
Sweetener System Intervention Shifted Neutrophils from Homeostasis to Priming
 , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Trial Design and Subjects
2.3. Human Blood Polymorphonuclear Neutrophil (PMN) Purification
2.4. RNA Isolation and cDNA Synthesis
2.5. RT-qPCR
2.6. Quantification of Artificial Sweeteners in Human Plasma
2.7. Gene Ontology (GO) Term-Based Network and Cluster Analysis
2.8. Spectrofluorimetric Ca2+ Influx Kinetics in Isolated PMNs In Vitro
2.9. Ca2+ Signaling Measured by Laser-Guided Flow Cytometry
2.10. Statistical Analysis
3. Results
3.1. Saccharin Upregulated Sweet Taste Receptor Gene Expression in Isolated Neutrophils In Vitro
3.2. Saccharin Upregulated Gene Expression of Neutrophil Chemokines and Their Receptors in Isolated Neutrophils In Vitro
3.3. Bio-Appearance of Sweeteners in Healthy Volunteers
3.4. Plasma-Typical Sweetener Concentrations Increased Transcript Levels of Their Cognate Taste Receptors in Isolated Neutrophils In Vitro
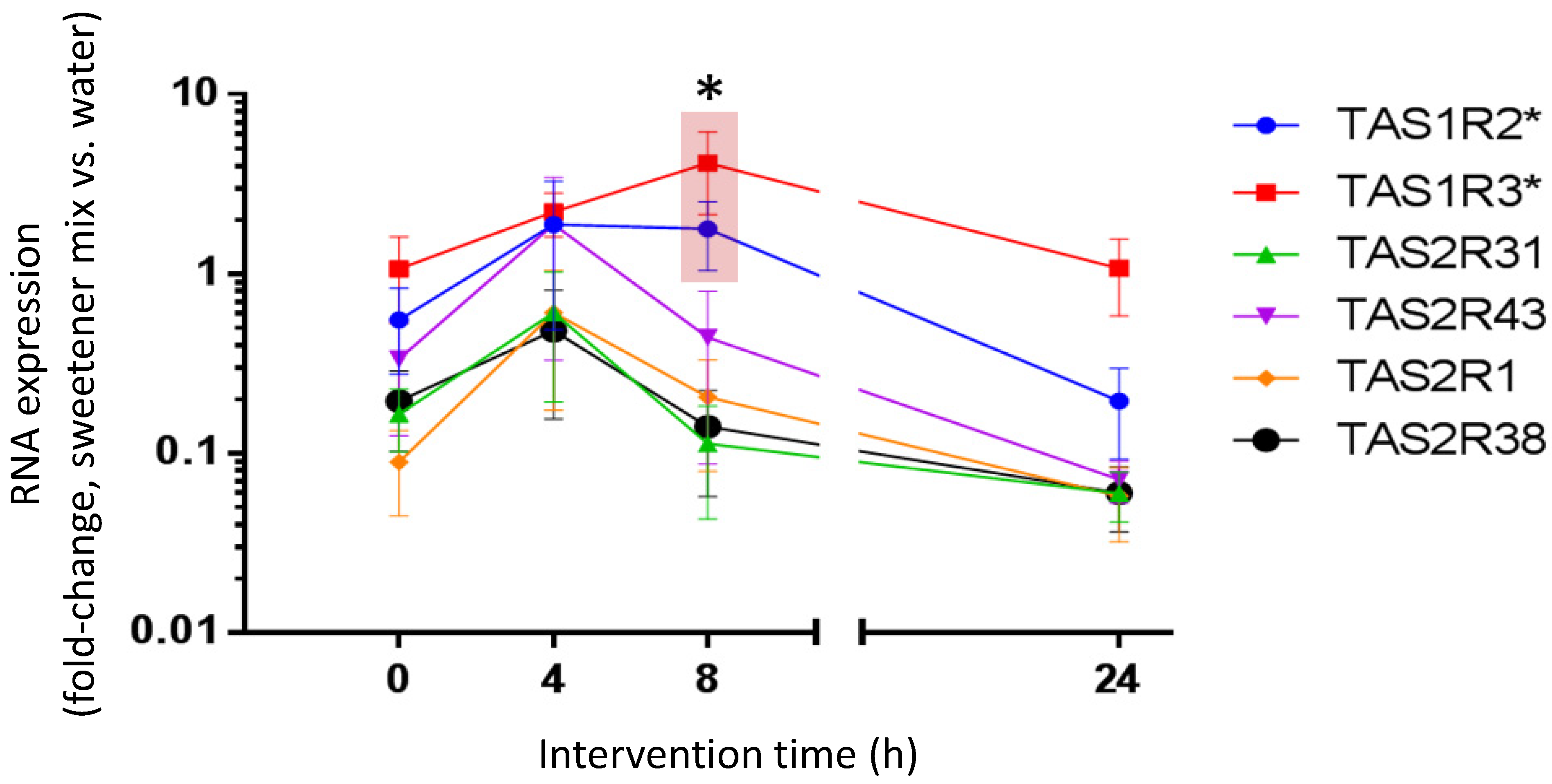
3.5. Beverage-Typical Sweetener Mix Intervention Regulated Transcript Levels of Cognate Taste Receptors in Isolated Neutrophils Ex Vivo
3.6. OPLS-DA Score Plots Reveal Reversible Differences between Water and Sweetener Intervention Groups over Time
3.7. VIP-Plots of Transcripts Indicate Post-Intervention-Specific and Functionally Diverse Gene Sets as Most Relevant Discriminants between Intervention Times
3.8. Network and Cluster Analyses Reveal That the Most Relevant Discriminants between Intervention Times Associate with Specific Functional Gene Ontology Networks
3.9. Sweetener Mix-Induced Ca2+ Influx Increased Neutrophils’ Sensitivity for fMLF
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Bes-Rastrollo, M.; Sayon-Orea, C.; Ruiz-Canela, M.; Martinez-Gonzalez, M.A. Impact of sugars and sugar taxation on body weight control: A comprehensive literature review. Obesity 2016, 24, 1410–1426. [Google Scholar] [CrossRef] [PubMed]
- Massougbodji, J.; Le Bodo, Y.; Fratu, R.; De Wals, P. Reviews examining sugar-sweetened beverages and body weight: Correlates of their quality and conclusions. Am. J. Clin. Nutr. 2014, 99, 1096–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chattopadhyay, S.; Raychaudhuri, U.; Chakraborty, R. Artificial sweeteners—A review. J. Food Sci. Technol. 2014, 51, 611–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martyn, D.; Darch, M.; Roberts, A.; Lee, H.Y.; Yaqiong Tian, T.; Kaburagi, N.; Belmar, P. Low-/No-Calorie Sweeteners: A Review of Global Intakes. Nutrients 2018, 10, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azaïs-Braesco, V.; Sluik, D.; Maillot, M.; Kok, F.; Moreno, L.A. A review of total & added sugar intakes and dietary sources in Europe. Nutr. J. 2017, 16, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connor, D.; Pang, M.; Castelnuovo, G.; Finlayson, G.; Blaak, E.; Gibbons, C.; Navas-Carretero, S.; Almiron-Roig, E.; Harrold, J.; Raben, A.; et al. A rational review on the effects of sweeteners and sweetness enhancers on appetite, food reward and metabolic/adiposity outcomes in adults. Food Funct. 2021, 12, 442–465. [Google Scholar] [CrossRef]
- Rogers, P.J.; Appleton, K.M. The effects of low-calorie sweeteners on energy intake and body weight: A systematic review and meta-analyses of sustained intervention studies. Int. J. Obes. Lond 2021, 45, 464–478. [Google Scholar] [CrossRef]
- Pang, M.D.; Goossens, G.H.; Blaak, E.E. The Impact of Artificial Sweeteners on Body Weight Control and Glucose Homeostasis. Front. Nutr. 2020, 7, 598340. [Google Scholar] [CrossRef]
- Babusyte, A.; Kotthoff, M.; Fiedler, J.; Krautwurst, D. Biogenic amines activate blood leukocytes via trace amine-associated receptors TAAR1 and TAAR2. J. Leukoc. Biol. 2013, 93, 387–394. [Google Scholar] [CrossRef]
- Clark, A.A.; Nurmukhambetova, S.; Li, X.; Munger, S.D.; Lees, J.R. Odorants specifically modulate chemotaxis and tissue retention of CD4+ T cells via cyclic adenosine monophosphate induction. J. Leukoc. Biol. 2016, 100, 699–709. [Google Scholar] [CrossRef] [Green Version]
- Csakai, A.; Smith, C.; Davis, E.; Martinko, A.; Coulup, S.; Yin, H. Saccharin derivatives as inhibitors of interferon-mediated inflammation. J. Med. Chem. 2014, 57, 5348–5355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekoff, M.; Choi, J.H.; James, A.; Dahlen, B.; Nilsson, G.; Dahlen, S.E. Bitter taste receptor (TAS2R) agonists inhibit IgE-dependent mast cell activation. J. Allergy Clin. Immunol. 2014, 134, 475–478. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.J.; Kofonow, J.M.; Rosen, P.L.; Siebert, A.P.; Chen, B.; Doghramji, L.; Xiong, G.; Adappa, N.D.; Palmer, J.N.; Kennedy, D.W.; et al. Bitter and sweet taste receptors regulate human upper respiratory innate immunity. J. Clin. Investig. 2014, 124, 1393–14052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malki, A.; Fiedler, J.; Fricke, K.; Ballweg, I.; Pfaffl, M.W.; Krautwurst, D. Class I odorant receptors, TAS1R and TAS2R taste receptors, are markers for subpopulations of circulating leukocytes. J. Leukoc. Biol. 2015, 97, 533–545. [Google Scholar] [CrossRef] [Green Version]
- Rahiman, F.; Pool, E.J. The in vitro effects of artificial and natural sweeteners on the immune system using whole blood culture assays. J. Immunoass. Immunochem. 2014, 35, 26–36. [Google Scholar] [CrossRef] [Green Version]
- Rijnierse, A.; Jeurink, P.V.; van Esch, B.C.; Garssen, J.; Knippels, L.M. Food-derived oligosaccharides exhibit pharmaceutical properties. Eur. J. Pharm. 2011, 668 (Suppl. S1), S117–S123. [Google Scholar] [CrossRef]
- Rodrigues, D.F.; Henriques, M.C.; Oliveira, M.C.; Menezes-Garcia, Z.; Marques, P.E.; Souza Dda, G.; Menezes, G.B.; Teixeira, M.M.; Ferreira, A.V. Acute intake of a high-fructose diet alters the balance of adipokine concentrations and induces neutrophil influx in the liver. J. Nutr. Biochem. 2014, 25, 388–394. [Google Scholar] [CrossRef]
- Azad, M.B.; Abou-Setta, A.M.; Chauhan, B.F.; Rabbani, R.; Lys, J.; Copstein, L.; Mann, A.; Jeyaraman, M.M.; Reid, A.E.; Fiander, M.; et al. Nonnutritive sweeteners and cardiometabolic health: A systematic review and meta-analysis of randomized controlled trials and prospective cohort studies. Can. Med. Assoc. J. 2017, 189, E929–E939. [Google Scholar] [CrossRef] [Green Version]
- Mattes, R.D.; Popkin, B.M. Nonnutritive sweetener consumption in humans: Effects on appetite and food intake and their putative mechanisms. Am. J. Clin. Nutr. 2009, 89, 26792. [Google Scholar] [CrossRef] [Green Version]
- Pepino, M.Y. Metabolic effects of non-nutritive sweeteners. Physiol. Behav. 2015, 152, 450–455. [Google Scholar] [CrossRef] [Green Version]
- Rolls, B.J. Effects of intense sweeteners on hunger, food intake, and body weight: A review. Am. J. Clin. Nutr. 1991, 53, 872–878. [Google Scholar] [CrossRef] [PubMed]
- Belloir, C.; Neiers, F.; Briand, L. Sweeteners and sweetness enhancers. Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Magnuson, B.A.; Carakostas, M.C.; Moore, N.H.; Poulos, S.P.; Renwick, A.G. Biological fate of low-calorie sweeteners. Nutr. Rev. 2016, 74, 670–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glendinning, J.I. Oral Post-Oral Actions of Low-Calorie Sweeteners: A Tale of Contradictions and Controversies. Obes. Silver Spring Md. 2018, 26 (Suppl. S3), S9–S17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colburn, W.A.; Bekersky, I.; Blumenthal, H.P. Dietary saccharin kinetics. Clin. Pharmacol. Ther. 1981, 30, 558–563. [Google Scholar] [CrossRef] [PubMed]
- Pantarotto, C.; Salmona, M.; Garattini, S. Plasma kinetics and urinary elimination of saccharin in man. Toxicol. Lett. 1981, 9, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Lasconi, C.; Pifferi, S.; Hernandez-Clavijo, A.; Merigo, F.; Cecchini, M.P.; Gonzalez-Velandia, K.Y.; Agostinelli, E.; Sbarbati, A.; Menini, A. Bitter tastants and artificial sweeteners activate a subset of epithelial cells in acute tissue slices of the rat trachea. Sci. Rep. 2019, 9, 8834. [Google Scholar] [CrossRef] [Green Version]
- Bokulich, N.A.; Blaser, M.J. A bitter aftertaste: Unintended effects of artificial sweeteners on the gut microbiome. Cell Metab. 2014, 20, 701–703. [Google Scholar] [CrossRef] [Green Version]
- Behrens, M.; Lang, T. Extra-Oral Taste Receptors-Function, Disease, and Perspectives. Front. Nutr. 2022, 9, 881177. [Google Scholar] [CrossRef]
- Buchanan, K.L.; Rupprecht, L.E.; Kaelberer, M.M.; Sahasrabudhe, A.; Klein, M.E.; Villalobos, J.A.; Liu, W.W.; Yang, A.; Gelman, J.; Park, S.; et al. The preference for sugar over sweetener depends on a gut sensor cell. Nat. Neurosci. 2022, 25, 191–200. [Google Scholar] [CrossRef]
- Suez, J.; Cohen, Y.; Valdés-Mas, R.; Mor, U.; Dori-Bachash, M.; Federici, S.; Zmora, N.; Leshem, A.; Heinemann, M.; Linevsky, R.; et al. Personalized microbiome-driven effects of non-nutritive sweeteners on human glucose tolerance. Cell 2022, 185, 3307–3328. [Google Scholar] [CrossRef] [PubMed]
- Brodin, P.; Davis, M.M. Human immune system variation. Nat. Rev. Immunol. 2017, 17, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Gannarapu, M.R.; Vasamsetti, S.B.; Punna, N.; Royya, N.K.; Pamulaparthy, S.R.; Nanubolu, J.B.; Kotamraju, S.; Banda, N. Synthesis of novel 1,2-benzothiazine 1,1-dioxide-3-ethanone oxime N-aryl acetamide ether derivatives as potent anti-inflammatory agents and inhibitors of monocyte-to-macrophage transformation. Eur. J. Med. Chem. 2014, 75, 143–150. [Google Scholar] [CrossRef]
- Ghanim, H.; Mohanty, P.; Pathak, R.; Chaudhuri, A.; Sia, C.L.; Dandona, P. Orange juice or fructose intake does not induce oxidative and inflammatory response. Diabetes Care 2007, 30, 1406–1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitsutomi, K.; Masaki, T.; Shimasaki, T.; Gotoh, K.; Chiba, S.; Kakuma, T.; Shibata, H. Effects of a nonnutritive sweetener on body adiposity and energy metabolism in mice with diet-induced obesity. Metab. Clin. Exp. 2014, 63, 69–78. [Google Scholar] [CrossRef]
- Suez, J.; Korem, T.; Zeevi, D.; Zilberman-Schapira, G.; Thaiss, C.A.; Maza, O.; Israeli, D.; Zmora, N.; Gilad, S.; Weinberger, A.; et al. Artificial sweeteners induce glucose intolerance by altering the gut microbiota. Nature 2014, 514, 181–186. [Google Scholar] [CrossRef]
- Swithers, S.E.; Martin, A.A.; Davidson, T.L. High-intensity sweeteners and energy balance. Physiol. Behav. 2010, 100, 55–62. [Google Scholar] [CrossRef] [Green Version]
- Aspinall, R.L.; Saunders, R.N.; Pautsch, W.F.; Nutting, E.F. The biological properties of aspartame. V. Effects on a variety of physiological parameters related to inflammation and metabolism. J. Environ. Pathol. Toxicol. 1980, 3, 387–395. [Google Scholar]
- Choudhary, A.K.; Pretorius, E. Revisiting the safety of aspartame. Nutr. Rev. 2017, 75, 718–730. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, A.K.; Pretorius, E. Corrigendum for “Revisiting the safety of aspartame” by Arbind Kumar Choudhary and Etheresia Pretorius Nutrition Reviews. Nutr. Rev. 2018, 76, 860. [Google Scholar] [CrossRef]
- FDA. High-Intensity Sweeteners. Available online: https://www.fda.gov/food/food-additives-petitions/high-intensity-sweeteners (accessed on 23 February 2023).
- FDA. Additional Information about High-Intensity Sweeteners Permitted for Use in Food in the United States. Available online: https://www.fda.gov/food/food-additives-petitions/additional-information-about-high-intensity-sweeteners-permitted-use-food-united-states (accessed on 23 February 2023).
- CSPI. Cyclamate (Not Legal in US). Available online: https://www.cspinet.org/article/cyclamate-not-legal-us (accessed on 17 February 2022).
- Leth, T.; Fabricius, N.; Fagt, S. Estimated intake of intense sweeteners from non-alcoholic beverages in Denmark. Food Addit. Contam. 2007, 24, 227–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horne, J.; Lawless, H.T.; Speirs, W.; Sposato, D. Bitter taste of saccharin and acesulfame-K. Chem. Senses 2002, 27, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, C.; Bufe, B.; Winnig, M.; Hofmann, T.; Frank, O.; Behrens, M.; Lewtschenko, T.; Slack, J.P.; Ward, C.D.; Meyerhof, W. Bitter taste receptors for saccharin and acesulfame K. J. Neurosci. 2004, 24, 10260–10265. [Google Scholar] [CrossRef] [Green Version]
- Roudnitzky, N.; Bufe, B.; Thalmann, S.; Kuhn, C.; Gunn, H.C.; Xing, C.; Crider, B.P.; Behrens, M.; Meyerhof, W.; Wooding, S.P. Genomic, genetic and functional dissection of bitter taste responses to artificial sweeteners. Hum. Mol. Genet. 2011, 20, 3437–3449. [Google Scholar] [CrossRef]
- Schiffman, S.S.; Booth, B.J.; Losee, M.L.; Pecore, S.D.; Warwick, Z.S. Bitterness of sweeteners as a function of concentration. Brain Res. Bull. 1995, 36, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Behrens, M.; Blank, K.; Meyerhof, W. Blends of Non-caloric Sweeteners Saccharin and Cyclamate Show Reduced Off-Taste due to TAS2R Bitter Receptor Inhibition. Cell Chem. Biol. 2017, 24, 1199–1204. [Google Scholar] [CrossRef] [Green Version]
- National Library of Medicine. Saccharin. In Drugs and Lactation Database (LactMed), 16-09-2016 eds.; National Library of Medicine (NLM): Bethesda, MD, USA, 2016. [Google Scholar]
- Marcinek, P.; Geithe, C.; Krautwurst, D. Chemosensory G Protein-Coupled Receptors (GPCR) in Blood Leukocytes. In Taste and Smell Receptors; Krautwurst, D., Ed.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 1–23. [Google Scholar]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Frank, O.; Kreissl, J.K.; Daschner, A.; Hofmann, T. Accurate determination of reference materials and natural isolates by means of quantitative 1H NMR spectroscopy. J. Agric. Food Chem. 2014, 62, 2506–2515. [Google Scholar] [CrossRef]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinform. Oxf. Engl. 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Maere, S.; Heymans, K.; Kuiper, M. BiNGO: A Cytoscape plugin to assess overrepresentation of gene ontology categories in biological networks. Bioinform. Oxf. Engl. 2005, 21, 3448–3449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- UniProt_Consortium. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Virgilio, F.; Steinberg, T.H.; Swanson, J.A.; Silverstein, S.C. Fura-2 secretion and sequestration in macrophages. A blocker of organic anion transport reveals that these processes occur via a membrane transport system for organic anions. J. Immunol. Baltim. Md. 1988, 140, 915–920. [Google Scholar] [CrossRef]
- Merritt, J.E.; McCarthy, S.A.; Davies, M.P.; Moores, K.E. Use of fluo-3 to measure cytosolic Ca2+ in platelets and neutrophils. Loading cells with the dye, calibration of traces, measurements in the presence of plasma, and buffering of cytosolic Ca2+. Biochem. J. 1990, 269, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Qiagen. Custom RT2 Profiler PCR Array Data Analysis Spreadsheet 1808; Template with Instructions for Analyzing Custom RT2 Profiler PCR Array Results (MS-EXCEL File, xlsx); Qiagen: Hilden, Germany, 2018. [Google Scholar]
- Wickham, H. tidyverse: Easily Install and Load the ‘Tidyverse’; R Package 2017; RStudio: Vienna, Austria, 2017. [Google Scholar]
- Thevenot, E.A.; Roux, A.; Xu, Y.; Ezan, E.; Junot, C. Analysis of the Human Adult Uri-nary Metabolome Variations with Age, Body Mass Index, and Gender by Implementing a Comprehensive Workflow for Univariate and OPLS Statistical Analyses. J. Proteome Res. 2015, 14, 3322–3335. [Google Scholar] [CrossRef]
- Wilke, C.O. Cowplot; R Package 2019; RStudio: Vienna, Austria, 2019. [Google Scholar]
- Wiklund, S.; Johansson, E.; Sjostrom, L.; Mellerowicz, E.J.; Edlund, U.; Shockcor, J.P.; Gottfries, J.; Moritz, T.; Trygg, J. Visualization of GC/TOF-MS-based metabolomics data for identification of biochemically interesting compounds using OPLS class models. Anal. Chem. 2008, 80, 115–122. [Google Scholar] [CrossRef]
- Comerford, I.; Harata-Lee, Y.; Bunting, M.D.; Gregor, C.; Kara, E.E.; McColl, S.R. A myriad of functions and complex regulation of the CCR7/CCL19/CCL21 chemokine axis in the adaptive immune system. Cytokine Growth Factor Rev. 2013, 24, 269–283. [Google Scholar] [CrossRef]
- Priego, T.; Sanchez, J.; Pico, C.; Ahrens, W.; De Henauw, S.; Kourides, Y.; Lissner, L.; Molnar, D.; Moreno, L.A.; Russo, P.; et al. TAS1R3 and UCN2 Transcript Levels in Blood Cells Are Associated With Sugary and Fatty Food Consumption in Children. J. Clin. Endocrinol. Metab. 2015, 100, 3556–3564. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, A.; Stockwell, P.A.; Rodger, E.J.; Duncan, E.J.; Parry, M.F.; Weeks, R.J.; Morison, I.M. Genome-wide DNA methylation map of human neutrophils reveals widespread inter-individual epigenetic variation. Sci. Rep. 2015, 5, 17328. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, A.; Stockwell, P.A.; Rodger, E.J.; Morison, I.M. Genome-scale DNA methylome and transcriptome profiling of human neutrophils. Sci. Data 2016, 3, 160019. [Google Scholar] [CrossRef] [Green Version]
- Galindo-Cuspinera, V.; Winnig, M.; Bufe, B.; Meyerhof, W.; Breslin, P.A. A TAS1R receptor-based explanation of sweet ‘water-taste’. Nature 2006, 441, 354–357. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Staszewski, L.; Xu, H.; Durick, K.; Zoller, M.; Adler, E. Human receptors for sweet and umami taste. Proc. Natl. Acad. Sci. USA 2002, 99, 4692–4696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winnig, M.; Kuhn, C.; Frank, O.; Bufe, B.; Behrens, M.; Hofmann, T.; Meyerhof, W. Saccharin: Artificial Sweetener, Bitter Tastant, and Sweet Taste Inhibitor. In Sweetness and Sweeteners; American Chemical Society: Washington, DC, USA, 2008; Volume 979, pp. 230–240. [Google Scholar]
- Zhao, G.Q.; Zhang, Y.; Hoon, M.A.; Chandrashekar, J.; Erlenbach, I.; Ryba, N.J.; Zuker, C.S. The receptors for mammalian sweet and umami taste. Cell 2003, 115, 255–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panaro, M.A.; Mitolo, V. Cellular responses to FMLP challenging: A mini-review. Immunopharmacol. Immunotoxicol. 1999, 21, 397–419. [Google Scholar] [CrossRef] [PubMed]
- Meyerhof, W.; Batram, C.; Kuhn, C.; Brockhoff, A.; Chudoba, E.; Bufe, B.; Appendino, G.; Behrens, M. The molecular receptive ranges of human TAS2R bitter taste receptors. Chem. Senses 2010, 35, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Bouwmeester, T.; Bauch, A.; Ruffner, H.; Angrand, P.O.; Bergamini, G.; Croughton, K.; Cruciat, C.; Eberhard, D.; Gagneur, J.; Ghidelli, S.; et al. A physical and functional map of the human TNF-alpha/NF-kappa B signal transduction pathway. Nat. Cell Biol. 2004, 6, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Kornerup, K.N.; Salmon, G.P.; Pitchford, S.C.; Liu, W.L.; Page, C.P. Circulating platelet-neutrophil complexes are important for subsequent neutrophil activation and migration. J. Appl. Physiol. Bethesda Md. 2010, 109, 758–767. [Google Scholar] [CrossRef] [PubMed]
- Siegemund, S.; Sauer, K. Balancing pro- and anti-inflammatory TLR4 signaling. Nat. Immunol. 2012, 13, 1031–1033. [Google Scholar] [CrossRef]
- Fumagalli, M.; Lecca, D.; Coppolino, G.T.; Parravicini, C.; Abbracchio, M.P. Pharmacological Properties and Biological Functions of the GPR17 Receptor, a Potential Target for Neuro-Regenerative Medicine. Adv. Exp. Med. Biol. 2017, 1051, 169–192. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Zepeda, E.A.; Rothenberg, M.E.; Ownbey, R.T.; Celestin, J.; Leder, P.; Luster, A.D. Human eotaxin is a specific chemoattractant for eosinophil cells and provides a new mechanism to explain tissue eosinophilia. Nat. Med. 1996, 2, 449–456. [Google Scholar] [CrossRef]
- Nakayama, T.; Watanabe, Y.; Oiso, N.; Higuchi, T.; Shigeta, A.; Mizuguchi, N.; Katou, F.; Hashimoto, K.; Kawada, A.; Yoshie, O. Eotaxin-3/CC chemokine ligand 26 is a functional ligand for CX3CR1. J. Immunol. Baltim. Md. 2010, 185, 6472–6479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petkovic, V.; Moghini, C.; Paoletti, S.; Uguccioni, M.; Gerber, B. Eotaxin-3/CCL26 is a natural antagonist for CC chemokine receptors 1 and 5. A human chemokine with a regulatory role. J. Biol. Chem. 2004, 279, 23357–23363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Regan, A.; Berman, J.S. Osteopontin: A key cytokine in cell-mediated and granulomatous inflammation. Int. J. Exp. Pathol. 2000, 81, 373–390. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, L.A.; Golenbock, D.; Bowie, A.G. The history of Toll-like receptors—Redefining innate immunity. Nat. Rev. Immunol. 2013, 13, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Hann, J.; Bueb, J.L.; Tolle, F.; Bréchard, S. Calcium signaling and regulation of neutrophil functions: Still a long way to go. J. Leukoc. Biol. 2020, 107, 285–297. [Google Scholar] [CrossRef]
- Hubner, K.; Surovtsova, I.; Yserentant, K.; Hansch, M.; Kummer, U. Ca2+ dynamics correlates with phenotype and function in primary human neutrophils. Biophys. Chem. 2013, 184, 116–125. [Google Scholar] [CrossRef]
- Watson, F.; Edwards, S.W. Stimulation of primed neutrophils by soluble immune complexes: Priming leads to enhanced intracellular Ca2+ elevations, activation of phospholipase D, and activation of the NADPH oxidase. Biochem. Biophys. Res. Commun. 1998, 247, 819–826. [Google Scholar] [CrossRef]
- Vogt, K.L.; Summers, C.; Chilvers, E.R.; Condliffe, A.M. Priming and de-priming of neutrophil responses in vitro and in vivo. Eur. J. Clin. Investig. 2018, 48 (Suppl. S2), e12967. [Google Scholar] [CrossRef]
- Chen, L.W.; Jan, C.R. Mechanisms and modulation of formyl-methionyl-leucyl-phenylalanine (fMLP)-induced Ca2+ mobilization in human neutrophils. Int. Immunopharmacol. 2001, 1, 1341–1349. [Google Scholar] [CrossRef]
- Clemens, R.A.; Lowell, C.A. Store-operated calcium signaling in neutrophils. J. Leukoc. Biol. 2015, 98, 497–502. [Google Scholar] [CrossRef] [Green Version]
- Pettit, E.J.; Hallett, M.B. Two distinct Ca2+ storage and release sites in human neutrophils. J. Leukoc. Biol. 1998, 63, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Krause, K.H.; Campbell, K.P.; Welsh, M.J.; Lew, D.P. The calcium signal and neutrophil activation. Clin. Biochem. 1990, 23, 159–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demaurex, N.; Saul, S. The role of STIM proteins in neutrophil functions. J. Physiol. 2018, 596, 2699–2708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Migeotte, I.; Communi, D.; Parmentier, M. Formyl peptide receptors: A promiscuous subfamily of G protein-coupled receptors controlling immune responses. Cytokine Growth Factor Rev. 2006, 17, 501–519. [Google Scholar] [CrossRef]
- Dahlgren, C.; Gabl, M.; Holdfeldt, A.; Winther, M.; Forsman, H. Basic characteristics of the neutrophil receptors that recognize formylated peptides, a danger-associated molecular pattern generated by bacteria and mitochondria. Biochem. Pharmacol. 2016, 114, 22–39. [Google Scholar] [CrossRef]
- Zhuang, Y.; Wang, L.; Guo, J.; Sun, D.; Wang, Y.; Liu, W.; Xu, H.E.; Zhang, C. Molecular recognition of formylpeptides and diverse agonists by the formylpeptide receptors FPR1 and FPR2. Nat. Commun. 2022, 13, 1054. [Google Scholar] [CrossRef]
- Vainer, B.; Lamberth, K.; Brimnes, J.; Nielsen, O.H.; Claësson, M.H. Ca2+ response in neutrophils after exposure to bacterial N-formyl-methionyl-leucyl-phenylalanine: Delayed response in ulcerative colitis. Eur. J. Gastroenterol. Hepatol. 2003, 15, 267–273. [Google Scholar] [CrossRef]
- Dahlgren, C.; Holdfeldt, A.; Lind, S.; Mårtensson, J.; Gabl, M.; Björkman, L.; Sundqvist, M.; Forsman, H. Neutrophil Signaling That Challenges Dogmata of G Protein-Coupled Receptor Regulated Functions. ACS Pharm. Transl. Sci. 2020, 3, 203–220. [Google Scholar] [CrossRef]
- Jiang, P.; Cui, M.; Zhao, B.; Liu, Z.; Snyder, L.A.; Benard, L.M.; Osman, R.; Margolskee, R.F.; Max, M. Lactisole interacts with the transmembrane domains of human T1R3 to inhibit sweet taste. J. Biol. Chem. 2005, 280, 15238–15246. [Google Scholar] [CrossRef] [Green Version]
- Andersen, G.; Kahlenberg, K.; Krautwurst, D.; Somoza, V. [6]-Gingerol facilitates CXCL8 secretion and ROS production in primary human neutrophils by targeting the TRPV1 channel. Mol. Nutr. Food Res. 2022, 67, 2200434. [Google Scholar] [CrossRef]
- Riera, C.E.; Vogel, H.; Simon, S.A.; le Coutre, J. Artificial sweeteners and salts producing a metallic taste sensation activate TRPV1 receptors. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R626–R634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, H.J.; Kokrashvili, Z.; Theodorakis, M.J.; Carlson, O.D.; Kim, B.J.; Zhou, J.; Kim, H.H.; Xu, X.; Chan, S.L.; Juhaszova, M.; et al. Gut-expressed gustducin and taste receptors regulate secretion of glucagon-like peptide-1. Proc. Natl. Acad. Sci. USA 2007, 104, 15069–15074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margolskee, R.F.; Dyer, J.; Kokrashvili, Z.; Salmon, K.S.; Ilegems, E.; Daly, K.; Maillet, E.L.; Ninomiya, Y.; Mosinger, B.; Shirazi-Beechey, S.P. T1R3 and gustducin in gut sense sugars to regulate expression of Na+-glucose cotransporter 1. Proc. Natl. Acad. Sci. USA 2007, 104, 15075–15080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, R.J.; Xiong, G.; Kofonow, J.M.; Chen, B.; Lysenko, A.; Jiang, P.; Abraham, V.; Doghramji, L.; Adappa, N.D.; Palmer, J.N.; et al. T2R38 taste receptor polymorphisms underlie susceptibility to upper respiratory infection. J. Clin. Investig. 2012, 122, 4145–4159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, X.C.; Chen, Z.H.; Xue, J.B.; Zhao, D.X.; Lu, C.; Li, Y.H.; Li, S.M.; Du, Y.W.; Liu, Q.; Wang, P.; et al. Infection by the parasitic helminth Trichinella spiralis activates a Tas2r-mediated signaling pathway in intestinal tuft cells. Proc. Natl. Acad. Sci. USA 2019, 116, 5564–5569. [Google Scholar] [CrossRef] [Green Version]
- Saunders, C.I.; Kunde, D.A.; Crawford, A.; Geraghty, D.P. Expression of transient receptor potential vanilloid 1 (TRPV1) and 2 (TRPV2) in human peripheral blood. Mol. Immunol. 2007, 44, 1429–1435. [Google Scholar] [CrossRef]
- Wang, J.P.; Tseng, C.S.; Sun, S.P.; Chen, Y.S.; Tsai, C.R.; Hsu, M.F. Capsaicin stimulates the non-store-operated Ca2+ entry but inhibits the store-operated Ca2+ entry in neutrophils. Toxicol. Appl. Pharmacol. 2005, 209, 134–144. [Google Scholar] [CrossRef]
- Heiner, I.; Eisfeld, J.; Luckhoff, A. Role and regulation of TRP channels in neutrophil granulocytes. Cell Calcium. 2003, 33, 533–540. [Google Scholar] [CrossRef]
- Pantaler, E.; Luckhoff, A. Inhibitors of TRP channels reveal stimulus-dependent differential activation of Ca2+ influx pathways in human neutrophil granulocytes. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2009, 380, 497–507. [Google Scholar] [CrossRef]
- Maekawa, A.; Xing, W.; Austen, K.F.; Kanaoka, Y. GPR17 regulates immune pulmonary inflammation induced by house dust mites. J. Immunol. Baltim. Md. 2010, 185, 1846–1854. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.S.; Lai, J.J.; Lukacs, N.W.; Kunkel, S.L. Granulocyte-macrophage colony stimulating factor up-regulates CCR1 in human neutrophils. J. Immunol. Baltim. Md. 2001, 166, 1178–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartl, D.; Krauss-Etschmann, S.; Koller, B.; Hordijk, P.L.; Kuijpers, T.W.; Hoffmann, F.; Hector, A.; Eber, E.; Marcos, V.; Bittmann, I.; et al. Infiltrated neutrophils acquire novel chemokine receptor expression and chemokine responsiveness in chronic inflammatory lung diseases. J. Immunol. Baltim. Md. 2008, 181, 8053–8067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudd, J.M.; Pulavendran, S.; Ashar, H.K.; Ritchey, J.W.; Snider, T.A.; Malayer, J.R.; Marie, M.; Chow, V.T.K.; Narasaraju, T. Neutrophils Induce a Novel Chemokine Receptors Repertoire During Influenza Pneumonia. Front. Cell. Infect. Microbiol. 2019, 9, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saini, V.; Marchese, A.; Majetschak, M. CXC chemokine receptor 4 is a cell surface receptor for extracellular ubiquitin. J. Biol. Chem. 2010, 285, 15566–15576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miralda, I.; Uriarte, S.M.; McLeish, K.R. Multiple Phenotypic Changes Define Neutrophil Priming. Front. Cell. Infect. Microbiol. 2017, 7, 217. [Google Scholar] [CrossRef] [Green Version]
- Sabroe, I.; Prince, L.R.; Jones, E.C.; Horsburgh, M.J.; Foster, S.J.; Vogel, S.N.; Dower, S.K.; Whyte, M.K. Selective roles for Toll-like receptor (TLR)2 and TLR4 in the regulation of neutrophil activation and life span. J. Immunol. Baltim. Md. 2003, 170, 5268–5275. [Google Scholar] [CrossRef] [Green Version]
- Aksoy, E.; Taboubi, S.; Torres, D.; Delbauve, S.; Hachani, A.; Whitehead, M.A.; Pearce, W.P.; Berenjeno, I.M.; Nock, G.; Filloux, A.; et al. The p110delta isoform of the kinase PI(3)K controls the subcellular compartmentalization of TLR4 signaling and protects from endotoxic shock. Nat. Immunol. 2012, 13, 1045–1054. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Bao, Z.; Tang, P.; Gong, W.; Yoshimura, T.; Wang, J.M. Chemokines in homeostasis and diseases. Cell. Mol. Immunol. 2018, 15, 324–334. [Google Scholar] [CrossRef]
- Forster, R.; Davalos-Misslitz, A.C.; Rot, A. CCR7 and its ligands: Balancing immunity and tolerance. Nat. Rev. Immunol. 2008, 8, 362–371. [Google Scholar] [CrossRef]
- Weber, G.F.; Cantor, H. The immunology of Eta-1/osteopontin. Cytokine Growth Factor Rev. 1996, 7, 241–248. [Google Scholar] [CrossRef]
- Sylvetsky, A.C.; Sen, S.; Merkel, P.; Dore, F.; Stern, D.B.; Henry, C.J.; Cai, H.; Walter, P.J.; Crandall, K.A.; Rother, K.I.; et al. Consumption of Diet Soda Sweetened with Sucralose and Acesulfame-Potassium Alters Inflammatory Transcriptome Pathways in Females with Overweight and Obesity. Mol. Nutr. Food Res. 2020, 64, e1901166. [Google Scholar] [CrossRef] [PubMed]
- Feng, P.; Jyotaki, M.; Kim, A.; Chai, J.; Simon, N.; Zhou, M.; Bachmanov, A.A.; Huang, L.; Wang, H. Regulation of bitter taste responses by tumor necrosis factor. Brain Behav. Immun. 2015, 49, 32–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Önnheim, K.; Christenson, K.; Gabl, M.; Burbiel, J.C.; Müller, C.E.; Oprea, T.I.; Bylund, J.; Dahlgren, C.; Forsman, H. A novel receptor cross-talk between the ATP receptor P2Y2 and formyl peptide receptors reactivates desensitized neutrophils to produce superoxide. Exp. Cell Res. 2014, 323, 209–217. [Google Scholar] [CrossRef] [PubMed]
- McCormick, B.; Chu, J.Y.; Vermeren, S. Cross-talk between Rho GTPases and PI3K in the neutrophil. Small GTPases 2019, 10, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Mol, S.; Hafkamp, F.M.J.; Varela, L.; Simkhada, N.; Taanman-Kueter, E.W.; Tas, S.W.; Wauben, M.H.M.; Groot Kormelink, T.; de Jong, E.C. Efficient Neutrophil Activation Requires Two Simultaneous Activating Stimuli. Int. J. Mol. Sci. 2021, 22, 106. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (h) | Saccharin | Acesulfame-K | Cyclamate |
|---|---|---|---|
| Plasma Concentration (µM) | |||
| 0 | 0.01 ± 0.01 | <0.01 | <0.01 |
| 4 | 0.85 ± 0.29 | 0.39 ± 0.18 | 1.38 ± 0.62 |
| 8 | 0.33 ± 0.17 | 0.14 ± 0.06 | 0.55 ± 0.25 |
| 24 | 0.07 ± 0.04 | <0.01 | 0.22 ± 0.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skurk, T.; Krämer, T.; Marcinek, P.; Malki, A.; Lang, R.; Dunkel, A.; Krautwurst, T.; Hofmann, T.F.; Krautwurst, D. Sweetener System Intervention Shifted Neutrophils from Homeostasis to Priming. Nutrients 2023, 15, 1260. https://doi.org/10.3390/nu15051260
Skurk T, Krämer T, Marcinek P, Malki A, Lang R, Dunkel A, Krautwurst T, Hofmann TF, Krautwurst D. Sweetener System Intervention Shifted Neutrophils from Homeostasis to Priming. Nutrients. 2023; 15(5):1260. https://doi.org/10.3390/nu15051260
Chicago/Turabian StyleSkurk, Thomas, Tamara Krämer, Patrick Marcinek, Agne Malki, Roman Lang, Andreas Dunkel, Tiffany Krautwurst, Thomas F. Hofmann, and Dietmar Krautwurst. 2023. "Sweetener System Intervention Shifted Neutrophils from Homeostasis to Priming" Nutrients 15, no. 5: 1260. https://doi.org/10.3390/nu15051260