulates the growth of the yeast, whereas for KGA synthesis a “peak” is observed at a certain low vitamin concentration (0.15–4 μg/L) specific to the kind of substrate, substrate feeding method and yeast strain applied for the process [
22,
23,
24,
25,
26]. Moreover, it should be noted that by-product formation of PA also might be affected by thiamine concentration when yeast is grown on glycolytic carbon sources (glucose, fructose, glycerol, etc.), which are utilized via pyruvate because of modulation of thiamine-dependent pyruvate dehydrogenase activity [
30].
The impact of exogenous biotin addition (0–1.5 mg/L) and all subsequent experiments were performed in media supplemented with 3 μg/L of thiamine. Biotin is another vitamin which may induce accumulation of KGA by affecting the activity of pyruvate carboxylase [
30,
31,
32]. In this study, biotin addition to the culture stimulated yeast growth—in the processes supplemented with the vitamin, biomass was at the level of 21.5–24.2 g/L, whereas in the control culture its concentration reached 20.5 g/L (
Figure 3B). However, no positive effect of biotin addition on KGA production was observed. The amount of KGA in the post-culture broth decreased from 69.1 to 51.1 g/L after culture supplementation with 1.5 mg/L of biotin. Simultaneously, PA concentration increased from 7.6 g/L in the culture not supplemented with biotin to 28.6 g/L in the process where 1.5 mg/L of the vitamin was used. Theoretically, biotin presence increases the activity of pyruvate carboxylase, which catalyzes the conversion of pyruvate to oxaloacetate. Therefore, biosynthesis of KGA from substrates metabolized by the glycolysis pathway (e.g., glycerol) should be enhanced by biotin supplementation. In the shake-flask culture of
Y. lipolytica WSH-Z06, addition of 0.8 mg/L of biotin had only small positive effect on KGA biosynthesis from glycerol whereas PA production was unaffected [
26]. Interesting observations were reported by Otto et al. [
30], who studied the changes in the by-product spectrum during KGA biosynthesis from glycerol by
Y. lipolytica H355A(PYC1) T3—a strain that overexpressed pyruvate carboxylase. In comparison to the mother strain H355, higher activity of the enzyme in the transformant strain resulted in a higher biomass level, a decrease in KGA production from 133.0 to 126.9 g/L, and a simultaneous slight increase in the formation of PA and other by-products. The positive effect on yeast growth was explained by accumulation of precursor molecules (oxaloacetic, malic, succinic and fumaric acids) caused by an imbalance between enhanced activity of pyruvate carboxylase and inhibited activity of pyruvate dehydrogenase (due to thiamine limitation). Assuming that, in the present study, the addition of biotin increased the activity of pyruvate carboxylase, the same tendency was noticeable—stimulation of yeast growth and PA production with a decrease in KGA formation. Similarly, the growth of
Y. lipolytica VKM Y-2412 was slightly increased whereas KGA production was decreased and no effect on PA formation was noted when the process was conducted on biodiesel waste media upon supplementation with biotin (10–40 μg/L) [
24].
2.4. The Effect of Iron
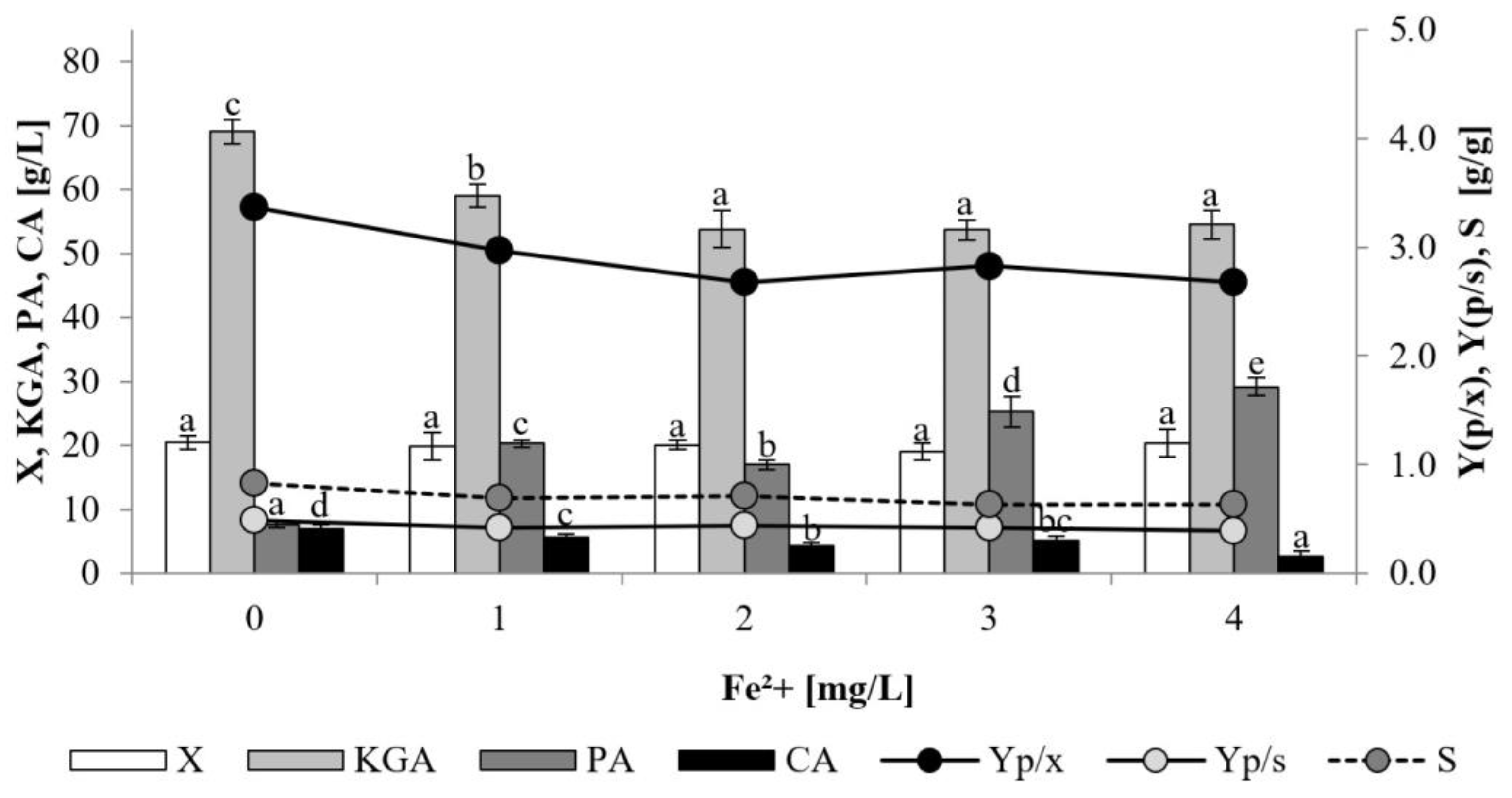
The presence of iron ions is another factor that may possibly affect KGA biosynthesis by modulating the activity of iron-dependent enzymes in the Krebs cycle, i.e., aconitase and succinate dehydrogenase [
33,
34]. In the present study the impact of iron was evaluated in the media supplemented with ammonium iron sulfate hexahydrate in an amount corresponding to 1–4 mg/L of iron ions. The presence of iron had no effect on growth of the examined yeast strain (
Figure 4). The biomass level in the cultures supplemented with iron ions was in the range of 19.0–20.4 g/L, which was comparable to the level obtained for the control culture (20.5 g/L). Addition of iron ions resulted in lower production of KGA and enhanced accumulation of PA. In the supplemented cultures yeast produced 53.7–59.0 g/L and 17.0–29.2 g/L of these acids, respectively. Compared to the non-iron-supplemented process, formation of CA was found to be slightly decreased upon the addition of iron ions (2.7–5.7 g/L). The effect of iron on KGA biosynthesis by
Y. lipolytica was investigated during cultivation on ethanol [
35,
36]. It was found that iron concentration of 0.5–2.0 mg/L stimulated KGA synthesis, whereas a further increase in iron concentration from 2–3 to 10.0 mg/L caused a gradual decrease in KGA accumulation. The positive impact of iron presence was also reported for KGA production on biodiesel waste by
Y. lipolytica [
24]. Effective biosynthesis of isocitric acid by
Y. lipolytica from rapeseed oil was noted also upon iron supplementation [
37]. In turn, the ability of
Y. lipolytica to produce erythritol was unaffected by iron ions when yeast was cultivated in bioreactor cultures on glycerol media [
38].
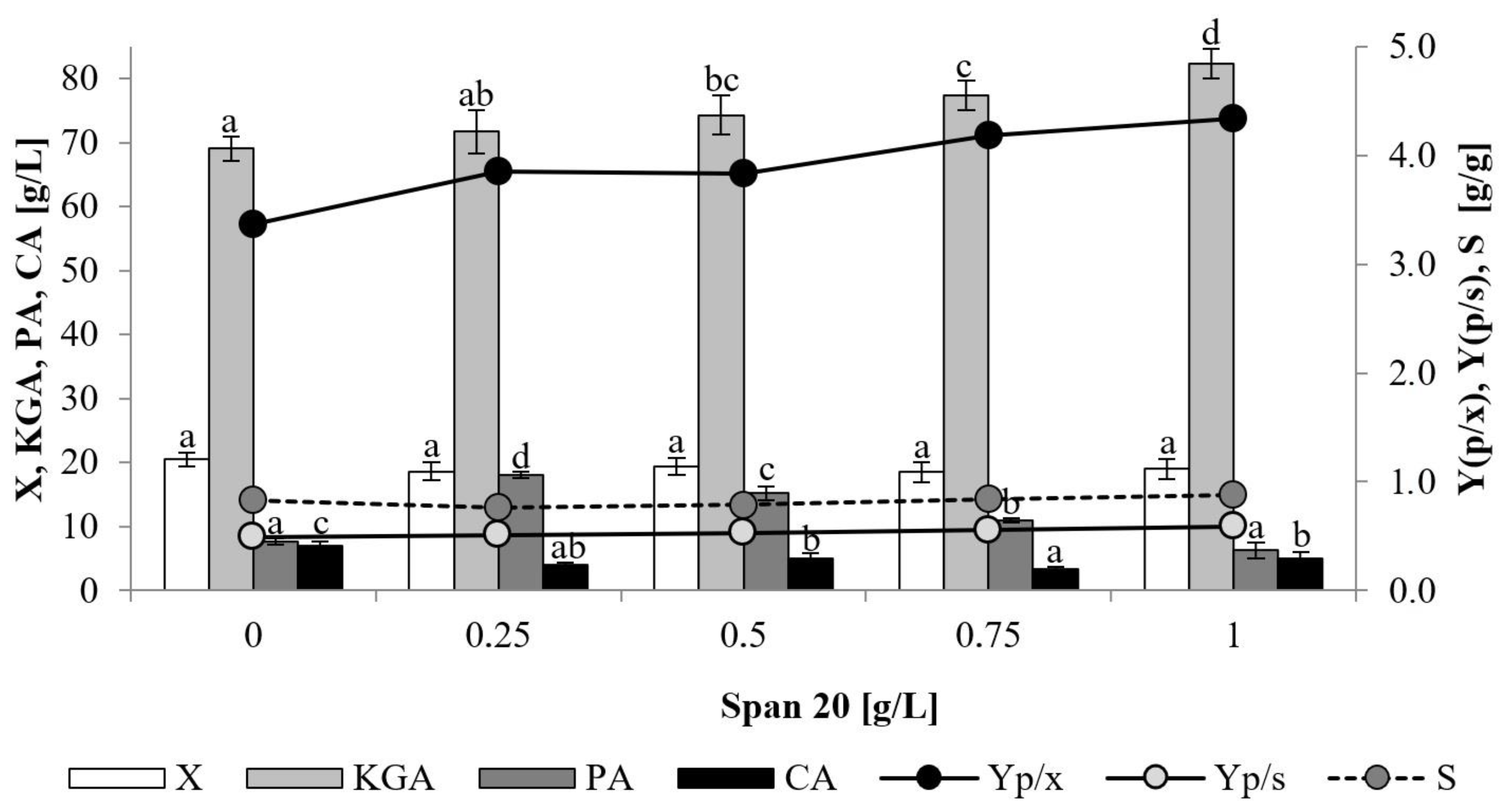
2.5. The Impact of Sorbitan Monolaurate
Sorbitan monolaurate, known under the trade name Span 20, is a non-ionic, water-insoluble, lipophilic emulsifier [
39]. This surfactant is approved for use as a food additive (E493) and the acceptable daily intake is 10 mg/kg [
40]. Literature reports mention a positive effect of Span 20 on the growth and excretion of some metabolites by microorganisms [
41,
42,
43,
44]. This surfactant, in addition to increasing the dispersion of oil substrates in the culture media, may also increase the permeability of microorganisms’ cell membranes. Thus, it facilitates the excretion of metabolites and extracellular proteins from the cell. In this study, Span 20 was applied to the culture media in the concentration ranging from 0.25 to 1 g/L (
Figure 5). In comparison to the control, the growth of the yeast was slightly lower in the presence of Span 20 and ranged 18.5–19.4 g/L. Nevertheless, addition of Span 20 had a significant positive impact on KGA biosynthesis. The increase in its addition up to 1 g/L resulted in a gradual increase in KGA production up to 82.4 g/L, corresponding to the yield (Yp/s) of 0.88 g/g and selectivity of 88% (
Figure 5 and
Figure 6). Comparatively high PA amounts (11.0–18.1 g/L) were obtained in the cultures with Span 20 addition of 0.25–0.75 g/L. In turn, production of CA was slightly decreased by the presence of Span 20 and was in the range 3.5–5.0 g/L. The positive effect of Span 20 addition was reported previously for production of oxalic acid from fatty acid waste by
Aspergillus niger [
41]. In the process with the addition of 0.75 g/L of the surfactant, the production of oxalic acid increased from 34.7 to 48.4 g/L, compared to the control culture. The addition of 0.25 g/L of Span 20 to the culture of
Y. lipolytica Wratislavia K1 cultivated on raw glycerol enhanced erythritol production from 149.6 g/L to 165.7 g/L [
42]. Moreover, although very low by-product formation of CA and KGA was noted in this process, synthesis of both acids was enhanced by the presence of the surfactant. Application of Span 20 was also proved to have a positive effect on biomass formation and β-carotene production by
Blakeslea trispora [
43,
44].
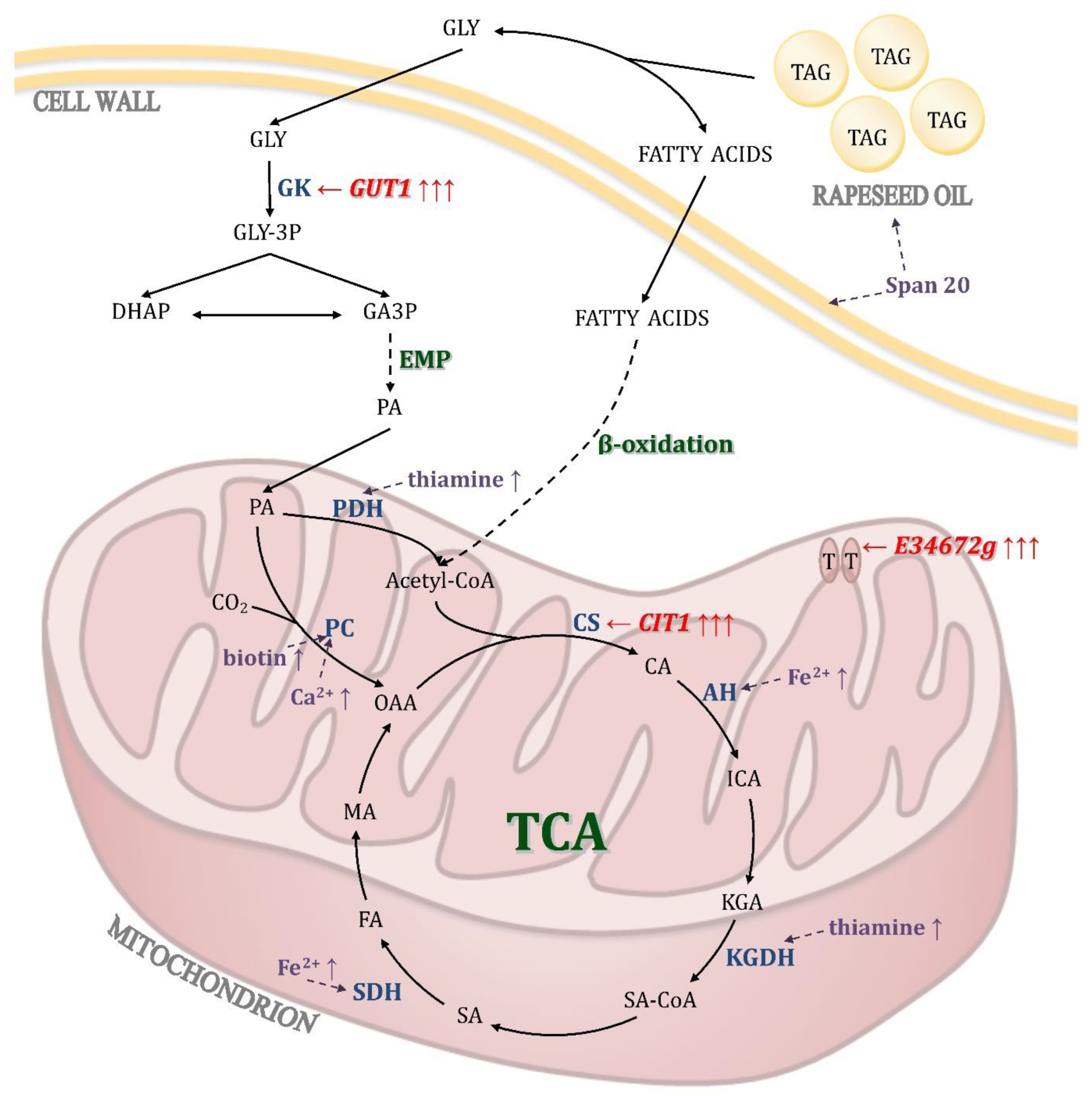
4. Conclusions
In this study the effects of selected media components and culture conditions were evaluated in order to enhance the biosynthesis of KGA by
Y. lipolytica CBS146773. The possible impact of evaluated factors is presented in
Figure 7. In the research, the source of carbon and energy was a mixture of glycerol and rapeseed oil. The addition of Span 20 was used to increase the dispersion of oil droplets and increase the permeability of cell membranes, which may facilitate the secretion of the produced metabolites [
41,
42]. Notably, substrates applied in this investigation are utilized by the yeast cell in different metabolic pathways. Glycerol is first transformed by the action of glycerol kinase to glycerol-3-phosphate, which, after being converted to glyceraldehyde, undergoes further transformations in the glycolytic pathway, resulting in the formation of pyruvate. In the mitochondrion, pyruvate dehydrogenase catalyzes the conversion of pyruvate into acetyl-CoA, which is incorporated into the tricarboxylic acid cycle with the action of citrate synthase. The transformant strain used in this study was characterized by overexpression of genes encoding glycerol kinase and citrate synthase, which was aimed at increasing the efficiency of the above-described metabolic pathway [
21]. Moreover, a gene encoding previously uncharacterized mitochondrial organic acid transporter was overexpressed in the yeast strain to investigate whether it might facilitate secretion of organic acids from mitochondrion and increase the extracellular concentration of KGA as a final product.
The second substrate—rapeseed oil—is hydrolyzed by extracellular lipases to glycerol and fatty acids. In the cell the latter undergo β-oxidation, resulting in the formation of acetyl-CoA—a compound that connects the pathways of glycerol and oil utilization. Formed from acetyl-CoA and oxaloacetate, citrate is further converted to isocitric acid by aconitase, and the subsequent transformation leads to the formation of KGA. Theoretically, at this stage, metabolism can be stimulated by the presence of iron ions, since aconitase is an iron-dependent enzyme [
33,
34]. In the presented study, however, we did not observe a positive effect of iron supplementation on KGA biosynthesis. Overproduction of KGA requires inhibition of thiamine-dependent ketoglutarate dehydrogenase, catalyzing KGA conversion to succinyl-CoA. In
Y. lipolytica, this is possible by limiting exogenous thiamine, as this yeast is unable to synthesize the pyrimidine structure of this vitamin [
24]. In accordance with literature reports, in the presented study, the concentration of thiamine was noted as the key factor determining the effective production of KGA by
Y. lipolytica [
22,
23,
24,
25,
26]. However, it should be noted that the differences in the amount of thiamine reported as necessary for KGA production might be dependent not only on the strain but also on the kind of substrate applied in the process. In comparison to glycolytic substrates, the use of fatty substrates omits the reaction catalyzed by the second thiamine-dependent enzyme—pyruvate dehydrogenase; hence, the cell’s need for this vitamin may be lower in such a process. A side effect of using thiamine limitation in cultures conducted on glycerol media is the inhibition of pyruvate dehydrogenase resulting in accumulation of pyruvate. This effect can be counteracted by increasing the activity of pyruvate carboxylase, which converts pyruvate to oxaloacetate [
31]. In this study, two factors that might stimulate the activity of this enzyme were examined: biotin and calcium ions. The presence of calcium ions was found to significantly enhance the biosynthesis of KGA, whereas a positive effect of biotin supplementation was not observed.
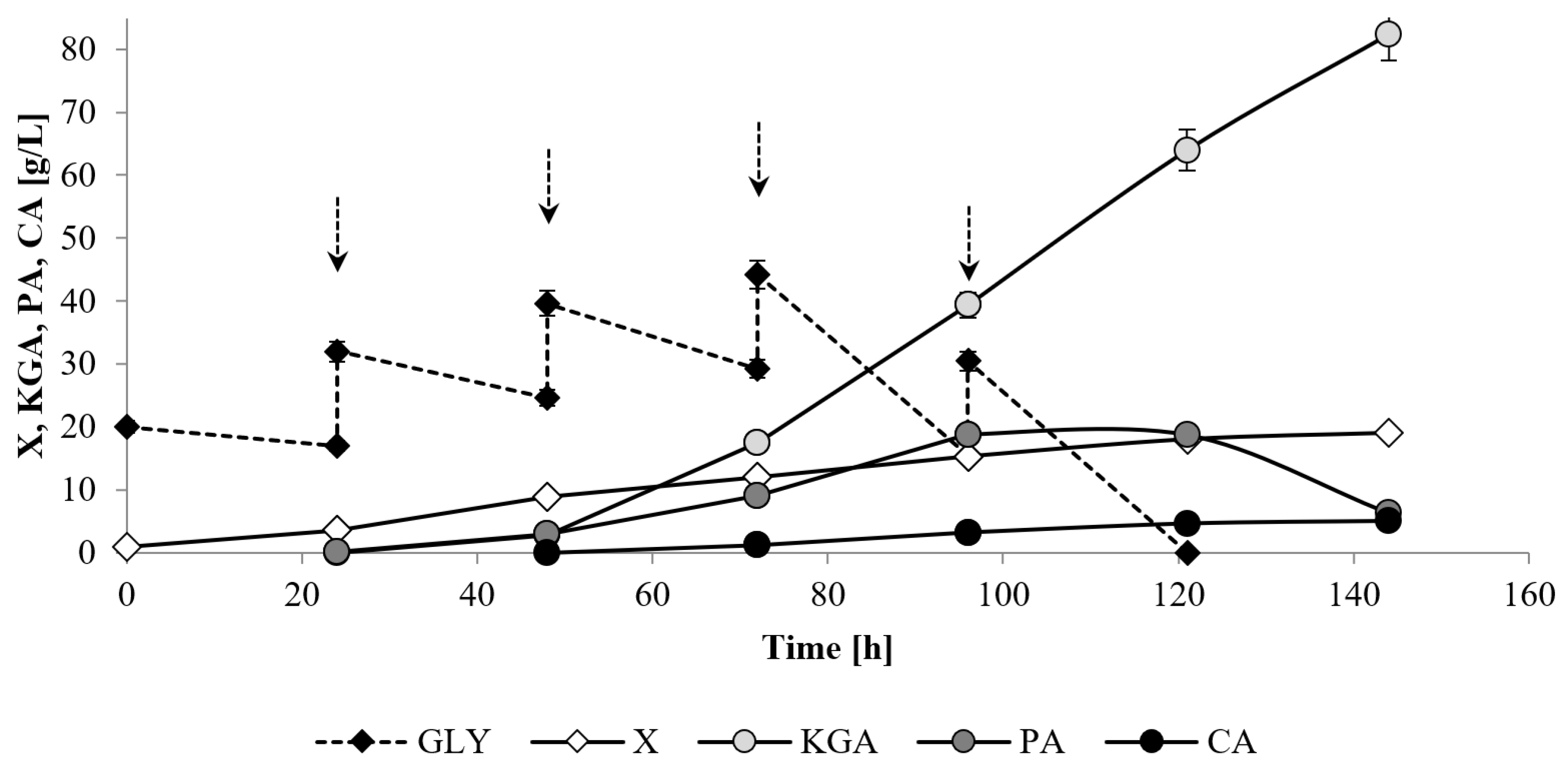
The results of the experiments performed in this study to identify the best conditions for effective KGA biosynthesis indicated the following: maintenance of pH at 3.5 by neutralization with the use of Ca(OH)
2, an agitation rate of 800 rpm and the addition of 3 μg/L of thiamine and 1 g/L of Span 20. In our earlier investigation, this transformant strain was identified as a good producer of KGA from mixed media which, after preliminary optimization of the process conditions (C:N:P ratio), was able to biosynthesize 53.1 g/L of KGA with productivity of 0.35 g/L h and yield (Yp/s) of 0.53 g/g [
21]. In the present work selection of culture conditions enabled the increase in KGA biosynthesis to 82.4 g/L. Moreover, the parameters of KGA biosynthesis were significantly improved—the productivity increased to 0.57 g/L h and the yield (Yp/s) reached 0.59 g/g. A similar amount of the acid was obtained after optimization of the KGA production process performed by
Y. lipolytica VKM Y-2412 on biodiesel waste containing glycerol and fatty acids [
24]. In the optimal conditions, yeast produced 80.4 g/L of KGA with the selectivity of 96.7%. However, the cultivation process was significantly longer (192 h) than in the present research (144 h). In the literature data the highest KGA production was reported for
Y. lipolytica H355A(PYC1-IDP1) T5 engineered for overexpression of isocitrate dehydrogenase and pyruvate carboxylase, which was able to synthesize 186.0 g/L of KGA from raw glycerol with productivity of 1.75 g/L h [
45]. However, the yield of KGA production, which is one of the factors determining the process attractiveness in terms of industrial production, obtained in the process with strain H355A(PYC1-IDP1) T5 reached only 0.36 g/g and was significantly lower than the yield obtained in this research with strain CBS146773.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






