
Biodegradation of Crystalline and Nonaqueous Phase Liquid-Dissolved ATRAZINE by Arthrobacter sp. ST11 with Cd2+ Resistance
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
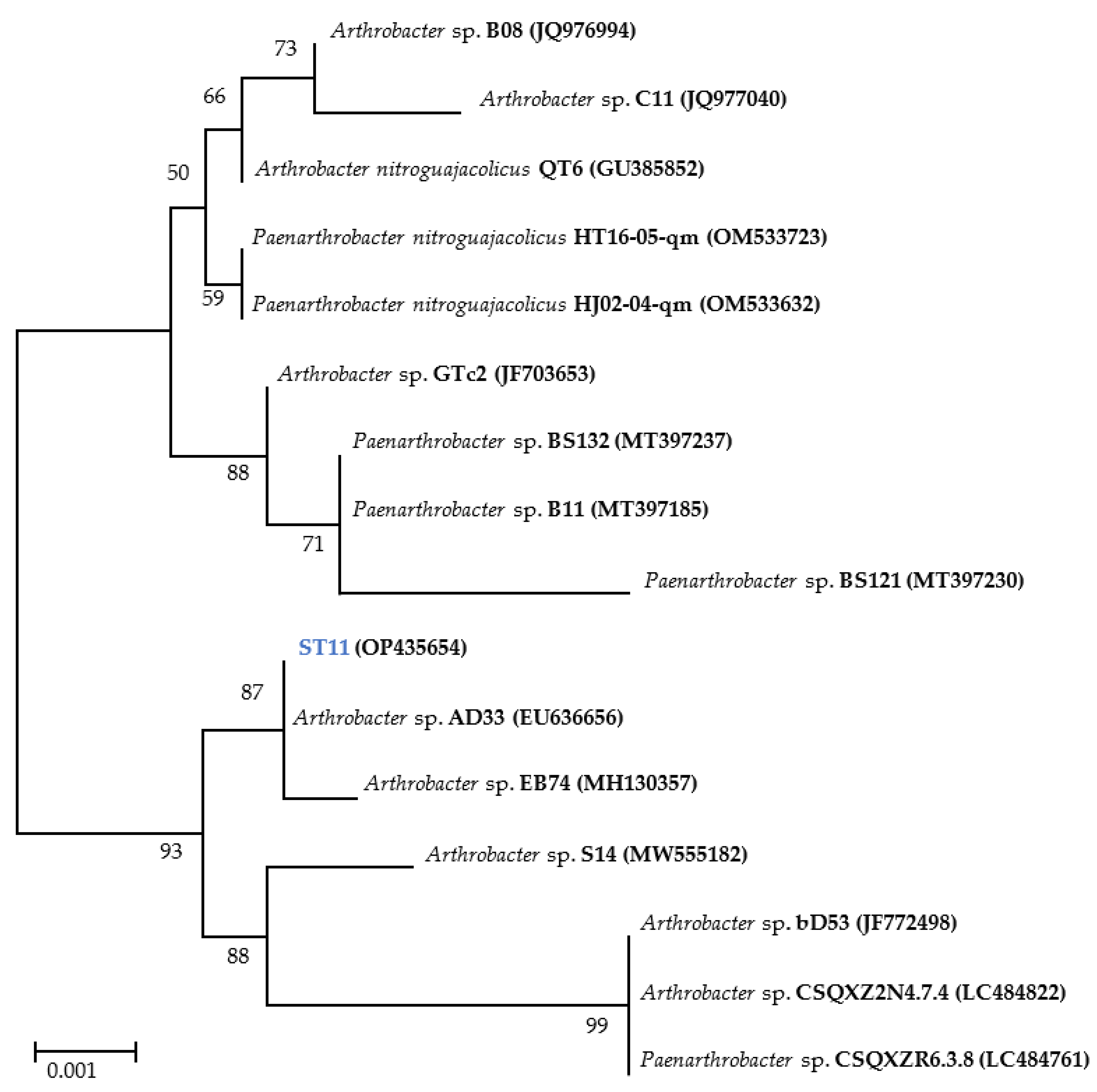
2.1. Identification and Characterization of Test Strain
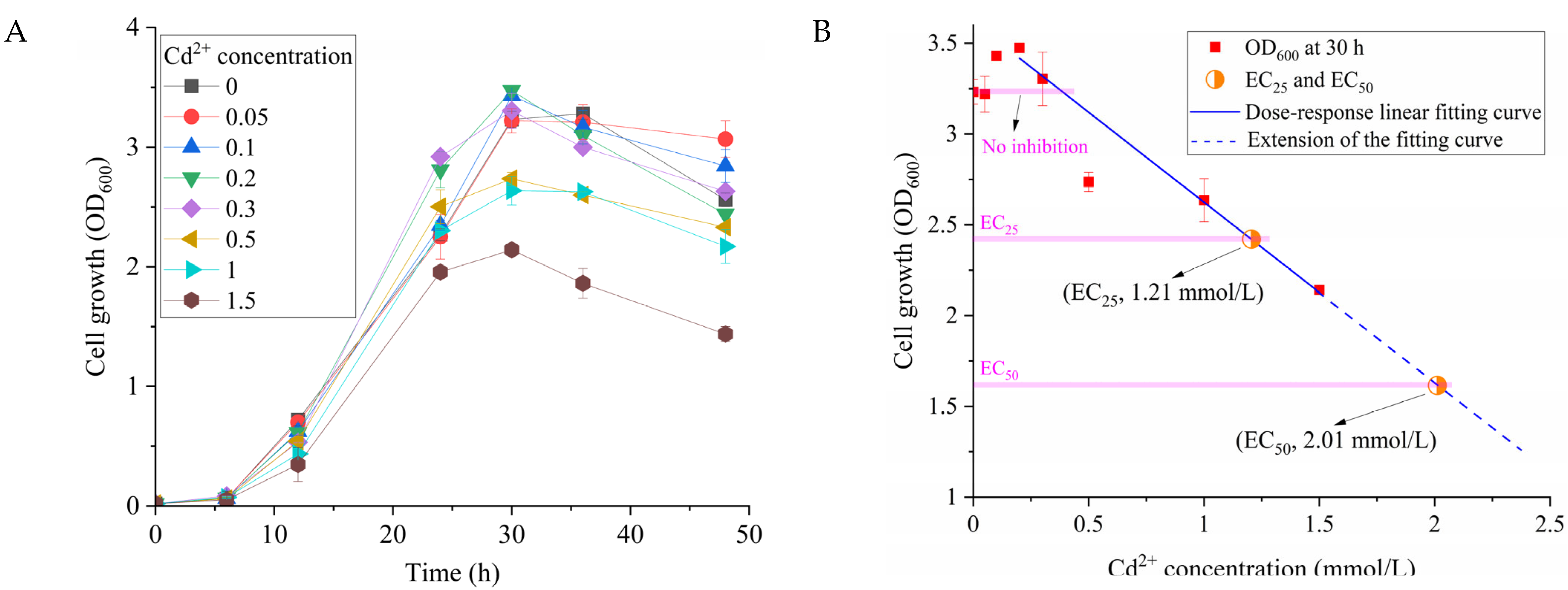
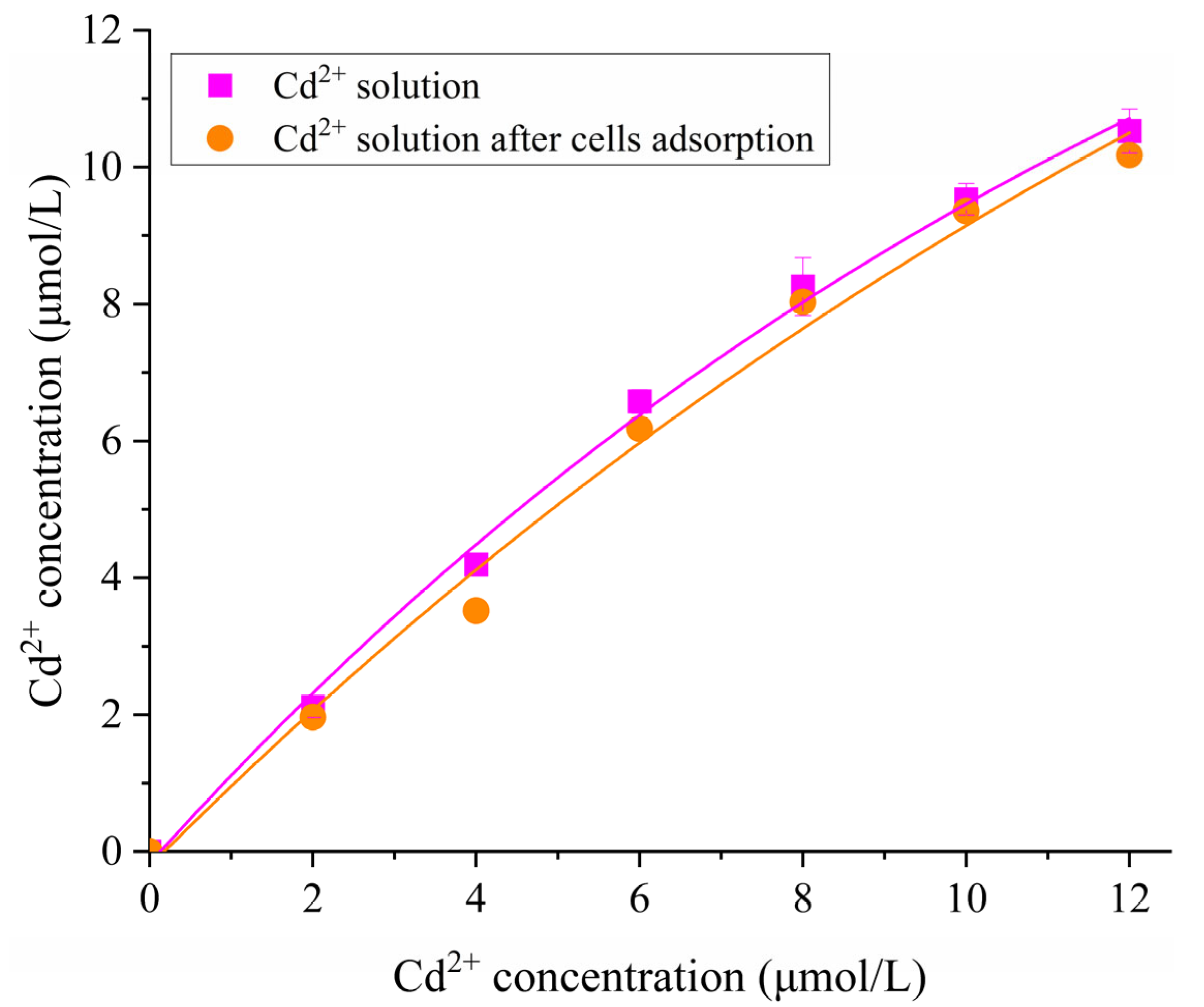
2.2. Characterization of Cd2+ Resistance
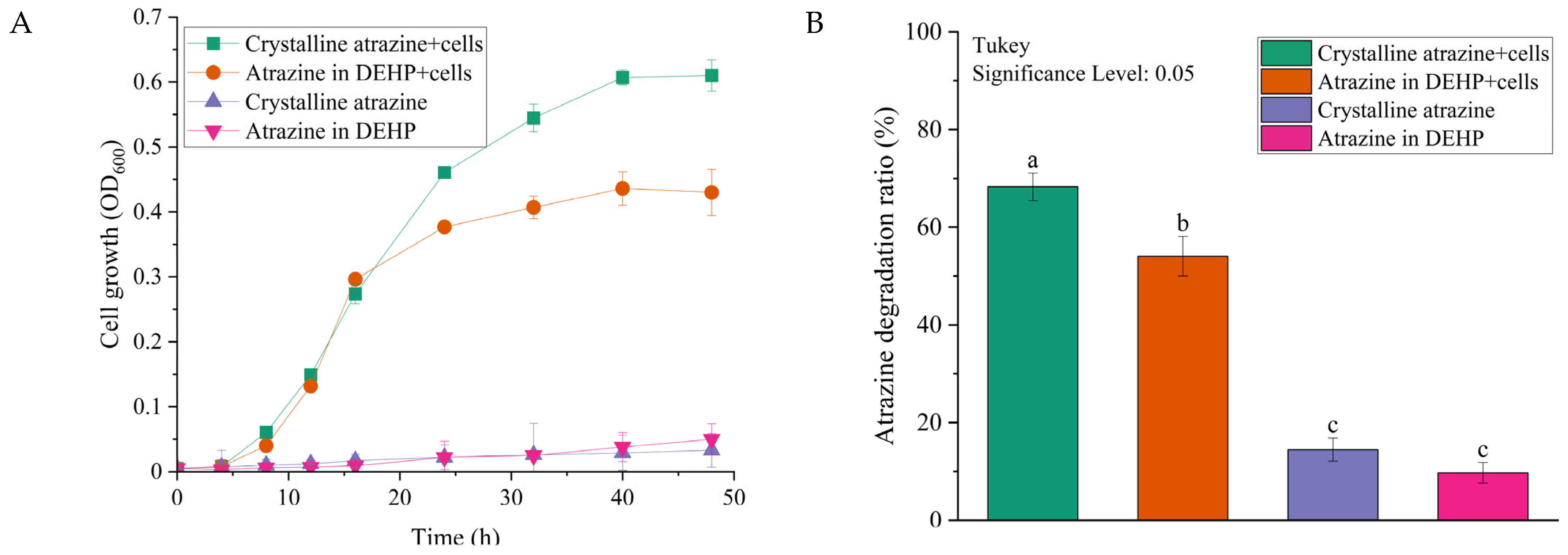
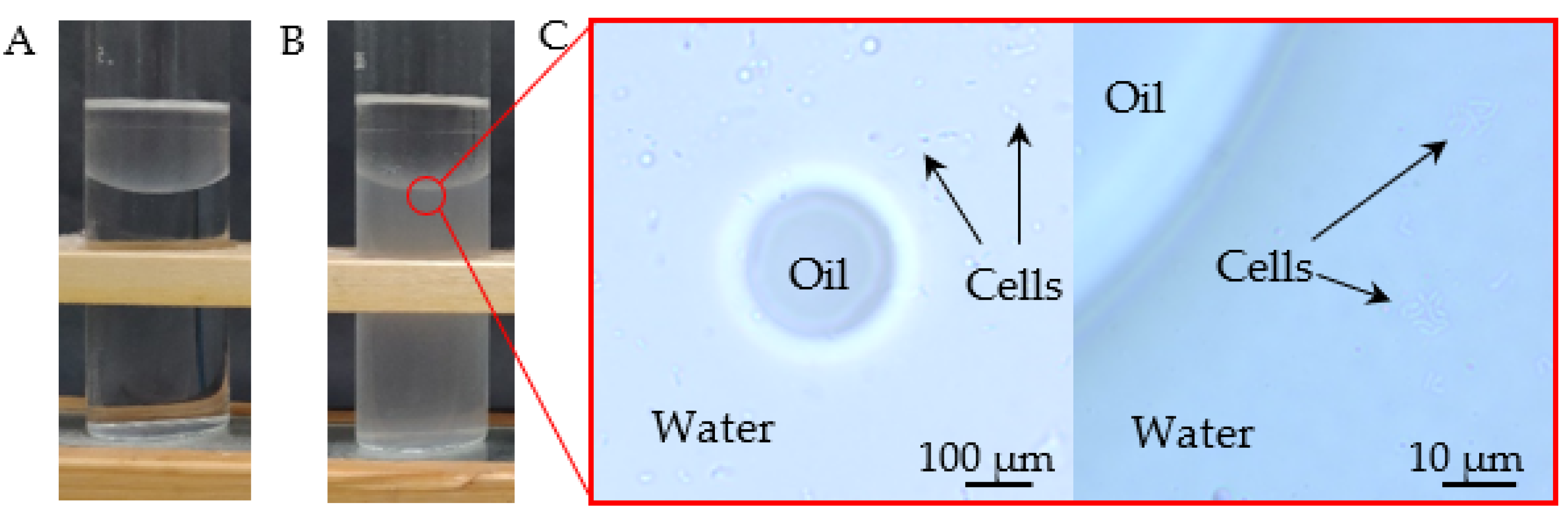
2.3. Biodegradation of Crystalline and NAPL-Dissolved Atrazine
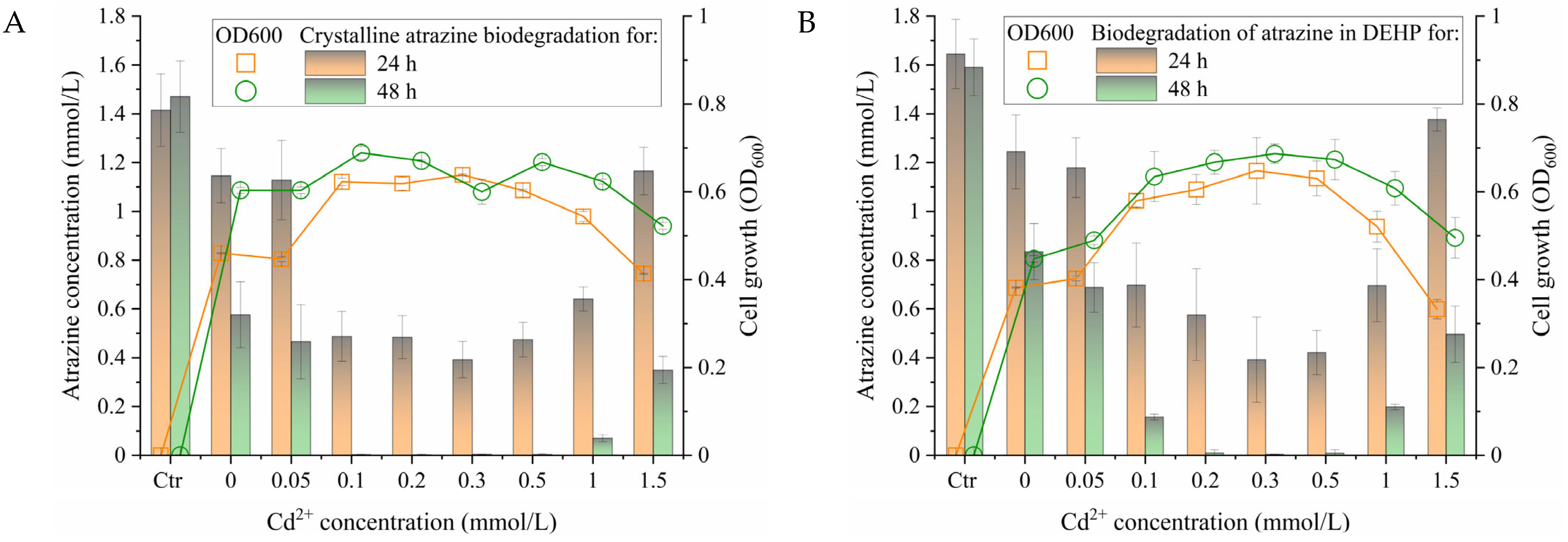
2.4. Effect of Cd2+ on the Growth of Strain ST11 and Atrazine Biodegradation
3. Materials and Methods
3.1. Chemicals
3.2. Microorganism and Culture Conditions
3.3. Cd2+ Resistance and Growth Curve of ST11
3.4. Cd2+ Adsorption Experiment
3.5. Biodegradation of Crystalline Atrazine
3.6. Biodegradation of Atrazine in NAPL
3.7. Effect of Cd2+ Ions
3.8. Analytical Methods
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wołejko, E.; Wydro, U.; Odziejewicz, J.I.; Koronkiewicz, A.; Jabłońska-Trypuć, A. Biomonitoring of Soil Contaminated with Herbicides. Water 2022, 14, 1534. [Google Scholar] [CrossRef]
- Rostami, S.; Jafari, S.; Moeini, Z.; Jaskulak, M.; Keshtgar, L.; Badeenezhad, A.; Azhdarpoor, A.; Rostami, M.; Zorena, K.; Dehghani, M. Current Methods and Technologies for Degradation of Atrazine in Contaminated Soil and Water: A Review. Environ. Technol. Innov. 2021, 24, 102019. [Google Scholar] [CrossRef]
- Jowa, L.; Howd, R. Should Atrazine and Related Chlorotriazines Be Considered Carcinogenic for Human Health Risk Assessment? J. Environ. Sci. Health Part C Environ. Carcinog. Ecotoxicol. Rev. 2011, 29, 91–144. [Google Scholar] [CrossRef] [PubMed]
- Mudhoo, A.; Garg, V.K. Sorption, Transport and Transformation of Atrazine in Soils, Minerals and Composts: A Review. Pedosphere 2011, 21, 11–25. [Google Scholar] [CrossRef]
- Vig, K.; Megharaj, M.; Sethunathan, N.; Naidu, R. Bioavailability and Toxicity of Cadmium to Microorganisms and Their Activities in Soil: A Review. Adv. Environ. Res. 2003, 8, 121–135. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, D.; Zhong, T.; Zhang, X.; Cheng, M.; Li, X. Assessment of Cadmium (Cd) Concentration in Arable Soil in China. Environ. Sci. Pollut. Res. 2015, 22, 4932–4941. [Google Scholar] [CrossRef]
- Genchi, G.; Sinicropi, M.S.; Lauria, G.; Carocci, A.; Catalano, A. The Effects of Cadmium Toxicity. Int. J. Environ. Res. Public. Health 2020, 17, 3782. [Google Scholar] [CrossRef]
- Zhu, J.; Fu, L.; Jin, C.; Meng, Z.; Yang, N. Study on the Isolation of Two Atrazine-Degrading Bacteria and the Development of a Microbial Agent. Microorganisms 2019, 7, 80. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Liang, S.; Wang, X.; Lu, Z.; Sun, P.; Zhang, H.; Sun, F. Biodegradation of Atrazine by the Novel Klebsiella Variicola Strain FH-1. Biomed Res. Int. 2019, 2019, 4756579. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Xie, C.; Wang, L.; Li, Q.; Wang, Y. Biodegradation of Persistent Environmental Pollutants by Arthrobacter sp. Environ. Sci. Pollut. Res. 2019, 26, 8429–8443. [Google Scholar] [CrossRef]
- Fang, H.; Lian, J.; Wang, H.; Cai, L.; Yu, Y. Exploring Bacterial Community Structure and Function Associated with Atrazine Biodegradation in Repeatedly Treated Soils. J. Hazard. Mater. 2015, 286, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Zhang, J.; Gao, J.; Li, H.; Liang, D.; Liu, R. Isolation and Characterization of Atrazine-Degrading Strain Shewanella sp. YJY4 from Cornfield Soil. Lett. Appl. Microbiol. 2016, 63, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Kolekar, P.D.; Patil, S.M.; Suryavanshi, M.; Suryawanshi, S.S.; Khandare, R.; Govindwar, S.P.; Jadhav, J.P. Microcosm Study of Atrazine Bioremediation by Indigenous Microorganisms and Cytotoxicity of Biodegraded Metabolites. J. Hazard. Mater. 2019, 374, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Radwan, E.K.; El Sebai, T.N.M.; Ghafar, H.H.A.; Khattab, A.E.-N.A. Atrazine Mineralization by Stenotrophomonas Maltophilia and Agrobacterium Tumefaciens Egyptian Soil Isolates. Desalination Water Treat. 2019, 171, 325–330. [Google Scholar] [CrossRef]
- Sharma, A.; Kalyani, P.; Trivedi, V.D.; Kapley, A.; Phale, P.S. Nitrogen-Dependent Induction of Atrazine Degradation Pathway in Pseudomonas sp. Strain AKN5. FEMS Microbiol. Lett. 2019, 366, fny277. [Google Scholar] [CrossRef]
- Chen, S.; Li, Y.; Fan, Z.; Liu, F.; Liu, H.; Wang, L.; Wu, H. Soil Bacterial Community Dynamics Following Bioaugmentation with Paenarthrobacter sp. W11 in Atrazine-Contaminated Soil. Chemosphere 2021, 282, 130976. [Google Scholar] [CrossRef]
- Castro-Gutiérrez, V.; Masís-Mora, M.; Carazo-Rojas, E.; Mora-López, M.; Rodríguez-Rodríguez, C.E. Fungal and Bacterial Co-Bioaugmentation of a Pesticide-Degrading Biomixture: Pesticide Removal and Community Structure Variations during Different Treatments. Water. Air. Soil Pollut. 2019, 230, 247. [Google Scholar] [CrossRef]
- Lopes, P.R.M.; Cruz, V.H.; de Menezes, A.B.; Gadanhoto, B.P.; Moreira, B.R.D.A.; Mendes, C.R.; Mazzeo, D.E.C.; Dilarri, G.; Montagnolli, R.N. Microbial Bioremediation of Pesticides in Agricultural Soils: An Integrative Review on Natural Attenuation, Bioaugmentation and Biostimulation. Rev. Environ. Sci. Biotechnol. 2022, 21, 851–876. [Google Scholar] [CrossRef]
- Benson, J.J.; Sakkos, J.K.; Radian, A.; Wackett, L.P.; Aksan, A. Enhanced Biodegradation of Atrazine by Bacteria Encapsulated in Organically Modified Silica Gels. J. Colloid Interface Sci. 2018, 510, 57–68. [Google Scholar] [CrossRef]
- Jiang, Z.; Zhang, X.; Wang, Z.; Cao, B.; Deng, S.; Bi, M.; Zhang, Y. Enhanced Biodegradation of Atrazine by Arthrobacter sp. DNS10 during Co-Culture with a Phosphorus Solubilizing Bacteria: Enterobacter sp. P1. Ecotoxicol. Environ. Saf. 2019, 172, 159–166. [Google Scholar] [CrossRef]
- Jakinala, P.; Lingampally, N.; Kyama, A.; Hameeda, B. Enhancement of Atrazine Biodegradation by Marine Isolate Bacillus Velezensis MHNK1 in Presence of Surfactin Lipopeptide. Ecotoxicol. Environ. Saf. 2019, 182, 109372. [Google Scholar] [CrossRef] [PubMed]
- Meite, F.; Granet, M.; Imfeld, G. Ageing of Copper, Zinc and Synthetic Pesticides in Particle-Size and Chemical Fractions of Agricultural Soils. Sci. Total Environ. 2022, 824, 153860. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Fan, Z.; Zhao, J.; Dai, Z.; Zhao, Y.; Dai, Y. Copper Stimulates Neonicotinoid Insecticide Thiacloprid Degradation by Ensifer Adhaerens TMX-23. J. Appl. Microbiol. 2021, 131, 2838–2848. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Yao, G.; Jing, X.; Liu, X.; Liu, D.; Zhou, Z. Effects of Cd2+ and Pb2+ on Enantioselective Degradation Behavior of α-Cypermethrin in Soils and Their Combined Effect on Activities of Soil Enzymes. Environ. Sci. Pollut. Res. 2021, 28, 47099–47106. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-H.; Zhu, L.-S.; Meng, Y.; Wang, J.; Xie, H.; Zhang, Q.-M. The Combined Stress Effects of Atrazine and Cadmium on the Earthworm Eisenia Fetida. Environ. Toxicol. Chem. 2012, 31, 2035–2040. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Xie, D.; Peng, L.; Chen, C.; Li, C.; Que, X. Phytotoxicity of Atrazine Combined with Cadmium on Photosynthetic Apparatus of the Emergent Plant Species Iris Pseudacorus. Environ. Sci. Pollut. Res. 2022, 29, 34798–34812. [Google Scholar] [CrossRef]
- Lee, G.L.Y.; Zakaria, N.N.; Futamata, H.; Suzuki, K.; Zulkharnain, A.; Shaharuddin, N.A.; Convey, P.; Zahri, K.N.M.; Ahmad, S.A. Metabolic Pathway of Phenol Degradation of a Cold-Adapted Antarctic Bacteria, Arthrobacter sp. Catalysts 2022, 12, 1422. [Google Scholar] [CrossRef]
- Zhang, Y.; Ge, S.; Jiang, M.; Jiang, Z.; Wang, Z.; Ma, B. Combined Bioremediation of Atrazine-Contaminated Soil by Pennisetum and Arthrobacter sp. Strain DNS10. Environ. Sci. Pollut. Res. 2014, 21, 6234–6238. [Google Scholar] [CrossRef]
- Li, Y.; Liang, D.; Sha, J.; Zhang, J.; Gao, J.; Li, H.; Liu, R. Isolating and Identifying the Atrazine-Degrading Strain Arthrobacter sp. LY-1 and Applying It for the Bioremediation of Atrazine-Contaminated Soil. Pol. J. Environ. Stud. 2019, 28, 1267–1275. [Google Scholar] [CrossRef]
- Kundu, K.; Marozava, S.; Ehrl, B.; Merl-Pham, J.; Griebler, C.; Elsner, M. Defining Lower Limits of Biodegradation: Atrazine Degradation Regulated by Mass Transfer and Maintenance Demand in Arthrobacter Aurescens TC1. Isme J. 2019, 13, 2236–2251. [Google Scholar] [CrossRef]
- Niewerth, H.; Schuldes, J.; Parschat, K.; Kiefer, P.; Vorholt, J.A.; Daniel, R.; Fetzner, S. Complete Genome Sequence and Metabolic Potential of the Quinaldine-Degrading Bacterium Arthrobacter sp. Rue61a. BMC Genomics 2012, 13, 534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Shi, W.; Zeng, X.-C.; Yang, Y.; Zhou, L.; Mu, Y.; Liu, Y. Draft Genome Sequence of Arthrobacter sp. Strain B6 Isolated from the High-Arsenic Sediments in Datong Basin, China. Stand. Genomic Sci. 2017, 12, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luscombe, B.M.; Gray, T.R.G. Characteristics of Arthrobacter Grown in Continuous Culture. Microbiology 1974, 82, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Junco, M.; De Olmedo, E.; Ortega-Calvo, J.J. Bioavailability of Solid and Non-Aqueous Phase Liquid (NAPL)-Dissolved Phenanthrene to the Biosurfactant-Producing Bacterium Pseudomonas Aeruginosa 19SJ. Environ. Microbiol. 2001, 3, 561–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, H.; Liu, Y.; You, S.; Liu, J.; Xiao, H.; Tu, Z. A Review on Recent Treatment Technology for Herbicide Atrazine in Contaminated Environment. Int. J. Environ. Res. Public. Health 2019, 16, 5129. [Google Scholar] [CrossRef] [Green Version]
- Nemeth-Konda, L.; Füleky, G.; Morovjan, G.; Csokan, P. Sorption Behaviour of Acetochlor, Atrazine, Carbendazim, Diazinon, Imidacloprid and Isoproturon on Hungarian Agricultural Soil. Chemosphere 2002, 48, 545–552. [Google Scholar] [CrossRef]
- Pan, T.; Liu, C.; Wang, M.; Zhang, J. Interfacial Biodegradation of Phenanthrene in Bacteria-Carboxymethyl Cellulose-Stabilized Pickering Emulsions. Appl. Microbiol. Biotechnol. 2022, 106, 3829–3836. [Google Scholar] [CrossRef]
- Xu, X.; Zarecki, R.; Medina, S.; Ofaim, S.; Liu, X.; Chen, C.; Hu, S.; Brom, D.; Gat, D.; Porob, S.; et al. Modeling Microbial Communities from Atrazine Contaminated Soils Promotes the Development of Biostimulation Solutions. ISME J. 2019, 13, 494–508. [Google Scholar] [CrossRef] [Green Version]
- Ortega-Calvo, J.J.; Alexander, M. Roles of Bacterial Attachment and Spontaneous Partitioning in the Biodegradation of Naphthalene Initially Present in Nonaqueous-Phase Liquids. Appl. Environ. Microbiol. 1994, 60, 2643–2646. [Google Scholar] [CrossRef] [Green Version]
- Pal, A.; Bhattacharjee, S.; Saha, J.; Sarkar, M.; Mandal, P. Bacterial Survival Strategies and Responses under Heavy Metal Stress: A Comprehensive Overview. Crit. Rev. Microbiol. 2022, 48, 327–355. [Google Scholar] [CrossRef]
- Chen, S.; Yin, H.; Ye, J.; Peng, H.; Zhang, N.; He, B. Effect of Copper(II) on Biodegradation of Benzo[a]Pyrene by Stenotrophomonas Maltophilia. Chemosphere 2013, 90, 1811–1820. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Zhang, Z.-Z.; Wu, X.-J.; Xu, Y.-X.; Su, X.-L.; Zhang, M.; Wang, J.-X. Biodegradation of Decabromodiphenyl Ether (BDE-209) by a Metal Resistant Strain, Bacillus Cereus JP12. Bioresour. Technol. 2013, 149, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Helbig, K.; Grosse, C.; Nies, D.H. Cadmium Toxicity in Glutathione Mutants of Escherichia coli. J. Bacteriol. 2008, 190, 5439–5454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruins, M.R.; Kapil, S.; Oehme, F.W. Microbial Resistance to Metals in the Environment. Ecotoxicol. Environ. Saf. 2000, 45, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Elyamine, A.M.; Afzal, J.; Rana, M.S.; Imran, M.; Cai, M.; Hu, C. Phenanthrene Mitigates Cadmium Toxicity in Earthworms Eisenia Fetida (Epigeic Specie) and Aporrectodea Caliginosa (Endogeic Specie) in Soil. Int. J. Environ. Res. Public. Health 2018, 15, 2384. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Wang, M.; Zhang, J.; Qian, Y.; Xiao, K.; Wang, R.; Dong, W.; Pan, T. A Polycyclic Aromatic Hydrocarbon Degrading Strain and Its Potential to Degrade Phenanthrene in Various Enhanced Systems. Chin. J. Biotechnol. 2021, 37, 3696–3707. [Google Scholar]
- Pan, T.; Deng, T.; Zeng, X.; Dong, W.; Yu, S. Extractive Biodegradation and Bioavailability Assessment of Phenanthrene in the Cloud Point System by Sphingomonas Polyaromaticivorans. Appl. Microbiol. Biotechnol. 2016, 100, 431–437. [Google Scholar] [CrossRef]
- Rosenberg, M. Microbial Adhesion to Hydrocarbons: Twenty-Five Years of Doing MATH. FEMS Microbiol. Lett. 2006, 262, 129–134. [Google Scholar] [CrossRef]
- Wang, J.; Ren, Z.; Wu, L.; Li, M. Determination of Trace Cd in Water by Spectrophotometry after Cloud Point Extraction. Phys. Test. Chem. Anal. Part B Chem. Anal. 2012, 48, 735–736, 739. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Yu, Z.; Gao, Y.; Wang, M.; Wang, K.; Pan, T. Biodegradation of Crystalline and Nonaqueous Phase Liquid-Dissolved ATRAZINE by Arthrobacter sp. ST11 with Cd2+ Resistance. Catalysts 2022, 12, 1653. https://doi.org/10.3390/catal12121653
Zhang J, Yu Z, Gao Y, Wang M, Wang K, Pan T. Biodegradation of Crystalline and Nonaqueous Phase Liquid-Dissolved ATRAZINE by Arthrobacter sp. ST11 with Cd2+ Resistance. Catalysts. 2022; 12(12):1653. https://doi.org/10.3390/catal12121653
Chicago/Turabian StyleZhang, Jiameng, Zhiliang Yu, Yaling Gao, Meini Wang, Kai Wang, and Tao Pan. 2022. "Biodegradation of Crystalline and Nonaqueous Phase Liquid-Dissolved ATRAZINE by Arthrobacter sp. ST11 with Cd2+ Resistance" Catalysts 12, no. 12: 1653. https://doi.org/10.3390/catal12121653