Elateriospermum tapos Yogurt Supplement in Maternal Obese Dams during Pregnancy Modulates the Body Composition of F1 Generation

 ,
,  , , , ,
, , , ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection and Identification of E. tapos Seeds
2.2. Extraction of E. tapos Seeds
2.3. Formulation of E. tapos Yogurt
2.4. High-Fat Diet Preparation
2.5. Experimental Animals
2.6. Obesity Induction, Mating, Gestation, and Weaning
2.7. Plasma Biochemistry
2.8. Lipid Profile and Leptin Analysis
2.9. Gross Organ Weight and Histological Analysis
2.10. Quantitative Analysis of Fat Cells
2.11. Statistical Analysis
3. Results
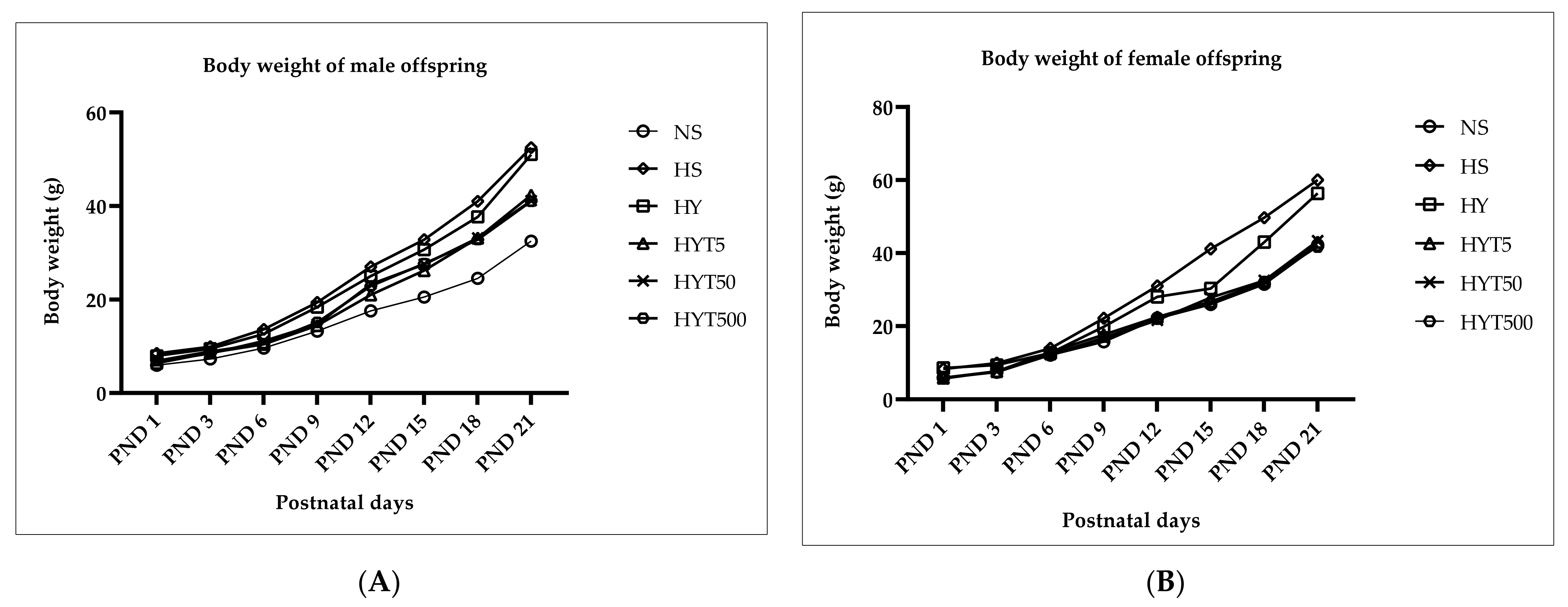
3.1. Changes in Body Weight in Male and Female Offspring from PND 1 to PND 21
3.2. Gross Organ Weight of Male and Female Offspring on PND 21
3.3. Liver Profile of Male and Female Offspring on PND 21
3.4. Renal Profiles of Male and Female Offspring on PND 21
3.5. Lipid Profiles of Male and Female Offspring on PND 21
3.6. Leptin Levels in Male and Female Offspring on PND 21
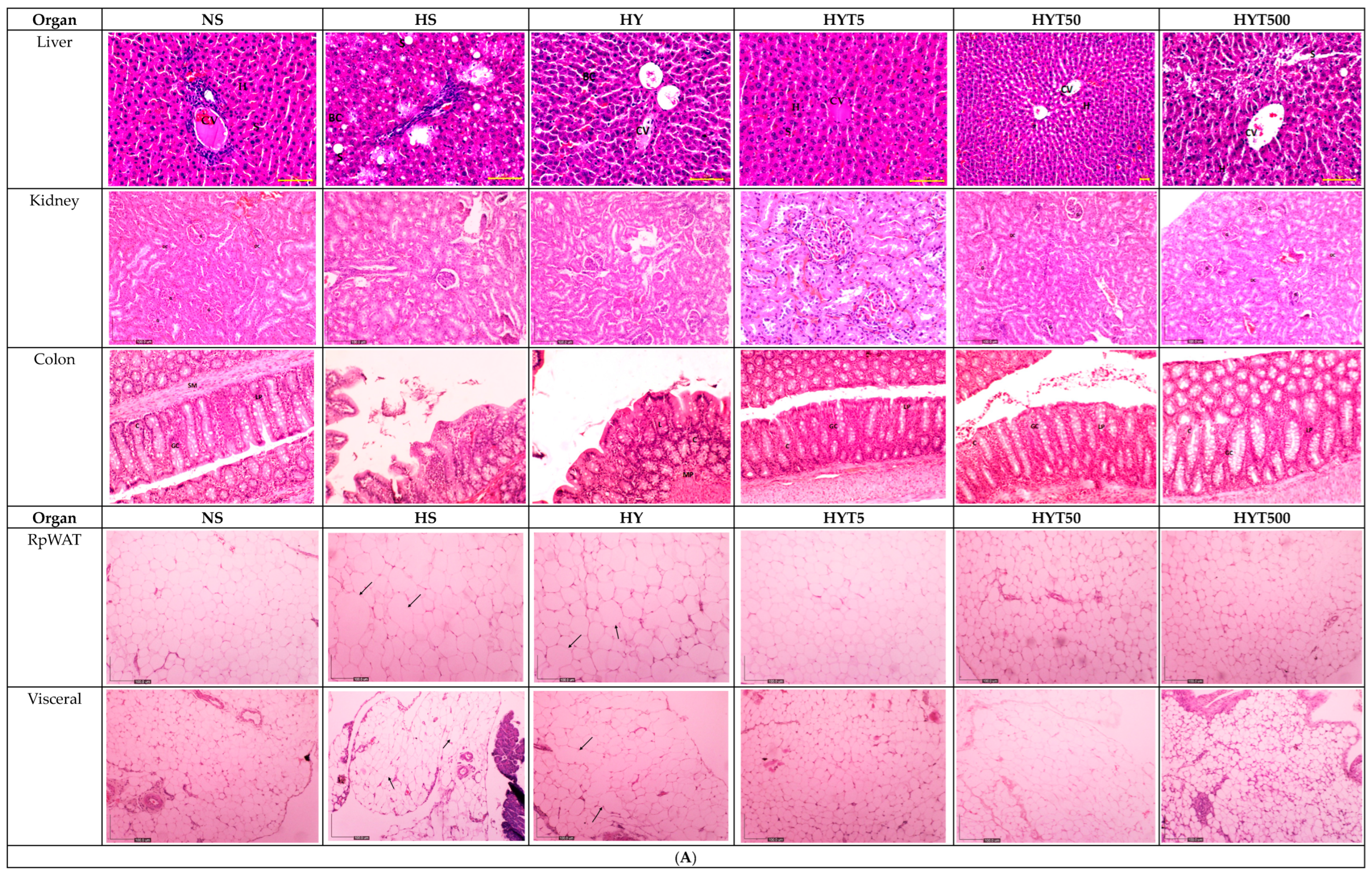
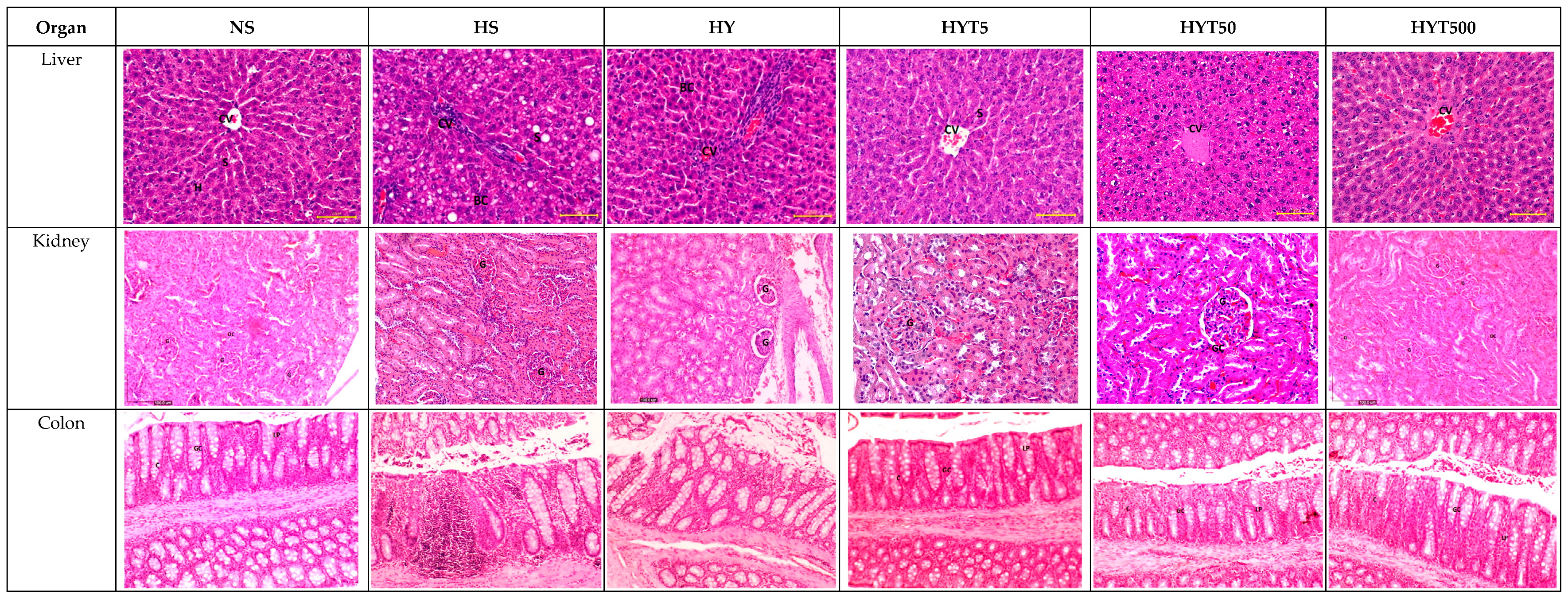
3.7. Histological Analysis of the Liver, Kidney, Colon, RpWAT, and Visceral Tissue of Male and Female Offspring on PND 21
3.8. Quantitative Analysis of Fat Hypertrophy of Male and Female Offspring on PND 21
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Denizli, M.; Capitano, M.L.; Kua, K.L. Maternal obesity and the impact of associated early-life inflammation on long-term health of offspring. Front. Cell. Infect. Microbiol. 2022, 12, 1348. [Google Scholar] [CrossRef] [PubMed]
- Forno, A.E.; Young, O.M.; Kumar, R.; Simhan, H.; Celedón, J.C. Maternal Obesity in Pregnancy, Gestational Weight Gain, and Risk of Childhood Asthma. Pediatrics 2014, 134, 535–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heslehurst, N.; Vieira, R.; Akhter, Z.; Bailey, H.; Slack, E.; Ngongalah, L.; Pemu, A.; Rankin, J. The association between maternal body mass index and child obesity: A systematic review and meta-analysis. PLoS Med. 2019, 16, e1002817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Xu, X.; Yan, Y. Estimated global overweight and obesity burden in pregnant women based on panel data model. PLoS ONE 2018, 13, e0202183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahari, H.; Abidin, A.Z.; Balan, S.S.; Perumal, K.V.; Rosli, N.S.; Lotafi, A.H.A.; Danabala, S.; Manimaran, M.; Shafie, N.H.; Abdullah, M.A.; et al. The effects of Elateriospermum tapos against obese maternal rat in mitigating obesity development among their adult female offspring. Pharmacogn. Mag. 2020, 16, 706–712. [Google Scholar] [CrossRef]
- Moschonis, G.; Siopis, G.; Anastasiou, C.; Iotova, V.; Stefanova, T.; Dimova, R.; Rurik, I.; Radó, A.S.; Cardon, G.; De Craemer, M.; et al. Prevalence of Childhood Obesity by Country, Family Socio-Demographics, and Parental Obesity in Europe: The Feel4Diabetes Study. Nutrients 2022, 14, 1830. [Google Scholar] [CrossRef]
- Wang, Y.W.; Yu, H.R.; Tiao, M.M.; Tain, Y.L.; Lin, I.C.; Sheen, J.M.; Lin, Y.J.; Chang, K.A.; Chen, C.C.; Tsai, C.C.; et al. Maternal Obesity Related to High Fat Diet Induces Placenta Remodeling and Gut Microbiome Shaping That Are Responsible for Fetal Liver Lipid Dysmetabolism. Front. Nutr. 2021, 8, 1114. [Google Scholar] [CrossRef]
- Gillman, M.W. Interrupting Intergenerational Cycles of Maternal Obesity. Nestle Nutr. Inst. Workshop Ser. 2016, 85, 59–69. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.W.; Aubert, A.M.; Shivappa, N.; Bernard, J.Y.; Mensink-Bout, S.M.; Geraghty, A.A.; Mehegan, J.; Suderman, M.; Polanska, K.; Hanke, W.; et al. Maternal dietary quality, inflammatory potential and childhood adiposity: An individual participant data pooled analysis of seven European cohorts in the ALPHABET consortium. BMC Med. 2021, 19, 33. [Google Scholar] [CrossRef]
- Gamboa-gómez, C.I.; Rocha-guzmán, N.E.; Gallegos-infante, J.A.; Moreno-jiménez, M.R.; Vázquez-cabral, B.D.; González-laredo, R.F. Palnts with potential use on obesity and its complications. EXCLI J. 2015, 14, 809–831. [Google Scholar] [CrossRef]
- Perumal, K.V.; Ja’afar, N.L.; Taib, C.N.M.; Shafie, N.H.; Bahari, H. Antiobesity activity of Elateriospermum tapos shell extract in obesity-induced sprague dawley rats. Molecules 2021, 26, 321. [Google Scholar] [CrossRef] [PubMed]
- Balan, S.S.; Abidin, A.Z.; Perumal, K.V.; Shafie, N.H.; Abdullah, M.A.; Jasni, A.S.; Kwaifa, I.K.; Bahari, H. Transgenerational evaluation of Elateriospermum tapos extracts on the male offspring of obesity-induced sprague dawley rats. Sains Malays. 2021, 50, 3045–3057. [Google Scholar] [CrossRef]
- Nor-Liyana, J.; Siroshini, K.T.; Nurul-Syahirah, M.B.; Chang, W.L.; Nurul-Husna, S.; Daryl, J.A.; Khairul-Kamilah, A.K.; Hasnah, B. Phytochemical analysis of Elateriospermum tapos and its inhibitory effects on alpha-amylase, alpha-glucosidase and pancreatic lipase. J. Trop. For. Sci. 2019, 31, 240–248. [Google Scholar] [CrossRef]
- Sayon-Orea, C.; Martínez-González, M.A.; Ruiz-Canela, M.; Bes-Rastrollo, M. Associations between Yogurt Consumption and Weight Gain and Risk of Obesity and Metabolic Syndrome: A Systematic Review. Adv. Nutr. Int. Rev. J. 2017, 8, 146S–154S. [Google Scholar] [CrossRef] [Green Version]
- Naomi, R.; Rusli, R.N.M.; Balan, S.S.; Othman, F.; Jasni, A.S.; Jumidil, S.H.; Bahari, H.; Yazid, M.D. E. tapos Yoghurt—A View from Nutritional Composition and Toxicological Evaluation. Foods 2022, 11, 1903. [Google Scholar] [CrossRef]
- Sita, T.; Thanaset, S.; Prasan, S.; Auamporn, R. Antioxidant and antiproliferative activities of ethanolic extracts of Elateriospermum tapos Blume (Euphorbiaceae). J. Med. Plants Res. 2018, 12, 474–482. [Google Scholar] [CrossRef] [Green Version]
- Siccama, J.W.; Pegiou, E.; Zhang, L.; Mumm, R.; Hall, R.D.; Boom, R.M.; Schutyser, M.A.I. Maltodextrin improves physical properties and volatile compound retention of spray-dried asparagus concentrate. LWT-Food Sci. Technol. 2021, 142, 111058. [Google Scholar] [CrossRef]
- Aril-Dela Cruz, J.V.; Bungihan, M.E.; Dela Cruz, T.E.E.; Sagum, R.S. Canarium ovatum engl. (Pili) exocarp crude extract as functional food colorant incorporated in yogurt developed product. Food Res. 2017, 2, 89–98. [Google Scholar] [CrossRef]
- Balan, S.S.; Abidin, A.Z.; Perumal, K.V.; Shafie, N.H.; Abdullah, M.A.; Jasni, A.S.; Kadir, K.K.A.; Bahari, H. Elacteriospermum tapos Ameliorates Maternal Obesity Effect on Serum Leptin Changes in Male Offspring. Proceedings 2020, 61, 3. [Google Scholar] [CrossRef]
- Ypsilantis, P.; Somalou, P.; Panidou, E.; Simopoulos, C. Laparoscopic early pregnancy diagnosis in the laboratory rat. Lab. Anim. 2018, 52, 265–270. [Google Scholar] [CrossRef]
- Kadir, N.A.A.; Rahmat, A.; Jaafar, H.Z.E. Protective Effects of Tamarillo (Cyphomandra betacea) Extract Against High Fat Diet Induced Obesity In Sprague Dawley Rats. J. Obes. 2015, 2015, 846041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aberare, O.L.; Okuonghae, P.; Mukoro, N.; Dirisu, J.O.; Osazuwa, F.; Odigie, E.; Omoregie, R. Triglycerides, total cholesterol, high density lipoprotein cholesterol and low density lipoprotein cholesterol in rats exposed to premium motor spirit fumes. N. Am. J. Med. Sci. 2011, 3, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Labban, R.S.M.; Alfawaz, H.A.; Amina, M.; Bhat, R.S.; Hassan, W.M.; El-Ansary, A. Synergism between Extracts of Garcinia mangostana Pericarp and Curcuma in Ameliorating Altered Brain Neurotransmitters, Systemic Inflammation, and Leptin Levels in High-Fat Diet-Induced Obesity in Male Wistar Albino Rats. Nutrients 2022, 14, 4630. [Google Scholar] [CrossRef] [PubMed]
- Paul, M. Histological Slide Preparation of Fish Tissues (Paraffin Method); Asutosh College: Kolkata, India, 2017. [Google Scholar] [CrossRef]
- Lundgren, M.; Svensson, M.; Lindmark, S.; Renström, F.; Ruge, T.; Eriksson, J.W. Fat cell enlargement is an independent marker of insulin resistance and “hyperleptinaemia”. Diabetologia 2006, 50, 625–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doulla, M.; McIntyre, A.D.; Hegele, R.A.; Callego, P.H. A novel MC4R mutation associated with childhood-onset obesity: A case report. Paediatr. Child Health 2014, 19, 515–518. [Google Scholar] [CrossRef] [Green Version]
- Kansra, A.R.; Lakkunarajah, S.; Jay, M.S. Childhood and Adolescent Obesity: A Review. Front. Pediatr. 2020, 8, 581461. [Google Scholar] [CrossRef]
- Durrani, A.M.; Rani, A. Effect of maternal dietary intake on the weight of the newborn in Aligarh city, India. Niger. Med. J. 2011, 52, 177–181. [Google Scholar] [CrossRef] [Green Version]
- Shah, B.; Tombeau Cost, K.; Fuller, A.; Birken, C.S.; Anderson, L.N. Sex and gender differences in childhood obesity: Contributing to the research agenda. BMJ Nutr. Prev. Health 2020, 3, 387–390. [Google Scholar] [CrossRef]
- Quyen, P.N.; Nga, H.T.; Chaffee, B.; Ngu, T.; King, J.C. Effect of maternal prenatal food supplementation, gestational weight gain, and breast-feeding on infant growth during the first 24 months of life in rural Vietnam. PLoS ONE 2020, 15, e0233671. [Google Scholar] [CrossRef]
- Sachdeva, P.; Patel, B.G.; Patel, B.K. Drug Use in Pregnancy: A Point to Ponder! Indian J. Pharm. Sci. 2009, 71, 1. [Google Scholar] [CrossRef] [Green Version]
- The Metabolomics Innovation Centre Human Metabolome Database: Showing Metabocard for 5’-Methoxybilobetin (HMDB0038101). Available online: https://hmdb.ca/metabolites/HMDB0038101 (accessed on 27 December 2022).
- Hung, T.C.; Kuo, C.C.; Chen, C.Y.C. In Silico investigation of cytochrome P450 2C9 in relation to aging using traditional Chinese medicine. Evid. Based Complement. Altern. Med. 2014, 2014, 404505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noor-E-Tabassum; Das, R.; Lami, M.S.; Chakraborty, A.J.; Mitra, S.; Tallei, T.E.; Idroes, R.; Mohamed, A.A.R.; Hossain, M.J.; Dhama, K.; et al. Ginkgo biloba: A Treasure of Functional Phytochemicals with Multimedicinal Applications. Evid. Based Complement. Altern. Med. 2022, 2022, 8288818. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Fang, Q.; Zhu, F.; Huang, D.; Yang, C. Structure and Function of Pancreatic Lipase-Related Protein 2 and Its Relationship With Pathological States. Front. Genet. 2021, 12, 693538. [Google Scholar] [CrossRef] [PubMed]
- Aabideen, Z.U.; Mumtaz, M.W.; Akhtar, M.T.; Mukhtar, H.; Raza, S.A.; Touqeer, T.; Saari, N. Anti-Obesity Attributes; UHPLC-QTOF-MS/MS-Based Metabolite Profiling and Molecular Docking Insights of Taraxacum officinale. Molecules 2020, 25, 4935. [Google Scholar] [CrossRef] [PubMed]
- Rospond, B.; Krakowska, A.; Krośniak, M.; Muszyńska, B.; Opoka, W. The influence of high-fat and high-sucrose feeding regimes on organ weight, body weight, and serum concentration of bioelements in rats. J. Trace Elem. Med. Biol. 2022, 73, 127020. [Google Scholar] [CrossRef]
- Gao, H.; Mejhert, N.; Fretz, J.A.; Arner, E.; Lorente-Cebrián, S.; Ehrlund, A.; Dahlman-Wright, K.; Gong, X.; Strömblad, S.; Douagi, I.; et al. Early B cell factor 1 Regulates adipocyte morphology and lipolysis in white adipose tissue. Cell Metab. 2014, 19, 981–992. [Google Scholar] [CrossRef] [Green Version]
- Gulhane, M.; Murray, L.; Lourie, R.; Tong, H.; Sheng, Y.H.; Wang, R.; Kang, A.; Schreiber, V.; Wong, K.Y.; Magor, G.; et al. High Fat Diets Induce Colonic Epithelial Cell Stress and Inflammation that is Reversed by IL-22. Sci. Rep. 2016, 6, 28990. [Google Scholar] [CrossRef] [Green Version]
- Holt, P.R.; Atillasoy, E.O.; Gilman, J.; Guss, J.; Moss, S.F.; Newmark, H.; Fan, K.; Yang, K.; Lipkin, M. Modulation of abnormal colonic epithelial cell proliferation and differentiation by low-fat dairy foods: A randomized controlled trial. Jama 1998, 280, 1074–1079. [Google Scholar] [CrossRef] [Green Version]
- Enriquez, J.R.; McCauley, H.A.; Zhang, K.X.; Sanchez, J.G.; Kalin, G.T.; Lang, R.A.; Wells, J.M. High fat diet initiates rapid adaptation of the intestine. bioRxiv 2022. [Google Scholar] [CrossRef]
- Soyocak, A.; Kurt, H.; Cosan, D.T.; Saydam, F.; Calis, I.U.; Kolac, U.K.; Koroglu, Z.O.; Degirmenci, I.; Mutlu, F.S.; Gunes, H.V. Tannic acid exhibits anti-inflammatory effects on formalin-induced paw edema model of inflammation in rats. Hum. Exp. Toxicol. 2019, 38, 1296–1301. [Google Scholar] [CrossRef]
- Tong, Z.; He, W.; Fan, X.; Guo, A. Biological Function of Plant Tannin and Its Application in Animal Health. Front. Vet. Sci. 2022, 8, 1597. [Google Scholar] [CrossRef] [PubMed]
- Nobushi, Y.; Wada, T.; Koike, Y.; Kaneko, H.; Shimba, S.; Uchiyama, T.; Kishikawa, Y. Inhibitory Effects of Hydrolysable Tannins on Lipid Accumulation in 3T3-L1 Cells. Biol. Pharm. Bull. 2022, 45, 1458–1465. [Google Scholar] [CrossRef]
- Yugarani, T.; Tan, B.K.H.; Teh, M.; Das, N.P. Effects of polyphenolic natural products on the lipid profiles of rats fed high fat diets. Lipids 1992, 27, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Manzoor, F.; Nisa, M.U.; Hussain, H.A.; Khan, M.K.; Ahmad, R.S.; Ahmad, N.; Imran, M.; Umbreen, H. Effect of hydrolysable tannin on nutrient intake obesity and other associated metabolic risk factors in polycystic rats. Transl. Med. Commun. 2021, 6, 10. [Google Scholar] [CrossRef]
- Zhukova, N.V.; Novgorodtseva, T.P.; Denisenko, Y.K. Effect of the prolonged high-fat diet on the fatty acid metabolism in rat blood and liver. Lipids Health Dis. 2014, 13, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tebib, K.; Besancon, P.; Rouanet, J.M. Dietary grape seed tannins affect lipoproteins, lipoprotein lipases and tissue lipids in rats fed hypercholesterolemic diets. J. Nutr. 1994, 124, 2451–2457. [Google Scholar] [CrossRef]
- Gato, N.; Kadowaki, A.; Hashimoto, N.; Yokoyama, S.I.; Matsumoto, K. Persimmon fruit tannin-rich fiber reduces cholesterol levels in humans. Ann. Nutr. Metab. 2013, 62, 1–6. [Google Scholar] [CrossRef]
- Soliman, A.; Yasin, M.; Kassem, A. Leptin in pediatrics: A hormone from adipocyte that wheels several functions in children. Indian J. Endocrinol. Metab. 2012, 16, S577–S587. [Google Scholar] [CrossRef]
- Harrell, J.S.; Bomar, P.; McMurray, R.; Bradley, C.; Deng, S. Leptin and Obesity in Mother-Child Pairs. Biol. Res. Nurs. 2001, 3, 55–64. [Google Scholar] [CrossRef]
- Ippagunta, S.; Hadenfeldt, T.J.; Miner, J.L.; Hargrave-Barnes, K.M. Dietary conjugated linoleic acid induces lipolysis in adipose tissue of coconut oil-fed mice but not soy oil-fed mice. Lipids 2011, 46, 821–830. [Google Scholar] [CrossRef]
- Hall, J.E. The kidney, hypertension, and obesity. Hypertension 2003, 41, 625–633. [Google Scholar] [CrossRef] [Green Version]
- Sun, K.; Su, T.; Li, M.; Xu, B.; Xu, M.; Lu, J.; Liu, J.; Bi, Y.; Ning, G. Serum potassium level is associated with metabolic syndrome: A population-based study. Clin. Nutr. 2013, 33, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Noireaud, J.; Bright, C.M.; Ellis, D. Effects of saponin on contractile force and intracellular ion activities of cardiac tissues. J. Mol. Cell. Cardiol. 1989, 21, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.N.; Pergulwar, A.; Siddiqui, A.M.; Shinde, A.R. Estimation of Serum Urea, Creatinine and Uric Acid in Obese Subjects. Int. J. Innov. Res. Med. Sci. 2018, 2, 1201–1203. [Google Scholar] [CrossRef]
- Kim, E.J.; Oh, H.A.; Choi, H.J.; Park, J.H.; Kim, D.H.; Kim, N.J. Heat-processed ginseng saponin ameliorates the adenine-induced renal failure in rats. J. Ginseng Res. 2013, 37, 87–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basolo, A.; Ando, T.; Chang, D.C.; Hollstein, T.; Krakoff, J.; Piaggi, P.; Votruba, S. Reduced Albumin Concentration Predicts Weight Gain and Higher Ad Libitum Energy Intake in Humans. Front. Endocrinol. 2021, 12, 642568. [Google Scholar] [CrossRef] [PubMed]
- Ali, N.; Sumon, A.H.; Fariha, K.A.; Asaduzzaman, M.; Kathak, R.R.; Molla, N.H.; Mou, A.D.; Barman, Z.; Hasan, M.; Miah, R.; et al. Assessment of the relationship of serum liver enzymes activity with general and abdominal obesity in an urban Bangladeshi population. Sci. Rep. 2021, 11, 6640. [Google Scholar] [CrossRef] [PubMed]
- Žiberna, L.; Jenko-Pražnikar, Z.; Petelin, A. Serum bilirubin levels in overweight and obese individuals: The importance of anti-inflammatory and antioxidant responses. Antioxidants 2021, 10, 1352. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gross Organ Weight of Male Offspring on PND 21 | ||||||||
|---|---|---|---|---|---|---|---|---|
| BAT (g) | Liver (g) | Kidney (g) | Colon (g) | Colon Length (cm) | RpWAT (g) | Visceral (g) | Gonadal (g) | |
| NS | 4.97 ± 1.27 a | 0.92 ± 0.09 a | 0.75 ± 0.17 a | 11.50 ± 1.50 a | 0.25 ± 0.07 a | 0.30 ± 0.08 a | 0.29 ± 0.10 a | |
| HS | 0.46 ± 0.02 b | 9.94 ± 1.15 b | 2.34 ± 0.13 b | 1.52 ± 0.09 b | 18.17 ± 0.83 b | 2.53 ± 0.34 b | 2.84 ± 0.37 b | 2.51 ± 0.42 b |
| HY | 0.48 ± 0.02 b | 7.66 ± 0.45 ab | 2.22 ± 0.13 b | 1.35 ± 0.05 bc | 15.80 ± 0.20 bc | 1.48 ± 0.01 c | 1.47 ± 0.04 c | 1.49 ± 0.15 c |
| HYT5 | 0.30 ± 0.04 a | 5.72 ± 0.16 a | 1.09 ± 0.12 a | 0.82 ± 0.17 ac | 11.67 ± 0.67 ac | 0.58 ± 0.13 a | 0.73 ± 0.11 ac | 0.66 ± 0.15 ac |
| HYT50 | 0.27 ± 0.02 a | 6.59 ± 0.18 ab | 1.22 ± 0.06 a | 0.99 ± 0.11 abc | 12.50 ± 0.50 ac | 0.76 ± 0.03 ac | 0.78 ± 0.02 ac | 0.81 ± 0.10 ac |
| HYT500 | 0.31 ± 0.01 a | 6.19 ± 0.18 ab | 1.34 ± 0.03 a | 0.88 ± 0.08 ac | 11.83 ± 1.09 ac | 0.51 ± 0.09 a | 0.79 ± 0.09 ac | 0.82 ± 0.08 ac |
| Gross Organ Weight of Female Offspring on PND 21 | ||||||||
| BAT (g) | Liver (g) | Kidney (g) | Colon (g) | Colon Length (cm) | RpWAT (g) | Visceral (g) | Gonadal (g) | |
| NS | 0.18 ± 0.04 a | 3.35 ± 0.53 a | 0.83 ± 0.15 a | 0.96 ± 0.10 | 11.93 ± 0.52 a | 0.14 ± 0.06 a | 0.49 ± 0.11 a | 0.29 ± 0.05 a |
| HS | 0.40 ± 0.03 b | 7.48 ± 0.65 b | 1.43 ± 0.14 b | 1.08 ± 0.09 | 17.67 ± 0.44 b | 1.84 ± 0.13 b | 2.03 ± 0.51 b | 3.30 ± 1.07 b |
| HY | 0.39 ± 0.02 b | 6.12 ± 0.44 bc | 1.32 ± 0.20 bc | 1.00 ± 0.08 | 14.67 ± 0.33 c | 1.38 ± 0.17 b | 1.25 ± 0.21 ab | 2.41 ± 0.79 b |
| HYT5 | 0.20 ± 0.03 a | 3.69 ± 0.53 a | 0.87 ± 0.18 a | 0.99 ± 0.10 | 11.87 ± 0.13 a | 0.54 ± 0.09 a | 0.73 ± 0.09 a | 0.49 ± 0.11 a |
| HYT50 | 0.21 ± 0.02 a | 3.69 ± 0.17 a | 0.93 ± 0.11 ac | 1.15 ± 0.08 | 11.67 ± 0.33 a | 0.44 ± 0.02 a | 0.46 ± 0.06 a | 0.39 ± 0.08 a |
| HYT500 | 0.22 ± 0.01 a | 4.27 ± 0.55 ac | 0.89 ± 0.15 a | 1.19 ± 0.14 | 11.83 ± 0.44 a | 0.49 ± 0.02 a | 0.72 ± 0.24 a | 0.71 ± 0.42 a |
| Liver Profile of Male Offspring on PND 21 | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Total Protein (g/L) | Albumin (g/L) | Globulin (g/L) | Albumin–Globulin Ratio | ALP (U/L) | AST (U/L) | ALT (U/L) | Gamma-Glutamyl Transferase (U/L) | Total Bilirubin (µmol/L) | |
| NS | 56.00 ± 0.40 | 41.33 ± 1.67 a | 18.67 ± 0.33 a | 1.77 ± 0.05 a | 301.67 ± 9.74 a | 152.67 ± 11.67 a | 55.33 ± 5.49 a | 1.67 ± 0.67 a | 2.33 ± 0.33 a |
| HS | 56.50 ± 0.65 | 35.33 ± 0.33 b | 19.33 ± 0.88 ab | 2.06 ± 0.03 b | 896.33 ± 29.46 b | 290.00 ± 7.57 b | 102.67 ± 7.42 b | 13.00 ± 0.00 b | 1.00 ± 0.00 b |
| HY | 57.25 ± 1.11 | 39.33 ± 0.33 ab | 20.00 ± 0.58 ab | 1.88 ± 0.01 a | 676.00 ± 35.47 c | 194.67 ± 6.84 a | 85.33 ± 0.88 bc | 9.67 ± 3.33 bc | 1.00 ± 0.00 b |
| HYT5 | 57.25 ± 0.25 | 41.00 ± 0.58 a | 19.33 ± 0.33 ab | 1.79 ± 0.02 a | 389.00 ± 38.55 ad | 161.67 ± 25.83 a | 59.67 ± 1.45 ad | 1.67 ± 0.33 a | 2.33 ± 0.33 a |
| HYT50 | 57.75 ± 0.48 | 38.00 ± 0.58 ab | 20.67 ± 0.33 ab | 1.85 ± 0.05 a | 484.67 ± 53.72 d | 178.67 ± 4.67 a | 96.33 ± 1.76 b | 4.00 ± 1.53 ac | 2.33 ± 0.33 a |
| HYT500 | 56.50 ± 0.29 | 40.67 ± 1.45 a | 21.67 ± 0.88 b | 1.86 ± 0.02 a | 528.67 ± 12.45 cd | 174.67 ± 15.07 a | 74.67 ± 1.86 cd | 5.67 ± 1.33 abc | 2.33 ± 0.33 a |
| Liver Profile of Female Offspring on PND 21 | |||||||||
| Total Protein (g/L) | Albumin (g/L) | Globulin (g/L) | Albumin–Globulin Ratio | ALT (U/L) | AST (U/L) | ALP (U/L) | Gamma-Glutamyl Transferase (U/L) | Total Bilirubin (µmol/L) | |
| NS | 59.17 ± 3.83 | 41.67 ± 1.45 a | 17.00 ± 2.08 a | 1.69 ± 0.04 a | 246.00 ± 46.65 a | 142.00 ± 11.93 a | 48.00 ± 5.57 a | 1.33 ± 0.33 a | 3.00 ± 0.00 a |
| HS | 56.29 ± 1.63 | 35.33 ± 0.67 bc | 23.67 ± 1.86 b | 2.04 ± 0.04 bc | 633.33 ± 99.09 b | 210.00 ± 23.64 b | 98.00 ± 2.65 b | 13.00 ± 0.00 b | 1.33 ± 0.33 b |
| HY | 60.67 ± 1.41 | 36.67 ± 0.88 c | 23.00 ± 0.58 b | 1.92 ± 0.04 c | 488.00 ± 22.52 ab | 191.33 ± 32.42 b | 87.00 ± 2.08 b | 6.67 ± 3.28 ab | 1.33 ± 0.33 b |
| HYT5 | 61.00 ± 2.02 | 41.00 ± 0.58 a | 19.33 ± 0.33 ab | 1.68 ± 0.03 a | 271.00 ± 48.00 a | 150.67 ± 9.02 a | 47.33 ± 2.19 a | 1.33 ± 0.33 a | 2.67 ± 0.33 ab |
| HYT50 | 59.33 ± 1.23 | 39.33 ± 0.67 ac | 19.67 ± 0.33 ab | 1.80 ± 0.07 ac | 254.00 ± 30.17 a | 148.67 ± 13.09 a | 60.33 ± 3.18 a | 3.33 ± 1.86 a | 2.33 ± 0.33 ab |
| HYT500 | 59.29 ± 2.07 | 38.33 ± 0.33 abc | 18.67 ± 0.33 ab | 1.75 ± 0.05 ac | 305.00 ± 35.03 a | 134.33 ± 2.40 a | 52.33 ± 6.23 a | 3.33 ± 1.86 a | 2.33 ± 0.33 ab |
| Renal Profiles of Male Offspring on PND 21 | |||||
|---|---|---|---|---|---|
| Na (mmol/L) | K (mmol/L) | Cl− (mmol/L) | Urea (mmol/L) | Creatinine (mmol/L) | |
| NS | 147.33 ± 0.88 a | 8.60 ± 0.44 ab | 101.67 ± 2.09 | 6.47 ± 0.15 ab | 30.33 ± 0.88 a |
| HS | 141.67 ± 0.33 b | 10.03 ± 0.03 c | 102.86 ± 2.11 | 7.53 ± 0.29 a | 40.00 ± 1.53 b |
| HY | 143.00 ± 0.58 bc | 9.73 ± 0.22 ac | 100.67 ± 0.76 | 5.97 ± 0.03 bc | 39.00 ± 1.15 bc |
| HYT5 | 145.67 ± 0.33 ad | 8.17 ± 0.55 b | 100.38 ± 0.78 | 6.23 ± 0.07 ab | 30.67 ± 0.88 a |
| HYT50 | 143.67 ± 0.33 cd | 8.90 ± 0.21 bc | 101.33 ± 1.71 | 7.27 ± 0.58 ab | 34.33 ± 1.45 ac |
| HYT500 | 145.67 ± 0.33 ad | 8.48 ± 0.13 ab | 103.50 ± 1.69 | 6.07 ± 0.32 bc | 32.67 ± 1.20 a |
| Renal Profiles of Female Offspring on PND 21 | |||||
| Na (mmol/L) | K (mmol/L) | Cl−(mmol/L) | Urea (mmol/L) | Creatinine (mmol/L) | |
| NS | 143.83 ± 1.67 | 8.27 ± 0.27 ab | 102.00 ± 1.55 | 5.57 ± 0.13 a | 27.67 ± 4.67 a |
| HS | 144.14 ± 0.83 | 10.60 ± 0.40 c | 103.40 ± 2.18 | 7.40 ± 0.50 b | 39.67 ± 1.67 b |
| HY | 144.00 ± 0.47 | 9.27 ± 0.27 abc | 101.67 ± 0.67 | 5.93 ± 0.24 a | 39.33 ± 0.33 b |
| HYT5 | 144.00 ± 0.68 | 8.03 ± 0.20 ab | 102.25 ± 0.25 | 5.70 ± 0.44 a | 28.33 ± 2.03 a |
| HYT50 | 142.83 ± 1.25 | 9.03 ± 0.48 ab | 106.00 ± 1.87 | 6.43 ± 0.09 ab | 30.33 ± 2.85 a |
| HYT500 | 144.29 ± 0.79 | 9.53 ± 0.12 ac | 104.60 ± 1.03 | 5.83 ± 0.03 a | 28.67 ± 3.18 a |
| Lipid Profiles of Male Offspring on PND 21 | |||||
|---|---|---|---|---|---|
| Total Cholesterol (mmol/L) | HDL (mmol/L) | Non-HDL (mmol/L) | LDL (mmol/L) | Triglyceride (mmol/L) | |
| NS | 2.13 ± 0.03 a | 0.87 ± 0.03 a | 0.97 ± 0.03 a | 0.73 ± 0.03 a | 0.73 ± 0.06 a |
| HS | 2.53 ± 0.03 bc | 0.60 ± 0.00 b | 1.73 ± 0.03 b | 1.30 ± 0.00 b | 1.23 ± 0.25 b |
| HY | 2.60 ± 0.06 b | 0.53 ± 0.03 b | 1.53 ± 0.13 b | 1.17 ± 0.07 b | 1.15 ± 0.26 a |
| HYT5 | 2.27 ± 0.03 a | 0.77 ± 0.03 a | 1.57 ± 0.07 b | 0.90 ± 0.15 a | 0.90 ± 0.10 a |
| HYT50 | 2.27 ± 0.13 a | 0.83 ± 0.03 a | 1.63 ± 0.03 b | 1.07 ± 0.09 b | 1.00 ± 0.26 a |
| HYT500 | 2.30 ± 0.00 ac | 0.77 ± 0.03 a | 1.70 ± 0.06 b | 1.13 ± 0.07 b | 0.80 ± 0.10 a |
| Lipid Profiles of Female Offspring on PND 21 | |||||
| Total Cholesterol (mmol/L) | HDL (mmol/L) | Non-HDL (mmol/L) | LDL (mmol/L) | Triglyceride (mmol/L) | |
| NS | 2.20 ± 0.12 a | 0.97 ± 0.03 a | 1.60 ± 0.06 a | 0.63 ± 0.09 a | 0.70 ± 0.10 a |
| HS | 3.00 ± 0.12 bc | 0.70 ± 0.00 b | 2.10 ± 0.06 bc | 1.70 ± 0.06 b | 2.13 ± 0.07 bc |
| HY | 2.77 ± 0.12 bc | 0.90 ± 0.06 a | 2.00 ± 0.10 c | 1.43 ± 0.03 b | 1.53 ± 0.26 c |
| HYT5 | 2.30 ± 0.06 ac | 0.90 ± 0.06 a | 1.57 ± 0.09 a | 1.00 ± 0.06 c | 0.97 ± 0.03 ac |
| HYT50 | 2.47 ± 0.07 ac | 1.00 ± 0.10 a | 1.67 ± 0.07 ac | 0.90 ± 0.12 ac | 1.10 ± 0.15 ac |
| HYT500 | 2.17 ± 0.12 a | 0.90 ± 0.06 a | 1.57 ± 0.09 a | 1.07 ± 0.03 c | 0.87 ± 0.09 a |
| Quantitative Fat Analysis of Male Offspring on PND 21 | ||
|---|---|---|
| Visceral (µm) | RpWAT (µm) | |
| NS | 16.52 ± 1.34 a | 16.96 ± 0.98 a |
| HS | 62.37 ± 5.48 b | 63.58 ± 3.55 b |
| HY | 36.74 ± 3.30 c | 36.84 ± 1.99 c |
| HYT5 | 24.58 ± 2.65 a | 24.21 ± 2.53 a |
| HYT50 | 19.32 ± 0.54 a | 19.08 ± 0.47 a |
| HYT500 | 21.53 ± 1.29 a | 22.05 ± 1.18 a |
| Quantitative Fat Analysis of Female Offspring on PND 21 | ||
| Visceral (µm) | RpWAT (µm) | |
| NS | 16.42 ± 1.55 a | 16.16 ± 1.09 a |
| HS | 61.24 ± 5.49 b | 62.31 ± 3.45 b |
| HY | 35.83 ± 3.18 c | 35.76 ± 1.98 c |
| HYT5 | 23.57 ± 2.60 a | 23.22 ± 2.56 a |
| HYT50 | 20.52 ± 1.28 a | 20.40 ± 0.68 a |
| HYT500 | 18.27 ± 1.28 a | 18.26 ± 1.78 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naomi, R.; Rusli, R.N.M.; Othman, F.; Balan, S.S.; Abidin, A.Z.; Embong, H.; Teoh, S.H.; Jasni, A.S.; Jumidil, S.H.; Matraf, K.S.Y.B.; et al. Elateriospermum tapos Yogurt Supplement in Maternal Obese Dams during Pregnancy Modulates the Body Composition of F1 Generation. Nutrients 2023, 15, 1258. https://doi.org/10.3390/nu15051258
Naomi R, Rusli RNM, Othman F, Balan SS, Abidin AZ, Embong H, Teoh SH, Jasni AS, Jumidil SH, Matraf KSYB, et al. Elateriospermum tapos Yogurt Supplement in Maternal Obese Dams during Pregnancy Modulates the Body Composition of F1 Generation. Nutrients. 2023; 15(5):1258. https://doi.org/10.3390/nu15051258
Chicago/Turabian StyleNaomi, Ruth, Rusydatul Nabila Mahmad Rusli, Fezah Othman, Santhra Segaran Balan, Azrina Zainal Abidin, Hashim Embong, Soo Huat Teoh, Azmiza Syawani Jasni, Siti Hadizah Jumidil, Khaled Salem Yaslam Ba Matraf, and et al. 2023. "Elateriospermum tapos Yogurt Supplement in Maternal Obese Dams during Pregnancy Modulates the Body Composition of F1 Generation" Nutrients 15, no. 5: 1258. https://doi.org/10.3390/nu15051258