Beyond Angiogenesis: The Multitasking Approach of the First PEGylated Vascular Endothelial Growth Factor (CdtVEGF) from Brazilian Rattlesnake Venom

 ,
,  , ,
, ,  , ,
, ,  ,
,  ,
,
Abstract
:1. Introduction
2. Results
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Venoms, Cells, and Animals
5.2. CdtVEGF Isolation
5.3. Recognition of CdtVEGF and PEG-CdtVEGF Using Enzyme-Linked Immunosorbent Assay (ELISA)
5.4. SDS-PAGE
5.5. Reactivity of Crotalid Antivenom towards CdtVEGF and Prediction of Potential Epitopes within the Molecule
5.6. Molecular Mass of CdtVEGF
5.7. Amino Acid Sequence Determination of CdtVEGF
5.8. CdtVEGF Homology Model
5.9. Bioconjugation of CdtVEGF with mPEG-Maleimide
5.10. In Vitro Assays
5.10.1. Angiogenesis
5.10.2. Metabolic Activity
5.10.3. Wound Healing Assay
5.11. In Vivo Assays
5.11.1. Evaluation of Leukocyte Influx into the Peritoneal Cavity
5.11.2. Protein Quantification
5.11.3. Cytokine Measurement
5.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lawrence, P.B.; Price, J.L. How PEGylation Influences Protein Conformational Stability. Curr. Opin. Chem. Biol. 2016, 34, 88. [Google Scholar] [CrossRef] [Green Version]
- Gefen, T.; Vaya, J.; Khatib, S.; Harkevich, N.; Artoul, F.; Heller, E.D.; Pitcovski, J.; Aizenshtein, E. The Impact of PEGylation on Protein Immunogenicity. Int. Immunopharmacol. 2013, 15, 254–259. [Google Scholar] [CrossRef]
- Harris, J.M.; Chess, R.B. Effect of Pegylation on Pharmaceuticals. Nat. Rev. Drug Discov. 2003, 2, 214–221. [Google Scholar] [CrossRef]
- Mullard, A. 2021 FDA Approvals. Nat. Rev. Drug Discov. 2022, 21, 83–88. [Google Scholar] [CrossRef]
- Hiu, J.J.; Yap, M.K.K. Cytotoxicity of Snake Venom Enzymatic Toxins: Phospholipase A2 and l-Amino Acid Oxidase. Biochem. Soc. Trans. 2020, 48, 719–731. [Google Scholar] [CrossRef] [Green Version]
- Olaoba, O.T.; Karina Dos Santos, P.; Selistre-de-Araujo, H.S.; Ferreira de Souza, D.H. Snake Venom Metalloproteinases (SVMPs): A Structure-Function Update. Toxicon X 2020, 7, 100052. [Google Scholar] [CrossRef]
- Péterfi, O.; Boda, F.; Szabó, Z.; Ferencz, E.; Bába, L. Hypotensive Snake Venom Components-A Mini-Review. Molecules 2019, 24, 2778. [Google Scholar] [CrossRef] [Green Version]
- Jj, C. The Continuing Saga of Snake Venom Disintegrins. Toxicon Off. J. Int. Soc. Toxinol. 2013, 62, 40–49. [Google Scholar] [CrossRef]
- Ja, E. Structurally Robust and Functionally Highly Versatile-C-Type Lectin (-Related) Proteins in Snake Venoms. Toxins 2019, 11, 136. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, I.G.; Pucca, M.B.; de Oliveira, I.S.; Cerni, F.A.; da Silva Jacob, B.D.C.; Arantes, E.C. Snake Venom Vascular Endothelial Growth Factors (SvVEGFs): Unravelling Their Molecular Structure, Functions, and Research Potential. Cytokine Growth Factor Rev. 2021, 60, 133–143. [Google Scholar] [CrossRef]
- Bordon, K.d.C.F.; Cologna, C.T.; Fornari-Baldo, E.C.; Pinheiro-Júnior, E.L.; Cerni, F.A.; Amorim, F.G.; Anjolette, F.A.P.; Cordeiro, F.A.; Wiezel, G.A.; Cardoso, I.A.; et al. From Animal Poisons and Venoms to Medicines: Achievements, Challenges and Perspectives in Drug Discovery. Front. Pharmacol. 2020, 11, 1132. [Google Scholar] [CrossRef] [PubMed]
- Tadokoro, T.; Modahl, C.M.; Maenaka, K.; Aoki-Shioi, N. Cysteine-Rich Secretory Proteins (CRISPs) From Venomous Snakes: An Overview of the Functional Diversity in A Large and Underappreciated Superfamily. Toxins 2020, 12, 175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Rabies and Envenomings: A Neglected Public Health Issue: Report of a Consultative Meeting; World Health Organization: Geneva, Switzerland, 2007. [Google Scholar]
- Saúde, Ministério da Ministerio Da Saude, Acidentes Por Animais Peçonhentos–Notificações Registradas No Sistema de Informação de Agravos de Notificação, Brazil. 2021. Available online: http://tabnet.datasus.gov.br/cgi/tabcgi.exe?sinannet/cnv/animaisbr.def (accessed on 24 January 2023).
- Duarte, M.R.; Menezes, F.A. Is the Population of Crotalus Durissus (Serpentes, Viperidae) Expanding in Brazil? J. Venom. Anim. Toxins Incl. Trop. Dis. 2013, 19, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiezel, G.A.; Shibao, P.Y.T.; Cologna, C.T.; Morandi Filho, R.; Ueira-Vieira, C.; De Pauw, E.; Quinton, L.; Arantes, E.C. In-Depth Venome of the Brazilian Rattlesnake Crotalus Durissus Terrificus: An Integrative Approach Combining Its Venom Gland Transcriptome and Venom Proteome. J. Proteome Res. 2018, 17, 3941–3958. [Google Scholar] [CrossRef]
- Melincovici, C.S.; Boşca, A.B.; Şuşman, S.; Mărginean, M.; Mihu, C.; Istrate, M.; Moldovan, I.M.; Roman, A.L.; Mihu, C.M. Vascular Endothelial Growth Factor (VEGF)–Key Factor in Normal and Pathological Angiogenesis. Rom. J. Morphol. Embryol. 2018, 59, 455–467. [Google Scholar]
- Junqueira de Azevedo, I.L.; Farsky, S.H.; Oliveira, M.L.; Ho, P.L. Molecular Cloning and Expression of a Functional Snake Venom Vascular Endothelium Growth Factor (VEGF) from the Bothrops Insularis Pit Viper. A New Member of the VEGF Family of Proteins. J. Biol. Chem. 2001, 276, 39836–39842. [Google Scholar] [CrossRef] [Green Version]
- Suto, K.; Yamazaki, Y.; Morita, T.; Mizuno, H. Crystal Structures of Novel Vascular Endothelial Growth Factors (VEGF) from Snake Venoms: Insight into Selective VEGF Binding to Kinase Insert Domain-Containing Receptor but Not to Fms-like Tyrosine Kinase-1. J. Biol. Chem. 2005, 280, 2126–2131. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, Y.; Takani, K.; Atoda, H.; Morita, T. Snake Venom Vascular Endothelial Growth Factors (VEGFs) Exhibit Potent Activity through Their Specific Recognition of KDR (VEGF Receptor 2). J. Biol. Chem. 2003, 278, 51985–51988. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, Y.; Matsunaga, Y.; Nakano, Y.; Morita, T. Identification of Vascular Endothelial Growth Factor Receptor-Binding Protein in the Venom of Eastern Cottonmouth. A New Role of Snake Venom Myotoxic Lys49-Phospholipase A2. J. Biol. Chem. 2005, 280, 29989–29992. [Google Scholar] [CrossRef] [Green Version]
- De Vries, C.; Escobedo, J.A.; Ueno, H.; Houck, K.; Ferrara, N.; Williams, L.T. The Fms-like Tyrosine Kinase, a Receptor for Vascular Endothelial Growth Factor. Science 1992, 255, 989–991. [Google Scholar] [CrossRef]
- Terman, B.I.; Dougher-Vermazen, M.; Carrion, M.E.; Dimitrov, D.; Armellino, D.C.; Gospodarowicz, D.; Böhlen, P. Identification of the KDR Tyrosine Kinase as a Receptor for Vascular Endothelial Cell Growth Factor. Biochem. Biophys. Res. Commun. 1992, 187, 1579–1586. [Google Scholar] [CrossRef]
- Pajusola, K.; Aprelikova, O.; Pelicci, G.; Weich, H.; Claesson-Welsh, L.; Alitalo, K. Signalling Properties of FLT4, a Proteolytically Processed Receptor Tyrosine Kinase Related to Two VEGF Receptors. Oncogene 1994, 9, 3545–3555. [Google Scholar] [PubMed]
- Yamazaki, Y.; Morita, T. Molecular and Functional Diversity of Vascular Endothelial Growth Factors. Mol. Divers. 2006, 10, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.; Wu, J.; Cui, Y.; Li, R.; Zhu, S.; Rong, M.; Lu, Q.; Lai, R. Vascular Endothelial Growth Factor from Trimeresurus Jerdonii Venom Specifically Binds to VEGFR-2. Biochimie 2015, 116, 1–7. [Google Scholar] [CrossRef]
- Yamazaki, Y.; Matsunaga, Y.; Tokunaga, Y.; Obayashi, S.; Saito, M.; Morita, T. Snake Venom Vascular Endothelial Growth Factors (VEGF-Fs) Exclusively Vary Their Structures and Functions among Species. J. Biol. Chem. 2009, 284, 9885–9891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, I.G.; Pucca, M.B.; Cardoso, I.A.; de Castro Figueiredo Bordon, K.; Wiezel, G.A.; Amorim, F.G.; Rodrigues, R.S.; de Melo Rodrigues, V.; Lucia de Campos Brites, V.; Rosa, J.C.; et al. Insights into Structure and Function of CdcVEGFs, the Vascular Endothelial Growth Factor from Crotalus durissus collilineatus Snake Venom. Biochimie 2022, 200, 68–78. [Google Scholar] [CrossRef]
- Messadi, E.; Aloui, Z.; Belaidi, E.; Vincent, M.-P.; Couture-Lepetit, E.; Waeckel, L.; Decorps, J.; Bouby, N.; Gasmi, A.; Karoui, H.; et al. Cardioprotective Effect of VEGF and Venom VEGF-like Protein in Acute Myocardial Ischemia in Mice: Effect on Mitochondrial Function. J. Cardiovasc. Pharmacol. 2014, 63, 274–281. [Google Scholar] [CrossRef]
- Matsunaga, Y.; Yamazaki, Y.; Suzuki, H.; Morita, T. VEGF-A and VEGF-F Evoke Distinct Changes in Vascular Ultrastructure. Biochem. Biophys. Res. Commun. 2009, 379, 872–875. [Google Scholar] [CrossRef]
- Nakamura, H.; Murakami, T.; Imamura, T.; Toriba, M.; Chijiwa, T.; Ohno, M.; Oda-Ueda, N. Discovery of a Novel Vascular Endothelial Growth Factor (VEGF) with No Affinity to Heparin in Gloydius tsushimaensis Venom. Toxicon 2014, 86, 107–115. [Google Scholar] [CrossRef]
- Zittermann, S.I.; Issekutz, A.C. Endothelial Growth Factors VEGF and BFGF Differentially Enhance Monocyte and Neutrophil Recruitment to Inflammation. J. Leukoc. Biol. 2006, 80, 247–257. [Google Scholar] [CrossRef] [Green Version]
- Melani, R.; Gabriel, D.; Carvalho, P.; Goto-Silva, L.; Nogueira, F.; Junqueira, M.; Domont, G. Seeing beyond the Tip of the Iceberg: A Deep Analysis of the Venome of the Brazilian Rattlesnake, Crotalus Durissus Terrificus. EuPA Open Proteom. 2015, 8, 144–156. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, I.S.; Cardoso, I.A.; Bordon, K.d.C.F.; Carone, S.E.I.; Boldrini-França, J.; Pucca, M.B.; Zoccal, K.F.; Faccioli, L.H.; Sampaio, S.V.; Rosa, J.C.; et al. Global Proteomic and Functional Analysis of Crotalus Durissus Collilineatus Individual Venom Variation and Its Impact on Envenoming. J. Proteom. 2019, 191, 153–165. [Google Scholar] [CrossRef]
- Robert, X.; Gouet, P. Deciphering Key Features in Protein Structures with the New ENDscript Server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef] [Green Version]
- Komori, Y.; Nikai, T.; Taniguchi, K.; Masuda, K.; Sugihara, H. Vascular Endothelial Growth Factor VEGF-like Heparin-Binding Protein from the Venom of Vipera aspis aspis (Aspic Viper). Biochemistry 1999, 38, 11796–11803. [Google Scholar] [CrossRef]
- de Junqueira-de-Azevedo, I.L.M.; da Silva, M.B.; Chudzinski-Tavassi, A.M.; Ho, P.L. Identification and Cloning of Snake Venom Vascular Endothelial Growth Factor (SvVEGF) from Bothrops Erythromelas Pitviper. Toxicon 2004, 44, 571–575. [Google Scholar] [CrossRef] [PubMed]
- Bénard-Valle, M.; Neri-Castro, E.; Yañez-Mendoza, M.F.; Lomonte, B.; Olvera, A.; Zamudio, F.; Restano-Cassulini, R.; Possani, L.D.; Jiménez-Ferrer, E.; Alagón, A. Functional, Proteomic and Transcriptomic Characterization of the Venom from Micrurus browni browni: Identification of the First Lethal Multimeric Neurotoxin in Coral Snake Venom. J. Proteom. 2020, 225, 103863. [Google Scholar] [CrossRef] [PubMed]
- Boldrini-França, J.; Rodrigues, R.S.; Fonseca, F.P.P.; Menaldo, D.L.; Ferreira, F.B.; Henrique-Silva, F.; Soares, A.M.; Hamaguchi, A.; Rodrigues, V.M.; Otaviano, A.R.; et al. Crotalus Durissus Collilineatus Venom Gland Transcriptome: Analysis of Gene Expression Profile. Biochimie 2009, 91, 586–595. [Google Scholar] [CrossRef] [PubMed]
- Boldrini-França, J.; Corrêa-Netto, C.; Silva, M.M.S.; Rodrigues, R.S.; Nogueira, R.A.; Rodrigues, V.M.; Sanz, L.; Calvete, J.J. Snake Venomics and Antivenomics of Crotalus durissus Subspecies from Brazil: Assessment of Geographic Variation and Its Implication on Snakebite Management. J. Proteom. 2010, 73, 1758–1776. [Google Scholar] [CrossRef]
- Tasima, L.J.; Hatakeyama, D.M.; Serino-Silva, C.; Rodrigues, C.F.B.; de Lima, E.O.V.; Sant’Anna, S.S.; Grego, K.F.; de Morais-Zani, K.; Sanz, L.; Calvete, J.J.; et al. Comparative Proteomic Profiling and Functional Characterization of Venom Pooled from Captive Crotalus durissus terrificus Specimens and the Brazilian Crotalic Reference Venom. Toxicon 2020, 185, 26–35. [Google Scholar] [CrossRef]
- Aloui, Z.; Hoos, S.; Geretti, E.; Kharmachi, H.; Haumont, P.Y.; Mejdoub, H.; Klagsbrun, M.; England, P.; Gasmi, A. Novel SvVEGF Isoforms from Macrovipera Lebetina Venom Interact with Neuropilins. Biochem. Biophys. Res. Commun. 2009, 389, 10–15. [Google Scholar] [CrossRef]
- Boldrini-França, J.; Cologna, C.T.; Pucca, M.B.; de Bordon, K.C.F.; Amorim, F.G.; Anjolette, F.A.P.; Cordeiro, F.A.; Wiezel, G.A.; Cerni, F.A.; Pinheiro-Junior, E.L.; et al. Minor Snake Venom Proteins: Structure, Function and Potential Applications. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2017, 1861, 824–838. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-L.; Tsai, I.-H.; Hong, T.-M.; Tsai, S.-H. Crotalid Venom Vascular Endothelial Growth Factors Has Preferential Affinity for VEGFR-1. Characterization of Protobothrops mucrosquamatus Venom VEGF. Thromb. Haemost. 2005, 93, 331–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bremel, R.D.; Homan, E.J. An Integrated Approach to Epitope Analysis II: A System for Proteomic-Scale Prediction of Immunological Characteristics. Immunome Res. 2010, 6, 8. [Google Scholar] [CrossRef] [Green Version]
- Sivalingam, G.N.; Shepherd, A.J. An Analysis of B-Cell Epitope Discontinuity. Mol. Immunol. 2012, 51, 304–309. [Google Scholar] [CrossRef]
- da-Silva-Freitas, D.; Boldrini-França, J.; Arantes, E.C. PEGylation: A Successful Approach to Improve the Biopharmaceutical Potential of Snake Venom Thrombin-like Serine Protease. Protein Pept. Lett. 2015, 22, 1133–1139. [Google Scholar] [CrossRef]
- Pinheiro-Junior, E.L.; Boldrini-França, J.; Takeda, A.A.S.; Costa, T.R.; Peigneur, S.; Cardoso, I.A.; de Oliveira, I.S.; Sampaio, S.V.; de Mattos Fontes, M.R.; Tytgat, J.; et al. Towards Toxin PEGylation: The Example of RCollinein-1, a Snake Venom Thrombin-like Enzyme, as a PEGylated Biopharmaceutical Prototype. Int. J. Biol. Macromol. 2021, 190, 564–573. [Google Scholar] [CrossRef]
- Wan, X.; Zhang, J.; Yu, W.; Shen, L.; Ji, S.; Hu, T. Effect of Protein Immunogenicity and PEG Size and Branching on the Anti-PEG Immune Response to PEGylated Proteins. Process Biochem. 2017, 52, 183–191. [Google Scholar] [CrossRef]
- Hoang Thi, T.T.; Pilkington, E.H.; Nguyen, D.H.; Lee, J.S.; Park, K.D.; Truong, N.P. The Importance of Poly(Ethylene Glycol) Alternatives for Overcoming PEG Immunogenicity in Drug Delivery and Bioconjugation. Polymers 2020, 12, 298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurfürst, M.M. Detection and Molecular Weight Determination of Polyethylene Glycol-Modified Hirudin by Staining after Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis. Anal Biochem 1992, 200, 244–248. [Google Scholar] [CrossRef]
- Bailon, P.; Palleroni, A.; Schaffer, C.A.; Spence, C.L.; Fung, W.J.; Porter, J.E.; Ehrlich, G.K.; Pan, W.; Xu, Z.X.; Modi, M.W.; et al. Rational Design of a Potent, Long-Lasting Form of Interferon: A 40 KDa Branched Polyethylene Glycol-Conjugated Interferon Alpha-2a for the Treatment of Hepatitis C. Bioconjug Chem 2001, 12, 195–202. [Google Scholar] [CrossRef]
- Gurram, S.; Srivastava, G.; Badve, V.; Nandre, V.; Gundu, S.; Doshi, P. Pyridine Borane as Alternative Reducing Agent to Sodium Cyanoborohydride for the PEGylation of L-Asparaginase. Appl Biochem Biotechnol 2022, 194, 827–847. [Google Scholar] [CrossRef]
- Lee, H.; Jang, I.H.; Ryu, S.H.; Park, T.G. N-Terminal Site-Specific Mono-PEGylation of Epidermal Growth Factor. Pharm. Res. 2003, 20, 818–825. [Google Scholar] [CrossRef]
- Eming, S.A.; Krieg, T.; Davidson, J.M. Inflammation in Wound Repair: Molecular and Cellular Mechanisms. J. Investig. Dermatol. 2007, 127, 514–525. [Google Scholar] [CrossRef] [Green Version]
- Barrientos, S.; Brem, H.; Stojadinovic, O.; Tomic-Canic, M. Clinical Application of Growth Factors and Cytokines in Wound Healing. Wound Repair. Regen. 2014, 22, 569–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasmi, A.; Bourcier, C.; Aloui, Z.; Srairi, N.; Marchetti, S.; Gimond, C.; Wedge, S.R.; Hennequin, L.; Pouysségur, J. Complete Structure of an Increasing Capillary Permeability Protein (ICPP) Purified from Vipera lebetina Venom. ICPP Is Angiogenic via Vascular Endothelial Growth Factor Receptor Signalling. J. Biol. Chem. 2002, 277, 29992–29998. [Google Scholar] [CrossRef] [Green Version]
- Santos, J.H.P.M.; Torres-Obreque, K.M.; Meneguetti, G.P.; Amaro, B.P.; Rangel-Yagui, C.O. Protein PEGylation for the Design of Biobetters: From Reaction to Purification Processes. Braz. J. Pharm. Sci. 2018, 54, e01009. [Google Scholar] [CrossRef] [Green Version]
- Springer, T.A. Traffic Signals on Endothelium for Lymphocyte Recirculation and Leukocyte Emigration. Annu. Rev. Physiol. 1995, 57, 827–872. [Google Scholar] [CrossRef] [PubMed]
- Detmar, M.; Brown, L.F.; Schön, M.P.; Elicker, B.M.; Velasco, P.; Richard, L.; Fukumura, D.; Monsky, W.; Claffey, K.P.; Jain, R.K. Increased Microvascular Density and Enhanced Leukocyte Rolling and Adhesion in the Skin of VEGF Transgenic Mice. J. Investig. Dermatol. 1998, 111, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Dirkx, A.E.M.; Oude Egbrink, M.G.A.; Kuijpers, M.J.E.; van der Niet, S.T.; Heijnen, V.V.T.; Bouma-ter Steege, J.C.A.; Wagstaff, J.; Griffioen, A.W. Tumor Angiogenesis Modulates Leukocyte-Vessel Wall Interactions in Vivo by Reducing Endothelial Adhesion Molecule Expression. Cancer Res. 2003, 63, 2322–2329. [Google Scholar]
- Mortaz, E.; Alipoor, S.D.; Adcock, I.M.; Mumby, S.; Koenderman, L. Update on Neutrophil Function in Severe Inflammation. Front. Immunol. 2018, 9, 2171. [Google Scholar] [CrossRef] [Green Version]
- Santos, E.W.; de Oliveira, D.C.; Hastreiter, A.; Silva, G.B.d.; Beltran, J.S.d.O.; Tsujita, M.; Crisma, A.R.; Neves, S.M.P.; Fock, R.A.; Borelli, P. Hematological and Biochemical Reference Values for C57BL/6, Swiss Webster and BALB/c Mice. Braz. J. Vet. Res. Anim. Sci. 2016, 53, 138–145. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, H.; Hattori, S.; Iwamatsu, A.; Takizawa, H.; Shibuya, M. A Novel Snake Venom Vascular Endothelial Growth Factor (VEGF) Predominantly Induces Vascular Permeability through Preferential Signaling via VEGF Receptor-1. J. Biol. Chem. 2004, 279, 46304–46314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furie, M.B.; Randolph, G.J. Chemokines and Tissue Injury. Am. J. Pathol. 1995, 146, 1287–1301. [Google Scholar] [PubMed]
- Taibi-Djennah, Z.; Laraba-Djebari, F. Effect of Cytokine Antibodies in the Immunomodulation of Inflammatory Response and Metabolic Disorders Induced by Scorpion Venom. Int. Immunopharmacol. 2015, 27, 122–129. [Google Scholar] [CrossRef]
- Eliason, J.F. Pegylated Cytokines: Potential Application in Immunotherapy of Cancer. BioDrugs 2001, 15, 705–711. [Google Scholar] [CrossRef]
- Calvete, J.J.; Juárez, P.; Sanz, L. Snake Venomics. Strategy and Applications. J. Mass. Spectrom. 2007, 42, 1405–1414. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Skoog, B. Determination of Polyethylene Glycols 4000 and 6000 in Plasma Protein Preparations. Vox Sang 1979, 37, 345–349. [Google Scholar] [CrossRef]
- de Oliveira, I.S.; Pucca, M.B.; Wiezel, G.A.; Cardoso, I.A. Unraveling the Structure and Function of CdcPDE: A Novel Phosphodiesterase from Crotalus durissus collilineatus Snake Venom. Int. J. Biol. Macromol. 2021, 13, 180–192. [Google Scholar] [CrossRef]
- Saha, S.; Raghava, G.P.S. Prediction of Continuous B-Cell Epitopes in an Antigen Using Recurrent Neural Network. Proteins 2006, 65, 40–48. [Google Scholar] [CrossRef]
- Söding, J.; Biegert, A.; Lupas, A.N. The HHpred Interactive Server for Protein Homology Detection and Structure Prediction. Nucleic Acids Res. 2005, 33, W244–W248. [Google Scholar] [CrossRef] [Green Version]
- Webb, B.; Sali, A. Protein Structure Modeling with MODELLER. Methods Mol. Biol. 2021, 2199, 239–255. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology Modelling of Protein Structures and Complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laskowski, R.A.; Rullmannn, J.A.; MacArthur, M.W.; Kaptein, R.; Thornton, J.M. AQUA and PROCHECK-NMR: Programs for Checking the Quality of Protein Structures Solved by NMR. J. Biomol. NMR 1996, 8, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Grant, D.S.; Tashiro, K.-I.; Segui-Real, B.; Yamada, Y.; Martin, G.R.; Kleinman, H.K. Two different laminin domains mediate the differentiation of human endothelial cells into capillary-like structures in vitro. Cell 1989, 58, 933–943. [Google Scholar] [CrossRef]
- Yeh, C.H.; Peng, H.C.; Yang, R.S.; Huang, T.F. Rhodostomin, a Snake Venom Disintegrin, Inhibits Angiogenesis Elicited by Basic Fibroblast Growth Factor and Suppresses Tumor Growth by a Selective Alpha(v)Beta(3) Blockade of Endothelial Cells. Mol. Pharmacol. 2001, 59, 1333–1342. [Google Scholar] [CrossRef] [Green Version]
- Page, B.; Page, M.; Noel, C. A New Fluorometric Assay for Cytotoxicity Measurements In-Vitro. Int. J. Oncol. 1993, 3, 473–476. [Google Scholar] [CrossRef]
- Liang, C.-C.; Park, A.Y.; Guan, J.-L. In Vitro Scratch Assay: A Convenient and Inexpensive Method for Analysis of Cell Migration in Vitro. Nat. Protoc. 2007, 2, 329–333. [Google Scholar] [CrossRef] [Green Version]
- Marion, M.B. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
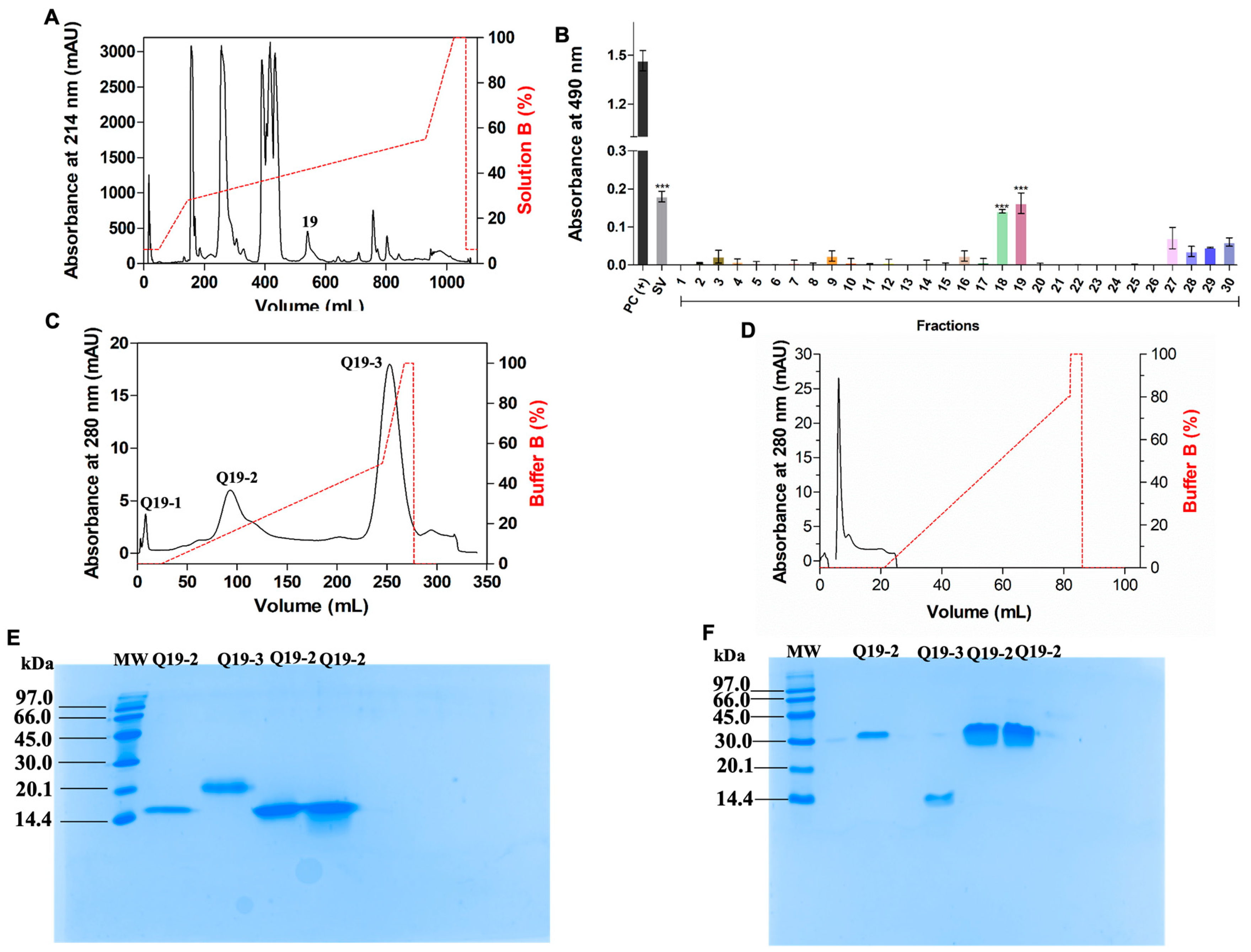
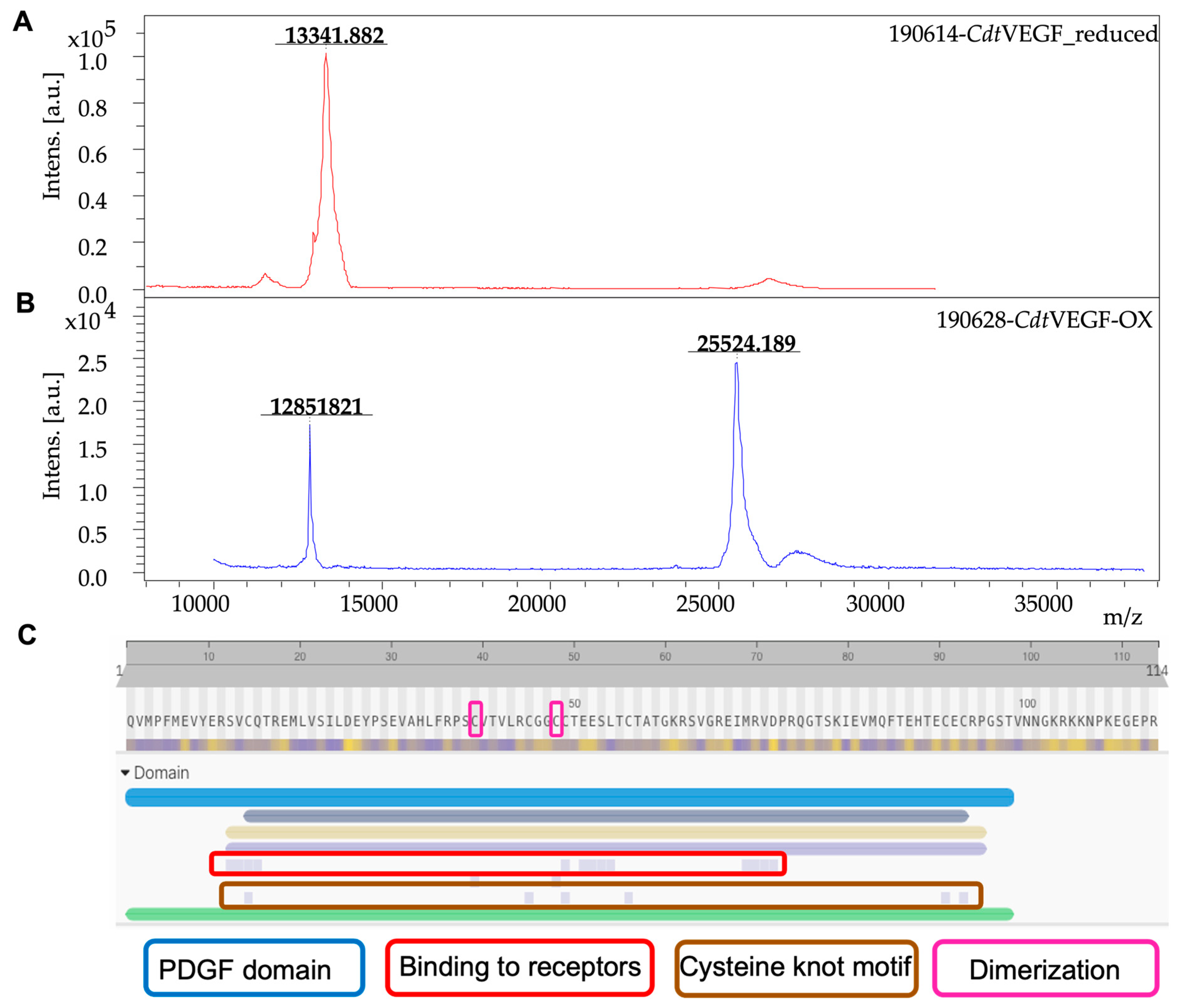
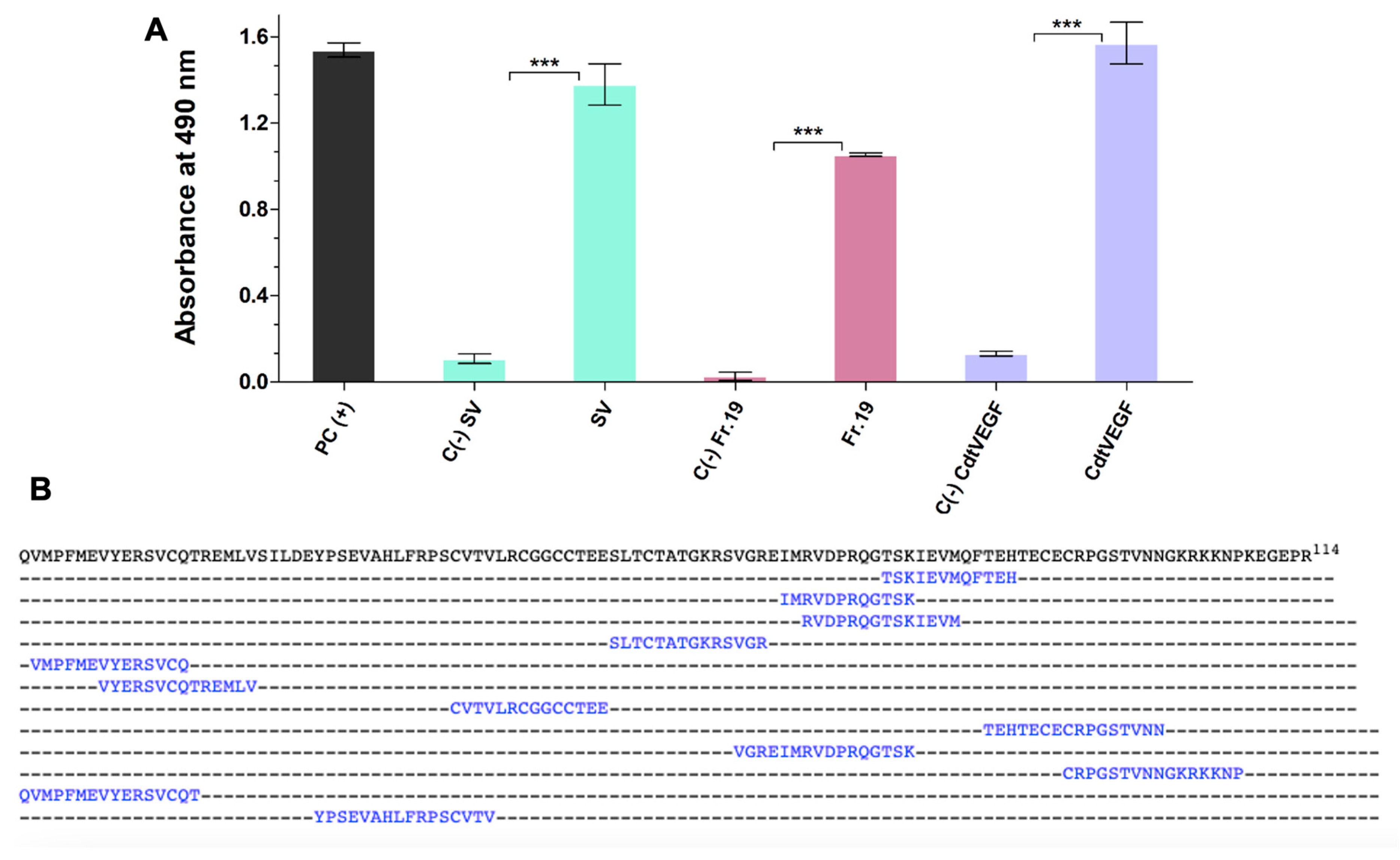
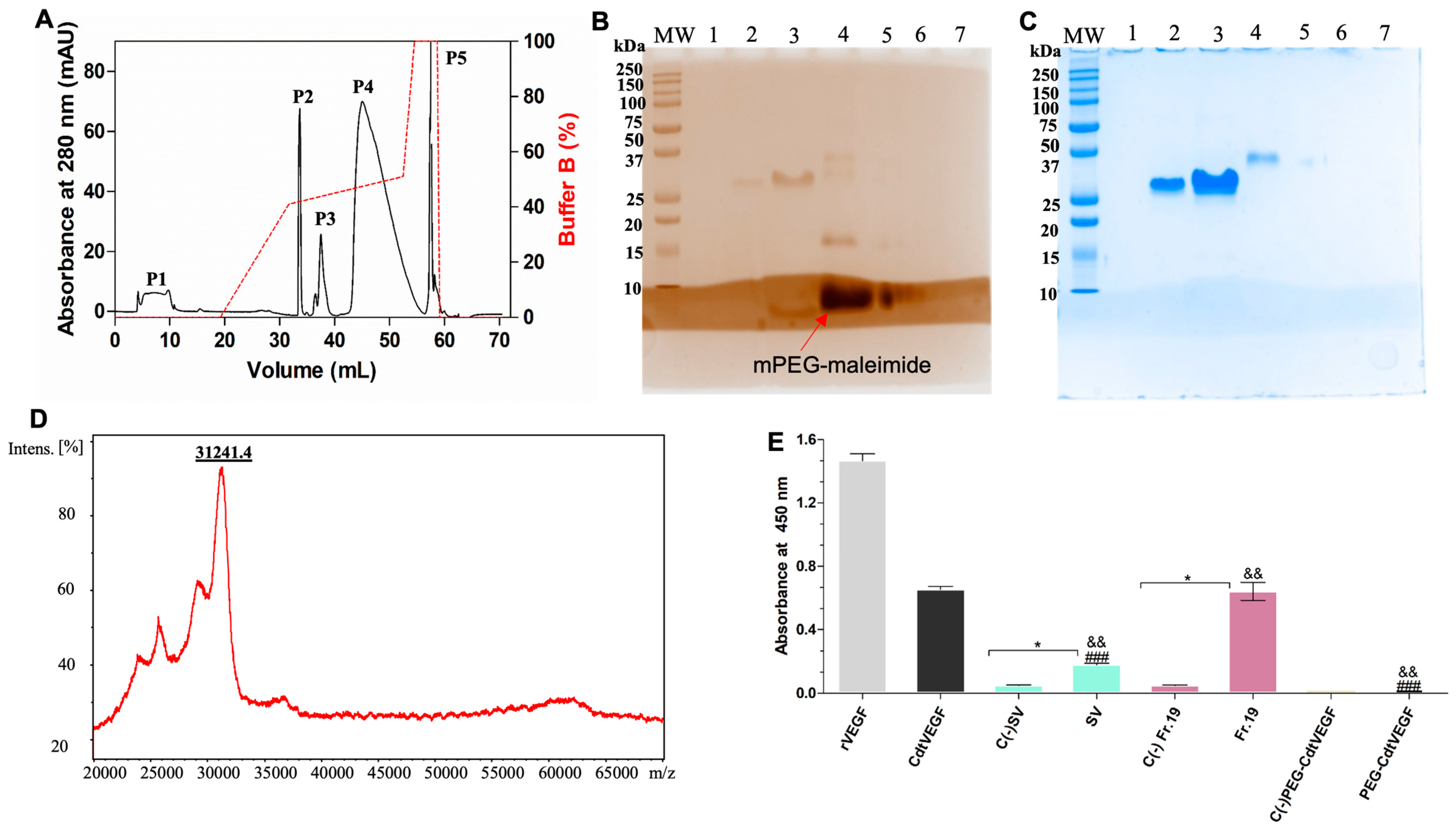
| Sample | Mass (mg) | Purification Steps | Recovery (%) |
|---|---|---|---|
| Crude venom | 50.0 | Supernatant | 100 |
| Fraction 19 | 3.1 | Reversed phase | 6.2 |
| CdtVEGF | 1.0 | Anion–ion exchange | 2.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferreira, I.; Oliveira, I.; Bordon, K.; Reis, M.; Wiezel, G.; Sanchez, C.; Santos, L.; Santos-Filho, N.; Pucca, M.; Antunes, L.; et al. Beyond Angiogenesis: The Multitasking Approach of the First PEGylated Vascular Endothelial Growth Factor (CdtVEGF) from Brazilian Rattlesnake Venom. Toxins 2023, 15, 483. https://doi.org/10.3390/toxins15080483
Ferreira I, Oliveira I, Bordon K, Reis M, Wiezel G, Sanchez C, Santos L, Santos-Filho N, Pucca M, Antunes L, et al. Beyond Angiogenesis: The Multitasking Approach of the First PEGylated Vascular Endothelial Growth Factor (CdtVEGF) from Brazilian Rattlesnake Venom. Toxins. 2023; 15(8):483. https://doi.org/10.3390/toxins15080483
Chicago/Turabian StyleFerreira, Isabela, Isadora Oliveira, Karla Bordon, Mouzarllem Reis, Gisele Wiezel, Caroline Sanchez, Luísa Santos, Norival Santos-Filho, Manuela Pucca, Lusânia Antunes, and et al. 2023. "Beyond Angiogenesis: The Multitasking Approach of the First PEGylated Vascular Endothelial Growth Factor (CdtVEGF) from Brazilian Rattlesnake Venom" Toxins 15, no. 8: 483. https://doi.org/10.3390/toxins15080483