
Disentangling the Complexity of Nutrition, Frailty and Gut Microbial Pathways during Aging: A Focus on Hippuric Acid
 ,
,
Abstract
:1. Introduction
2. Urinary Hippuric Acid as a Nutritional Marker
3. Hippuric Acid Metabolism in Aging and Age-Related Conditions
3.1. The Physiology of Hippuric Acid in Aging
3.2. The Role of Chronic Kidney Disease
3.3. The Role of Age-Related Gut Microbiota Changes
4. Hippuric Acid in Physical Frailty and Sarcopenia
5. Hippuric Acid and Cognition in Older Adults
6. Hippuric Acid and Other Age-Related Chronic Conditions
7. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Picca, A.; Calvani, R.; Cesari, M.; Landi, F.; Bernabei, R.; Coelho-Júnior, H.J.; Marzetti, E. Biomarkers of physical frailty and sarcopenia: Coming up to the place? Int. J. Mol. Sci. 2020, 21, 5635. [Google Scholar] [CrossRef] [PubMed]
- Rivero-Segura, N.A.; Bello-Chavolla, O.Y.; Barrera-Vázquez, O.S.; Gutierrez-Robledo, L.M.; Gomez-Verjan, J.C. Promising biomarkers of human aging: In search of a multi-omics panel to understand the aging process from a multidimensional perspective. Ageing Res. Rev. 2020, 64, 101164. [Google Scholar] [CrossRef] [PubMed]
- De Simone, G.; Balducci, C.; Forloni, G.; Pastorelli, R.; Brunelli, L. Hippuric acid: Could become a barometer for frailty and geriatric syndromes? Ageing Res. Rev. 2021, 72, 101466. [Google Scholar] [CrossRef] [PubMed]
- Lees, H.J.; Swann, J.R.; Wilson, I.D.; Nicholson, J.K.; Holmes, E. Hippurate: The natural history of a mammalian-microbial cometabolite. J. Proteome Res. 2013, 12, 1527–1546. [Google Scholar] [CrossRef] [PubMed]
- Geraldino, B.R.; Nunes, R.F.N.; Gomes, J.B.; da Poça, K.S.; Giardini, I.; Silva, P.V.B.; Souza, H.P.; Otero, U.B.; Sarpa, M. Evaluation of exposure to toluene and xylene in gasoline station workers. Adv. Prev. Med. 2021, 2021, 5553633. [Google Scholar] [CrossRef]
- Oginawati, K.; Anka, A.A.H.; Susetyo, S.H.; Febriana, S.A.; Tanziha, I.; Prakoeswa, C.R.S. Urinary hippuric acid level as a biological indicator of toluene exposure on batik workers. Heliyon 2021, 7, e07775. [Google Scholar] [CrossRef]
- Wu, L.; Xie, X.; Liang, T.; Ma, J.; Yang, L.; Yang, J.; Li, L.; Xi, Y.; Li, H.; Zhang, J.; et al. Integrated multi-omics for novel aging biomarkers and antiaging targets. Biomolecules 2021, 12, 39. [Google Scholar] [CrossRef]
- Dey, P. Gut microbiota in phytopharmacology: A comprehensive overview of concepts, reciprocal interactions, biotransformations and mode of actions. Pharmacol. Res. 2019, 147, 104367. [Google Scholar] [CrossRef]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.E.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (poly)phenolics in human health: Structures, bioavailability, and evidence of protective effects against chronic diseases. Antioxid. Redox Signal. 2013, 18, 1818–1892. [Google Scholar] [CrossRef] [Green Version]
- Luca, S.V.; Macovei, I.; Bujor, A.; Miron, A.; Skalicka-Woźniak, K.; Aprotosoaie, A.C.; Trifan, A. Bioactivity of dietary polyphenols: The role of metabolites. Crit. Rev. Food Sci. Nutr. 2020, 60, 626–659. [Google Scholar] [CrossRef]
- Pruss, K.M.; Chen, H.; Liu, Y.; Van Treuren, W.; Higginbottom, S.K.; Jarman, J.B.; Fischer, C.R.; Mak, J.; Wong, B.; Cowan, T.M.; et al. Host-microbe co-metabolism via MCAD generates circulating metabolites including hippuric acid. Nat. Commun. 2023, 14, 512. [Google Scholar] [CrossRef]
- Pero, R.W. Health consequences of catabolic synthesis of hippuric acid in humans. Curr. Clin. Pharmacol. 2010, 5, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Olthof, M.R.; Hollman, P.C.H.; Buijsman, M.N.C.P.; van Amelsvoort, J.M.M.; Katan, M.B. Chlorogenic acid, quercetin-3-rutinoside and black tea phenols are extensively metabolized in humans. J. Nutr. 2003, 133, 1806–1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Temellini, A.; Mogavero, S.; Giulianotti, P.C.; Pietrabissa, A.; Mosca, F.; Pacifici, G.M. Conjugation of benzoic acid with glycine in human liver and kidney: A study on the interindividual variability. Xenobiotica 1993, 23, 1427–1433. [Google Scholar] [CrossRef] [PubMed]
- Vetrani, C.; Rivellese, A.A.; Annuzzi, G.; Adiels, M.; Borén, J.; Mattila, I.; Orešič, M.; Aura, A.M. Metabolic transformations of dietary polyphenols: Comparison between in vitro colonic and hepatic models and in vivo urinary metabolites. J. Nutr. Biochem. 2016, 33, 111–118. [Google Scholar] [CrossRef]
- del Olmo, A.; Calzada, J.; Nuñez, M. Benzoic acid and its derivatives as naturally occurring compounds in foods and as additives: Uses, exposure, and controversy. Crit. Rev. Food Sci. Nutr. 2017, 57, 3084–3103. [Google Scholar] [CrossRef] [PubMed]
- Deltombe, O.; Van Biesen, W.; Glorieux, G.; Massy, Z.; Dhondt, A.; Eloot, S. Exploring protein binding of uremic toxins in patients with different stages of chronic kidney disease and during hemodialysis. Toxins 2015, 7, 3933–3946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, Y.J.; Sidor, N.A.; Tonial, N.C.; Che, A.; Urquhart, B.L. Uremic toxins in the progression of chronic kidney disease and cardiovascular disease: Mechanisms and therapeutic targets. Toxins 2021, 13, 142. [Google Scholar] [CrossRef]
- Edwards, S.J.; Carter, S.; Nicholson, T.; Allen, S.L.; Morgan, P.T.; Jones, S.W.; Rendeiro, C.; Breen, L. (-)-Epicatechin and its colonic metabolite hippuric acid protect against dexamethasone-induced atrophy in skeletal muscle cells. J. Nutr. Biochem. 2022, 110, 109150. [Google Scholar] [CrossRef]
- Bitner, B.F.; Ray, J.D.; Kener, K.B.; Herring, J.A.; Tueller, J.A.; Johnson, D.K.; Tellez Freitas, C.M.; Fausnacht, D.W.; Allen, M.E.; Thomson, A.H.; et al. Common gut microbial metabolites of dietary flavonoids exert potent protective activities in β-cells and skeletal muscle cells. J. Nutr. Biochem. 2018, 62, 95–107. [Google Scholar] [CrossRef]
- Ohtsuki, S.; Asaba, H.; Takanaga, H.; Deguchi, T.; Hosoya, K.; Otagiri, M.; Terasaki, T. Role of blood-brain barrier organic anion transporter 3 (OAT3) in the efflux of indoxyl sulfate, a uremic toxin: Its involvement in neurotransmitter metabolite clearance from the brain. J. Neurochem. 2002, 83, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Wang, X.; Liu, X.; Wang, L.; Ren, F.; Wang, X.; Leng, X. Hippuric acid promotes renal fibrosis by disrupting redox homeostasis via facilitation of BRF2-KEAP1-CUL3 interactions in chronic kidney disease. Antioxidants 2020, 9, 783. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Wei, R.; Wang, Y.; Su, T.; Li, P.; Chen, X. The uremic toxin hippurate promotes endothelial dysfunction via the activation of Drp1-mediated mitochondrial fission. Redox Biol. 2018, 16, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Shang, F.; Wang, S.C.; Hsi, C.Y.; Miao, Y.; Martin, M.; Yin, Y.; Wu, C.C.; Wang, Y.T.; Wu, G.; Chien, S.; et al. MicroRNA-92a mediates endothelial dysfunction in CKD. J. Am. Soc. Nephrol. 2017, 28, 3251–3261. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Adiga, A.; Novack, J.; Etinger, A.; Chinitz, L.; Slater, J.; de Loor, H.; Meijers, B.; Holzman, R.S.; Lowenstein, J. The renal transport of hippurate and protein-bound solutes. Physiol. Rep. 2020, 8, e14349. [Google Scholar] [CrossRef] [Green Version]
- Leri, M.; Scuto, M.; Ontario, M.L.; Calabrese, V.; Calabrese, E.J.; Bucciantini, M.; Stefani, M. Healthy effects of plant polyphenols: Molecular mechanisms. Int. J. Mol. Sci. 2020, 21, 1250. [Google Scholar] [CrossRef] [Green Version]
- Caruso, G.; Godos, J.; Privitera, A.; Lanza, G.; Castellano, S.; Chillemi, A.; Bruni, O.; Ferri, R.; Caraci, F.; Grosso, G. Phenolic acid and prevention of cognitive decline: Polyphenols with a neuroprotective role in cognitive disorders and Alzheimer’s disease. Nutrients 2022, 14, 819. [Google Scholar] [CrossRef]
- Meccariello, R.; D’Angelo, S. Impact of polyphenolic-food on longevity: An elixir of life. An overview. Antioxidants 2021, 10, 507. [Google Scholar] [CrossRef]
- Huffman, D.M.; Schafer, M.J.; LeBrasseur, N.K. Energetic interventions for healthspan and resiliency with aging. Exp. Gerontol. 2016, 86, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Van Sumere, C.F.; Teuchy, H.; Pé, H.; Verbeke, R.; Bekaert, J. Quantitative investigation on the hippuric acid formation in healthy and diseased individuals. Clin. Chim. Acta 1969, 26, 85–88. [Google Scholar] [CrossRef]
- McMahon, T.F.; Diliberto, J.J.; Birnbaum, L.S. Age-related changes in the disposition of benzyl acetate. A model compound for glycine conjugation. Drug Metab. Dispos. 1989, 17, 506–512. [Google Scholar] [PubMed]
- Liu, H.; Garrett, T.J.; Su, Z.; Khoo, C.; Zhao, S.; Gu, L. Modifications of the urinary metabolome in young women after cranberry juice consumption were revealed using the UHPLC-Q-orbitrap-HRMS-based metabolomics approach. Food Funct. 2020, 11, 2466–2476. [Google Scholar] [CrossRef] [PubMed]
- Feliciano, R.P.; Boeres, A.; Massacessi, L.; Istas, G.; Ventura, M.R.; Dos Santos, C.N.; Heiss, C.; Rodriguez-Mateos, A. Identification and quantification of novel cranberry-derived plasma and urinary (poly)phenols. Arch. Biochem. Biophys. 2016, 599, 31–41. [Google Scholar] [CrossRef]
- Pereira-Caro, G.; Borges, G.; van der Hooft, J.; Clifford, M.N.; Del Rio, D.; Lean, M.E.J.; Roberts, S.A.; Kellerhals, M.B.; Crozier, A. Orange juice (poly)phenols are highly bioavailable in humans. Am. J. Clin. Nutr. 2014, 100, 1378–1384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Mello, V.D.; Lankinen, M.A.; Lindström, J.; Puupponen-Pimiä, R.; Laaksonen, D.E.; Pihlajamäki, J.; Lehtonen, M.; Uusitupa, M.; Tuomilehto, J.; Kolehmainen, M.; et al. Fasting serum hippuric acid is elevated after bilberry (Vaccinium mirtillus) consumption and associates with improvement of fasting serum glucose levels and insulin secretion in persons at high risk of developing type 2 diabetes. Mol. Nutr. Food Res. 2017, 61, 170019. [Google Scholar] [CrossRef]
- Curtis, P.J.; Berends, L.; van der Velpen, V.; Jennings, A.; Haag, L.; Chandra, P.; Kay, C.D.; Rimm, E.B.; Cassidy, A. Blueberry anthocyanin intake attenuates the postprandial cardiometabolic effect of an energy-dense food challenge: Results from a double-blind, randomized controlled trial in metabolic syndrome participants. Clin. Nutr. 2022, 41, 165–175. [Google Scholar] [CrossRef]
- Yuan, L.; Muli, S.; Huybrechts, I.; Nöthlings, U.; Ahrens, W.; Scalbert, A.; Floegel, A. Assessment of fruit and vegetables intake with biomarkers in children and adolescents and their level of validation: A systematic review. Metabolites 2022, 12, 126. [Google Scholar] [CrossRef]
- Woodside, J.V.; Draper, J.; Lloyd, A.; McKinley, M.C. Use of biomarkers to assess fruit and vegetable intake. Proc. Nutr. Soc. 2017, 76, 308–315. [Google Scholar] [CrossRef] [Green Version]
- Pérez Rodrigo, C.; Aranceta, J.; Salvador, G.; Varela-Moreiras, G. Food frequency questionnaires. Nutr. Hosp. 2015, 31, 49–56. [Google Scholar]
- Salvador Castell, G.; Serra-Majem, L.; Ribas-Barba, L. What and how much do we eat? 24-hour dietary recall method. Nutr. Hosp. 2015, 31, 46–48. [Google Scholar]
- Penczynski, K.J.; Krupp, D.; Bring, A.; Bolzenius, K.; Remer, T. Relative validation of 24-h urinary hippuric acid excretion as a biomarker for dietary flavonoid intake from fruit and vegetables in healthy adolescents. Eur. J. Nutr. 2017, 56, 757–766. [Google Scholar] [CrossRef]
- Clarke, E.D.; Rollo, M.E.; Collins, C.E.; Wood, L.; Callister, R.; Philo, M.; Kroon, P.A.; Haslam, R.L. The relationship between dietary polyphenol intakes and urinary polyphenol concentrations in adults prescribed a high vegetable and fruit diet. Nutrients 2020, 12, 3431. [Google Scholar] [CrossRef]
- Krupp, D.; Doberstein, N.; Shi, L.; Remer, T. Hippuric acid in 24-hour urine collections is a potential biomarker for fruit and vegetable consumption in healthy children and adolescents. J. Nutr. 2012, 142, 1314–1320. [Google Scholar] [CrossRef] [Green Version]
- Rafiq, T.; Azab, S.M.; Anand, S.S.; Thabane, L.; Shanmuganathan, M.; Morrison, K.M.; Atkinson, S.A.; Stearns, J.C.; Teo, K.K.; Britz-McKibbin, P.; et al. Sources of variation in food-related metabolites during pregnancy. Nutrients 2022, 14, 2503. [Google Scholar] [CrossRef]
- Krupp, D.; Shi, L.; Egert, S.; Wudy, S.A.; Remer, T. Prospective relevance of fruit and vegetable consumption and salt intake during adolescence for blood pressure in young adulthood. Eur. J. Nutr. 2015, 54, 1269–1279. [Google Scholar] [CrossRef]
- Guerra, A.; Folesani, G.; Mena, P.; Ticinesi, A.; Allegri, F.; Nouvenne, A.; Pinelli, S.; Del Rio, D.; Borghi, L.; Meschi, T. Hippuric acid in 24 h urine collections as a biomarker of fruits and vegetables intake in kidney stone formers. Int. J. Food Sci. Nutr. 2014, 65, 1033–1038. [Google Scholar] [CrossRef] [PubMed]
- Taylor, E.N.; Fung, T.T.; Curhan, G.C. DASH-style diet associates with reduced risk for kidney stones. J. Am. Soc. Nephrol. 2009, 20, 2253–2259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorensen, M.D.; Hsi, R.S.; Chi, T.; Shara, N.; Wactawski-Wende, J.; Kahn, A.J.; Wang, H.; Hou, L.; Stoller, M.L.; Women’s Health Initiative Writing Group. Dietary intake of fiber, fruit and vegetables decreases the risk of incident kidney stones in women: A Women’s Health Initiative report. J. Urol. 2014, 192, 1694–1699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meschi, T.; Maggiore, U.; Fiaccadori, E.; Schianchi, T.; Bosi, S.; Adorni, G.; Ridolo, E.; Guerra, A.; Allegri, F.; Novarini, A.; et al. The effect of fruits and vegetables on urinary stone risk factors. Kidney Int. 2004, 66, 2402–2410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerra, A.; Ticinesi, A.; Allegri, F.; Nouvenne, A.; Prati, B.; Pinelli, S.; Merli, R.; Tana, C.; Lauretani, F.; Aloe, R.; et al. Insights about urinary hippuric and citric acid as biomarkers of fruit and vegetable intake in patients with kidney stones: The role of age and sex. Nutrition 2019, 59, 83–89. [Google Scholar] [CrossRef]
- Penczynski, K.J.; Remer, T.; Herder, C.; Kalhoff, H.; Rienks, J.; Markgraf, D.F.; Roden, M.; Buyken, A.E. Habitual flavonoid intake from fruit and vegetables during adolescence and serum lipid levels in early adulthood: A prospective analysis. Nutrients 2018, 10, 488. [Google Scholar] [CrossRef] [Green Version]
- Penczynski, K.J.; Herder, C.; Krupp, D.; Rienks, J.; Egert, S.; Wudy, S.A.; Roden, M.; Remer, T.; Buyken, A.E. Flavonoid intake from fruit and vegetables during adolescence is prospectively associated with a favourable risk factor profile for type 2 diabetes in early adulthood. Eur. J. Nutr. 2019, 58, 1159–1172. [Google Scholar] [CrossRef] [PubMed]
- Calvani, R.; Miccheli, A.; Capuani, G.; Tomassini Miccheli, A.; Puccetti, C.; Delfini, M.; Iaconelli, A.; Nanni, G.; Mingrone, G. Gut microbiome-derived metabolites characterize a peculiar obese urinary phenotype. Int. J. Obes. 2010, 34, 1095–1098. [Google Scholar] [CrossRef] [Green Version]
- Pallister, T.; Jackson, M.A.; Martin, T.C.; Galstonbury, C.A.; Jennings, A.; Beaumont, M.; Mohney, R.P.; Small, K.S.; MacGregor, A.; Steves, C.J.; et al. Untangling the relationship between diet and visceral fat mass through blood metabolomics and gut microbiome profiling. Int. J. Obes. 2017, 41, 1106–1113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, L.; López Sánchez, G.F.; Veronese, N.; Soysal, P.; Oh, H.; Barnett, Y.; Keyes, H.; Butler, L.; Allen, P.; Kostev, K.; et al. Fruit and vegetable intake and non-communicable diseases among adults aged ≥50 years in low- and middle-income countries. J. Nutr. Health Aging 2022, 26, 1003–1009. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.E.; Donkin, A.J.; Morgan, K.; Neale, R.J.; Pagf, R.M.; Silburn, R.L. Fruit and vegetable consumption in later life. Age Ageing 1998, 27, 723–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Guardia, L.; Yubero-Serrano, E.M.; Delgado-Lista, J.; Perez-Martinez, P.; Garcia-Rios, A.; Marin, C.; Camargo, A.; Delgado-Casado, N.; Roche, H.M.; Perez-Jimenez, F.; et al. Effects of the Mediterranean diet supplemented with Coenzyme Q10 on metabolomic profiles in elderly men and women. J. Gerontol. A Biol. Sci. 2015, 70, 78–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alkhady, A.; Edwards, C.A.; Combet, E. The urinary phenolic acid profile varies between younger and older adults after a polyphenol-rich meal despite limited differences in in vitro colonic metabolism. Eur. J. Nutr. 2019, 58, 1095–1111. [Google Scholar] [CrossRef] [Green Version]
- Tennoune, N.; Ventura, G.; Le Plenier, S.; Choisy, C.; Neveux, N.; Nakib, S.; Sarfati, G.; Raynaud-Simon, A.; Cynober, L.; De Bandt, J.P. Effect of aging on the availability of amino acids from an immune-enhancing diet (IED) after a surgical stress in rats. Clin. Nutr. 2020, 39, 2793–2801. [Google Scholar] [CrossRef]
- Hommos, M.S.; Glassock, R.J.; Rule, A.D. Structural and functional changes in human kidneys with healthy aging. J. Am. Soc. Nephrol. 2017, 28, 2838–2844. [Google Scholar] [CrossRef] [Green Version]
- Eriksen, B.O.; Palsson, R.; Ebert, N.; Melsom, T.; van der Giet, M.; Gudnason, V.; Indridason, O.S.; Inker, L.A.; Jenssen, T.G.; Levey, A.S.; et al. GFR in healthy aging: An individual participant data meta-analysis of iohexol clearance in European population-based cohorts. J. Am. Soc. Nephrol. 2020, 31, 1602–1615. [Google Scholar] [CrossRef] [PubMed]
- Aucella, F.; Corsonello, A.; Leosco, D.; Brunori, G.; Gesualdo, L.; Antonelli-Incalzi, R. Beyond chronic kidney disease: The diagnosis of Renal Disease in the Elderly as an unmet need. A position paper endorsed by Italian Society of Nephrology (SIN) and Italian Society of Geriatrics and Gerontology (SIGG). J. Nephrol. 2019, 32, 165–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soraci, L.; Corica, F.; Corsonello, A.; Remelli, F.; Abete, P.; Bellelli, G.; Bo, M.; Cherubini, A.; Di Bari, M.; Maggio, M.; et al. Prognostic interplay of kidney function with sarcopenia, anemia, disability and cognitive impairment. The GLISTEN study. Eur. J. Intern. Med. 2021, 93, 57–63. [Google Scholar] [CrossRef] [PubMed]
- De Favari Signini, E.; Castro, A.; Rehder-Santos, P.; Millan-Mattos, J.C.; Magalhães de Oliveira, J.; Minatel, V.; Falasco Pantoni, C.B.; Sobreiro Selistre de Araújo, H.; Fabbrizzi, F.; Porta, A.; et al. Integrative perspective of the healthy aging process considering the metabolome, cardiac autonomic modulation and cardiorespiratory fitness evaluated in age groups. Sci. Rep. 2022, 12, 21314. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Zampino, M.; Moaddel, R.; Chen, T.K.; Tian, Q.; Ferrucci, L.; Semba, R.D. Plasma metabolites associated with chronic kidney disease and renal function in adults from the Baltimore Longitudinal Study of Aging. Metabolomics 2021, 17, 9. [Google Scholar] [CrossRef]
- Pignanelli, M.; Just, C.; Bogiatzi, C.; Dinculescu, V.; Gloor, G.B.; Allen-Vercoe, E.; Reid, G.; Urquart, B.L.; Ruetz, K.N.; Velenosi, T.J.; et al. Mediterranean Diet Score: Associations with metabolic products of the intestinal microbiome, carotid plaque burden, and renal function. Nutrients 2018, 10, 779. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, H.; Miyamoto, Y.; Otagiri, M.; Maruyama, T. Update on the pharmacokinetics and redox properties of protein-bound uremic toxins. J. Pharm. Sci. 2011, 100, 3682–3695. [Google Scholar] [CrossRef]
- Schoots, A.C.; De Vries, P.M.; Thiemann, R.; Hazejager, W.A.; Visser, S.L.; Oe, P.L. Biochemical and neurophysiological parameters in hemodialyzed patients with chronic renal failure. Clin. Chim. Acta 1989, 185, 91–107. [Google Scholar]
- Lin, Y.T.; Wu, P.H.; Lee, H.H.; Mubanga, M.; Chen, C.S.; Kuo, M.C.; Chiu, Y.W.; Luo, P.L.; Hwang, S.J. Indole-3 acetic acid increased risk of impaired cognitive function in patients receiving hemodialysis. Neurotoxicology 2019, 73, 85–91. [Google Scholar] [CrossRef]
- Te Linde, E.; van Rooij, C.J.M.; Meijers, B.K.I.; De Loor, H.; Kessels, R.P.C.; Wetzels, J.F.M. Cognitive function and uremic toxins after kidney transplantation: An exploratory study. Kidney360 2020, 1, 1398–1406. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, Y.; Zeng, L.; Chen, G.; Zhang, L.; Liu, M.; Sheng, H.; Hu, X.; Su, J.; Zhang, D.; et al. The role fo gut microbiota and microbiota-related serum metabolites in the progression of diabetic kidney disease. Front. Pharmacol. 2021, 12, 757508. [Google Scholar] [CrossRef]
- Pignanelli, M.; Bogiatzi, C.; Gloor, G.; Allen-Vercoe, E.; Reid, G.; Urquhart, B.L.; Ruetz, K.N.; Velenosi, T.J.; Spence, J.D. Moderate renal impairment and toxic metabolites produced by the intestinal microbiome: Dietary implications. J. Ren. Nutr. 2019, 29, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Mangiola, F.; Nicoletti, A.; Gasbarrini, A.; Ponziani, F.R. Gut microbiota and aging. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 7404–7413. [Google Scholar]
- O’Toole, P.W.; Jeffery, I.B. Gut microbiota and aging. Science 2015, 350, 1214–1215. [Google Scholar] [CrossRef] [PubMed]
- Vaiserman, A.M.; Koliada, A.K.; Marotta, F. Gut microbiota: A player in aging and a target for anti-aging intervention. Ageing Res. Rev. 2017, 35, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Ticinesi, A.; Nouvenne, A.; Cerundolo, N.; Catania, P.; Prati, B.; Tana, C.; Meschi, T. Gut microbiota, muscle mass and function in aging: A focus on physical frailty and sarcopenia. Nutrients 2019, 11, 1633. [Google Scholar] [CrossRef] [Green Version]
- Badal, V.D.; Vaccariello, E.D.; Murray, E.R.; Yu, K.E.; Knight, R.; Jeste, D.V.; Nguyen, T.T. The gut microbiome, aging and longevity: A systematic review. Nutrients 2020, 12, 3759. [Google Scholar] [CrossRef]
- Biagi, E.; Rampelli, S.; Turroni, S.; Quercia, S.; Candela, M.; Brigidi, P. The gut microbiota of centenarians: Signatures of longevity in the gut microbiota profile. Mech. Ageing Dev. 2017, 165, 180–184. [Google Scholar] [CrossRef]
- Santoro, A.; Ostan, R.; Candela, M.; Biagi, E.; Brigidi, P.; Capri, M.; Franceschi, C. Gut microbiota changes in the extreme decades of human life: A focus on centenarians. Cell. Mol. Life Sci. 2018, 75, 129–148. [Google Scholar] [CrossRef] [Green Version]
- Ticinesi, A.; Milani, C.; Lauretani, F.; Nouvenne, A.; Mancabelli, L.; Lugli, G.A.; Turroni, F.; Duranti, S.; Mangifesta, M.; Viappiani, A.; et al. Gut microbiota composition is associated with polypharmacy in elderly hospitalized patients. Sci. Rep. 2017, 7, 11102. [Google Scholar] [CrossRef] [Green Version]
- Strasser, B.; Wolters, M.; Weyh, C.; Krüger, K.; Ticinesi, A. The effects of lifestyle and diet on gut microbiota composition, inflammation and muscle performance in our aging society. Nutrients 2021, 13, 2045. [Google Scholar] [CrossRef]
- Haran, J.P.; Zeamer, A.; Ward, D.V.; Dutta, P.; Bucci, V.; McCormick, B.A. The nursing home older adult gut microbiome composition shows time-dependent dysbiosis and is influenced by medication exposures, age, environment, and frailty. J. Gerontol. A Biol. Sci. 2021, 76, 1930–1938. [Google Scholar] [CrossRef]
- Milani, C.; Ticinesi, A.; Gerritsen, J.; Nouvenne, A.; Lugli, G.A.; Mancabelli, L.; Turroni, F.; Duranti, S.; Mangifesta, M.; Viappiani, A.; et al. Gut microbiota composition and Clostridium difficile infection in hospitalized elderly individuals: A metagenomic study. Sci. Rep. 2016, 6, 25945. [Google Scholar] [CrossRef] [Green Version]
- Williams, H.R.T.; Cox, I.J.; Walker, D.G.; Cobbold, J.F.L.; Taylor-Robinson, S.D.; Marshall, S.E.; Orchard, T.R. Differences in gut microbial metabolism are responsible for reduced Hippurate synthesis in Crohn’s disease. BMC Gastroenterol. 2010, 10, 108. [Google Scholar] [CrossRef] [Green Version]
- Pallister, T.; Jackson, M.A.; Martin, T.C.; Zierer, J.; Jennings, A.; Mohney, R.P.; MacGregor, A.; Steves, C.J.; Cassidy, A.; Spector, T.D.; et al. Hippurate as a metabolomic marker of gut microbiome diversity: Modulation by diet and relationship to metabolic syndrome. Sci. Rep. 2017, 7, 13760. [Google Scholar] [CrossRef] [Green Version]
- Fung, T.T.; Struijk, E.A.; Rodriguez-Artalejo, F.; Willett, W.C.; Lopez-Garcia, E. Fruit and vegetable intake and risk of frailty in women 60 years old or older. Am. J. Clin. Nutr. 2020, 112, 1540–1546. [Google Scholar] [CrossRef] [PubMed]
- Kojima, G.; Iliffe, S.; Jivraj, S.; Walters, K. Fruit and vegetable consumption and incident prefrailty and frailty in community-dwelling older people: The English Longitudinal Study of Ageing. Nutrients 2020, 12, 3882. [Google Scholar] [CrossRef] [PubMed]
- García-Esquinas, E.; Rahi, B.; Peres, K.; Colpo, M.; Dartigues, J.F.; Bandinelli, S.; Feart, C.; Rodríguez-Artalejo, F. Consumption of fruit and vegetables and risk of frailty: A dose-response analysis of 3 prospective cohorts of community-dwelling older adults. Am. J. Clin. Nutr. 2016, 104, 132–142. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Jia, Y.; Dai, J.; Fu, H.; Wang, Y.; Yan, H.; Zhu, Y.; Nie, X. Association of fruit and vegetable intake and frailty among Chinese elders: A cross-sectional study in three cities. J. Nutr. Health Aging 2019, 23, 890–895. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lee, Y.; Kye, S.; Chung, Y.S.; Kim, K.M. Association of vegetables and fruits consumption with sarcopenia in older adults: The fourth Korea National Health and Nutrition Examination Survey. Age Ageing 2015, 44, 96–102. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, M.; Becker, W.; Cederholm, T.E.; Byberg, L. A posteriori dietary patterns in 71-year-old Swedish men and the prevalence of sarcopenia 16 years later. Br. J. Nutr. 2021, 128, 909–920. [Google Scholar] [CrossRef] [PubMed]
- Koyanagi, A.; Veronese, N.; Solmi, M.; Oh, H.; Shin, J.I.; Jacob, L.; Yang, L.; Haro, J.M.; Smith, L. Fruit and vegetable consumption and sarcopenia among older adults in low- and middle-income countries. Nutrients 2020, 12, 706. [Google Scholar] [CrossRef] [Green Version]
- Sotos-Prieto, M.; Struijk, E.A.; Fung, T.T.; Rodríguez-Artalejo, F.; Willett, W.C.; Hu, F.B.; Lopez-Garcia, E. Association between the quality of plant-based diets and risk of frailty. J. Cachexia Sarcopenia Muscle 2022, 13, 2854–2862. [Google Scholar] [CrossRef]
- Kiuchi, Y.; Doi, T.; Tsutsumimoto, K.; Nakakubo, S.; Kurita, S.; Nishimoto, K.; Makizako, H.; Shimada, H. Association between dietary diversity and sarcopenia in community-dwelling older adults. Nutrition 2023, 106, 111896. [Google Scholar] [CrossRef] [PubMed]
- Yeung, S.S.Y.; Zhu, Z.L.Y.; Chan, R.S.M.; Kwok, T.; Woo, J. Prospective analysis of fruit and vegetable variety on health outcomes in community-dwelling Chinese older adults. J. Nutr. Health Aging 2021, 25, 735–741. [Google Scholar] [CrossRef]
- Ghoreishy, S.M.; Asoudeh, F.; Jayedi, A.; Mohammadi, H. Fruit and vegetable intake and risk of frailty: A systematic review and dose-response meta-analysis. Ageing Res. Rev. 2021, 71, 101460. [Google Scholar] [CrossRef] [PubMed]
- Kojima, G.; Taniguchi, Y.; Urano, T. Fruit and vegetable consumption and incident frailty in older adults: A systematic review and meta-analysis. J. Frailty Aging 2022, 11, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Kojima, G.; Avgerinou, C.; Iliffe, S.; Jivraj, S.; Sekiguchi, K.; Walters, K. Fruit and vegetable consumption and frailty: A systematic review. J. Nutr. Health Aging 2018, 22, 1010–1017. [Google Scholar] [CrossRef] [Green Version]
- Ticinesi, A.; Lauretani, F.; Milani, C.; Nouvenne, A.; Tana, C.; Del Rio, D.; Maggio, M.; Ventura, M.; Meschi, T. Aging gut microbiota at the cross-road between nutrition, physical frailty, and sarcopenia: Is there a gut-muscle axis? Nutrients 2017, 9, 1303. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Cheung, W.H.; Li, J.; Chow, S.K.H.; Yu, J.; Wong, S.H.; Ip, M.; Sung, J.J.Y.; Wong, R.M.Y. Understanding the gut microbiota and sarcopenia: A systematic review. J. Cachexia Sarcopenia Muscle 2021, 12, 1393–1407. [Google Scholar] [CrossRef]
- Zhang, T.; Cheng, J.K.; Hu, Y.M. Gut microbiota as a promising therapeutic target for age-related sarcopenia. Ageing Res. Rev. 2022, 81, 101739. [Google Scholar] [CrossRef]
- Urpi-Sarda, M.; Andres-Lacueva, C.; Rabassa, M.; Ruggiero, C.; Zamora-Ros, R.; Bandinelli, S.; Ferrucci, L.; Cherubini, A. The relationship between urinary total polyphenols and the frailty phenotype in a community-dwelling older population: The InCHIANTI Study. J. Gerontol. A Biol. Sci. 2015, 70, 1141–1147. [Google Scholar] [CrossRef] [Green Version]
- Saoi, M.; Li, A.; McGlory, C.; Stokes, T.; von Allmen, M.T.; Phillips, S.M.; Britz-McKibbin, P. Metabolic perturbations from step reduction in older persons at risk for sarcopenia: Plasma biomarkers of abrupt changes in physical activity. Metabolites 2019, 9, 0134. [Google Scholar] [CrossRef] [Green Version]
- Brunelli, L.; Davin, A.; Sestito, G.; Mimmi, M.C.; De Simone, G.; Balducci, C.; Pansarasa, O.; Forloni, G.; Cereda, C.; Pastorelli, R.; et al. Plasmatic hippuric acid as a hallmark of frailty in a Italian cohort: The mediaton effect of fruit-vegetable intake. J. Gerontol. Ser. A 2021, 76, 2081–2089. [Google Scholar] [CrossRef]
- Kameda, M.; Teruya, T.; Yanagida, M.; Kondoh, H. Reduced uremic metabolites are prominent features of sarcopenia, distinct from antioxidative markers of frailty. Aging 2021, 13, 20915. [Google Scholar] [CrossRef]
- Harmsen, J.F.; van Weeghel, M.; Parsons, R.; Janssens, G.E.; Wefers, J.; van Moorsel, D.; Hansen, J.; Hoeks, J.; Hesselink, M.K.C.; Houtkooper, R.H.; et al. Divergent remodeling of the skeletal muscle metabolome over 24 h between young, healthy men and older, metabolically compromised men. Cell Rep. 2022, 41, 111786. [Google Scholar] [CrossRef]
- Douzi, W.; Bon, D.; Suikkanen, S.; Soukkio, P.; Boildieu, N.; Nenonen, A.; Hupli, M.; Kukkonen-Harjula, K.; Dugué, B. 1H NMR urinary metabolomic analysis in older adults after hip fracture surgery may provide valuable information for patient profiling—A preliminary investigation. Metabolites 2022, 12, 744. [Google Scholar] [CrossRef] [PubMed]
- Wyngaert, K.V.; Van Craenenbroeck, A.H.; Holvoet, E.; Calders, P.; Van Biesen, W.; Eloot, S. Composite uremic load and physical performance in hemodialysis patients: A cross-sectional study. Toxins 2020, 12, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, E.; Mori, T.; Mishima, E.; Suzuki, A.; Sugawara, S.; Kurasawa, N.; Saigusa, D.; Miura, D.; Morikawa-Ichinose, T.; Saito, R.; et al. Metabolic alterations by indoxyl sulfate in skeletal muscle induce uremic sarcopenia in chronic kidney disease. Sci. Rep. 2016, 6, 36618. [Google Scholar] [CrossRef] [Green Version]
- Boeing, H.; Bechthold, A.; Bub, A.; Ellinger, S.; Haller, D.; Kroke, A.; Leschik-Bonnet, E.; Müller, M.J.; Oberritter, H.; Schulze, M.; et al. Critical review: Vegetables and fruit in the prevention of chronic diseases. Eur. J. Nutr. 2012, 51, 637–663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Wang, J.; Cao, L.; Shi, M.; Liu, H.; Zhao, Y.; Xia, Y. Fruit and vegetable consumption and cognitive disorders in older adults: A meta-analysis of observational studies. Front. Nutr. 2022, 9, 871061. [Google Scholar] [CrossRef]
- Román, G.C.; Jackson, R.E.; Gadhia, R.; Román, N.; Reis, J. Mediterranean diet: The role of long-chain ω-3 fatty acids in fish; polyphenols in fruits, vegetables, cereals, coffee, tea, cacao and wine; probiotics and vitamins in prevention of stroke, age-related cognitive decline, and Alzheimer disease. Rev. Neurol. 2019, 175, 724–741. [Google Scholar] [CrossRef] [PubMed]
- Solfrizzi, V.; Custodero, C.; Lozupone, M.; Imbimbo, B.P.; Valiani, V.; Agosti, P.; Schilardi, A.; D’Introno, A.; La Montagna, M.; Calvani, M.; et al. Relationships of dietary patterns, foods, and micro- and macronutrients with Alzheimer’s disease and late-life cognitive disorders: A systematic review. J. Alzheimers Dis. 2017, 59, 815–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maggi, S.; Ticinesi, A.; Limongi, F.; Noale, M.; Ecarnot, F. The role of nutrition and the Mediterranean diet on the trajectories of cognitive decline. Exp. Gerontol. 2023, 173, 112110. [Google Scholar] [CrossRef]
- Ticinesi, A.; Tana, C.; Nouvenne, A.; Prati, B.; Lauretani, F.; Meschi, T. Gut microbiota, cognitive frailty and dementia in older individuals: A systematic review. Clin. Interv. Aging 2018, 13, 1497–1511. [Google Scholar] [CrossRef] [Green Version]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The microbiota-gut-brain axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef]
- Ticinesi, A.; Mancabelli, L.; Carnevali, L.; Nouvenne, A.; Meschi, T.; Del Rio, D.; Ventura, M.; Sgoifo, A.; Angelino, D. Interaction between diet and microbiota in the pathophysiology of Alzheimer’s disease: Focus on polyphenols and dietary fibers. J. Alzheimers Dis. 2022, 86, 961–982. [Google Scholar] [CrossRef]
- Yilmaz, A.; Ugur, Z.; Bisgin, H.; Akyol, S.; Bahado-Singh, R.; Wilson, G.; Imam, K.; Maddens, M.E.; Graham, S.F. Targeted metabolic profiling of urine highlights a potential biomarker panel for the diagnosis of Alzheimer’s disease and mild cognitive impairment: A pilot study. Metabolites 2020, 10, 357. [Google Scholar] [CrossRef]
- Bøhn, S.K.; Myhrstad, M.C.W.; Thoresen, M.; Erlund, I.; Vasstrand, A.K.; Marciuch, A.; Carlsen, M.H.; Bastani, N.E.; Engedal, K.; Flekkøy, K.M.; et al. Bilberry/red grape juice decreases plasma biomarkers of inflammation and tissue damage in aged men with subjective memory impairment—A randomized clinical trial. BMC Nutr. 2021, 7, 75. [Google Scholar] [CrossRef]
- Barfoot, K.L.; Istas, G.; Feliciano, R.P.; Lamport, D.J.; Riddell, P.; Rodriguez-Mateos, A.; Williams, C.M. Effects of daily consumption of wild blueberry on cognition and urinary metabolites in school-aged children: A pilot study. Eur. J. Nutr. 2021, 60, 4263–4278. [Google Scholar] [CrossRef]
- Rutledge, G.A.; Sandhu, A.K.; Miller, M.G.; Edirisinghe, I.; Burton-Freeman, B.B.; Shukitt-Hale, B. Blueberry phenolics are associated with cognitive enhancement in supplemented healthy older adults. Food Funct. 2021, 12, 107–118. [Google Scholar] [CrossRef]
- Rong, W.; Han, K.; Zhao, Z.; An, J.; Li, Q.; Bi, K. The protective effect of Xanthoceras sorbifolia bunge husks on cognitive disorder based on metabolomics and gut microbiota analysis. J. Ethnopharmacol. 2021, 279, 113094. [Google Scholar] [CrossRef]
- Cattaneo, A.; Cattane, N.; Galluzzi, S.; Provasi, S.; Lopizzo, N.; Festari, C.; Ferrari, C.; Guerra, U.P.; Paghera, B.; Muscio, C.; et al. Association of brail amyloidosis with pro-inflammatory bacterial taxa and peripheral inflammation markers in cognitively impaired elderly. Neurobiol. Aging 2017, 49, 60–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogt, N.M.; Kerby, R.L.; Dill-McFarland, K.A.; Harding, S.J.; Merluzzi, A.P.; Johnson, S.C.; Carlsson, C.M.; Asthana, S.; Zetterberg, H.; Blennow, K.; et al. Gut microbiome alterations in Alzheimer’s disease. Sci. Rep. 2017, 7, 13537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, Z.Q.; Shen, L.L.; Li, W.W.; Fu, X.; Zeng, F.; Gui, L.; Lu, Y.; Cai, M.; Zhu, C.; Tan, Y.L.; et al. Gut microbiota is altered in patients with Alzheimer’s disease. J. Alzheimers Dis. 2018, 63, 1337–1346. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Wu, L.; Peng, G.; Han, Y.; Tang, R.; Ge, J.; Zhang, L.; Jia, L.; Yue, S.; Zhou, K.; et al. Altered microbiomes distinguish Alzheimer’s disease from amnestic mild cognitive impairment and health in a Chinese cohort. Brain Behav. Immun. 2019, 80, 633–643. [Google Scholar] [CrossRef]
- Haran, J.P.; Bhattarai, S.K.; Foley, S.E.; Dutta, P.; Ward, D.V.; Bucci, V.; McCormick, B.A. Alzheimer’s disease microbiome is associated with dysregulation of the anti-inflammatory P-glycoprotein pathway. mBio 2019, 10, e00632-19. [Google Scholar] [CrossRef] [Green Version]
- Ling, Z.; Zhu, M.; Yan, X.; Cheng, Y.; Shao, L.; Liu, X.; Jiang, R.; Wu, S. Structural and functional dysbiosis of fecal microbiota in Chinese patients with Alzheimer’s disease. Front. Cell Dev. Biol. 2021, 8, 634069. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, Y.; Quan, M.; Zhao, H.; Jia, J. Gut microbiota changes and their correlation with cognitive and neuropsychiatric symptoms in Alzheimer’s disease. J. Alzheimers Dis. 2021, 81, 583–595. [Google Scholar] [CrossRef]
- Xi, J.; Ding, D.; Zhu, H.; Wang, R.; Su, F.; Wu, W.; Xiao, Z.; Linag, X.; Zhao, Q.; Hong, Z.; et al. Disturbed microbial ecology in Alzheimer’s disease: Evidence from the gut microbiota. BMC Microbiol. 2021, 21, 226. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.J.; Chen, C.C.; Liao, H.Y.; Wu, Y.W.; Liou, J.M.; Wu, M.S.; Kuo, C.H.; Lin, C.H. Alteration of gut microbial metabolites in the systemic circulation of patients with Parkinson’s disease. J. Park. Dis. 2022, 12, 1219–1230. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.H.; Tang, W.H.; Lu, Y.C.; Wang, C.P.; Hung, W.C.; Wu, C.C.; Tsai, I.T.; Chung, F.M.; Houng, J.Y.; Lan, W.C.; et al. Association between hippuric acid and left ventricular hypertrophy in maintenance hemodialysis patients. Clin. Chim. Acta 2018, 484, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Cason, C.A.; Dolan, K.T.; Sharma, G.; Tao, M.; Kulkarni, R.; Helenowski, I.B.; Doane, B.M.; Avram, M.J.; McDermott, M.M.; Chang, E.B.; et al. Plasma microbiome-modulated indole- and phenyl-derived metabolites associate with advanced atherosclerosis and postoperative outcomes. J. Vasc. Surg. 2018, 68, 1552–1562.e7. [Google Scholar] [CrossRef]
- Ho, K.J.; Ramirez, J.L.; Kulkarni, R.; Harris, K.G.; Helenowski, I.; Xiong, L.; Ozaki, C.K.; Grenon, S.M. Plasma gut microbe-derived metabolites associated with peripheral artery disease and major adverse cardiac events. Microorganisms 2022, 10, 2065. [Google Scholar] [CrossRef]
- Heffron, S.P.; Rockman, C.B.; Adelman, M.A.; Gianos, E.; Guo, Y.; Xu, J.F.; Berger, J.S. Greater frequency of fruit and vegetable consumption is associated with lower prevalence of peripheral artery disease. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 1234–1240. [Google Scholar] [CrossRef] [Green Version]
- Biscetti, F.; Nardella, E.; Cecchini, A.L.; Landolfi, R.; Flex, A. The role of the microbiota in the diabetic peripheral artery disease. Mediat. Inflamm. 2019, 2019, 4128682. [Google Scholar] [CrossRef] [Green Version]
- Brezmes, J.; Llambrich, M.; Cumeras, R.; Gumà, J. Urine NMR metabolomics for precision oncology in colorectal cancer. Int. J. Mol. Sci. 2022, 23, 11171. [Google Scholar] [CrossRef]
- Mallafré-Muro, C.; Llambrich, M.; Cumeras, R.; Pardo, A.; Brezmes, J.; Marco, S.; Gumà, J. Comprehensive volatilome and metabolome signatures of colorectal cancer in urine: A systematic review and meta-analysis. Cancers 2021, 13, 2534. [Google Scholar] [CrossRef]
- Catchpole, G.; Platzer, A.; Weikert, C.; Kempkensteffen, C.; Johannsen, M.; Krause, H.; Jung, K.; Miller, K.; Willmitzer, L.; Selbig, J.; et al. Metabolic profiling reveals key metabolic features of renal cell carcinoma. J. Cell. Mol. Med. 2011, 15, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Łuczykowski, K.; Warmuzińska, N.; Operacz, S.; Stryjak, I.; Bogusiewicz, J.; Jacyna, J.; Wawrzyniak, R.; Struck-Lewicka, W.; Markuszewski, M.J.; Bojko, B. Metabolic evaluation of urine from patients diagnosed with high grade (HG) bladder cancer by SPME-LC-MS method. Molecules 2021, 26, 2194. [Google Scholar] [CrossRef]
- Jacyna, J.; Kordalewska, M.; Artymowicz, M.; Markuszewski, M.; Matuszewski, M.; Markuszewski, M.J. Pre- and post-resection urine metabolic profiles of bladder cancer patients: Results of preliminary studies on time series metabolomics analysis. Cancers 2022, 14, 1210. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Pan, L.B.; Yu, H.; Han, P.; Fu, J.; Zhang, Z.W.; Hu, J.C.; Yang, X.Y.; Keranmu, A.; Zhang, H.J.; et al. Gut microbiota-derived metabolites in inflammatory diseases based on targeted metabolomics. Front. Pharmacol. 2022, 13, 919181. [Google Scholar] [CrossRef] [PubMed]




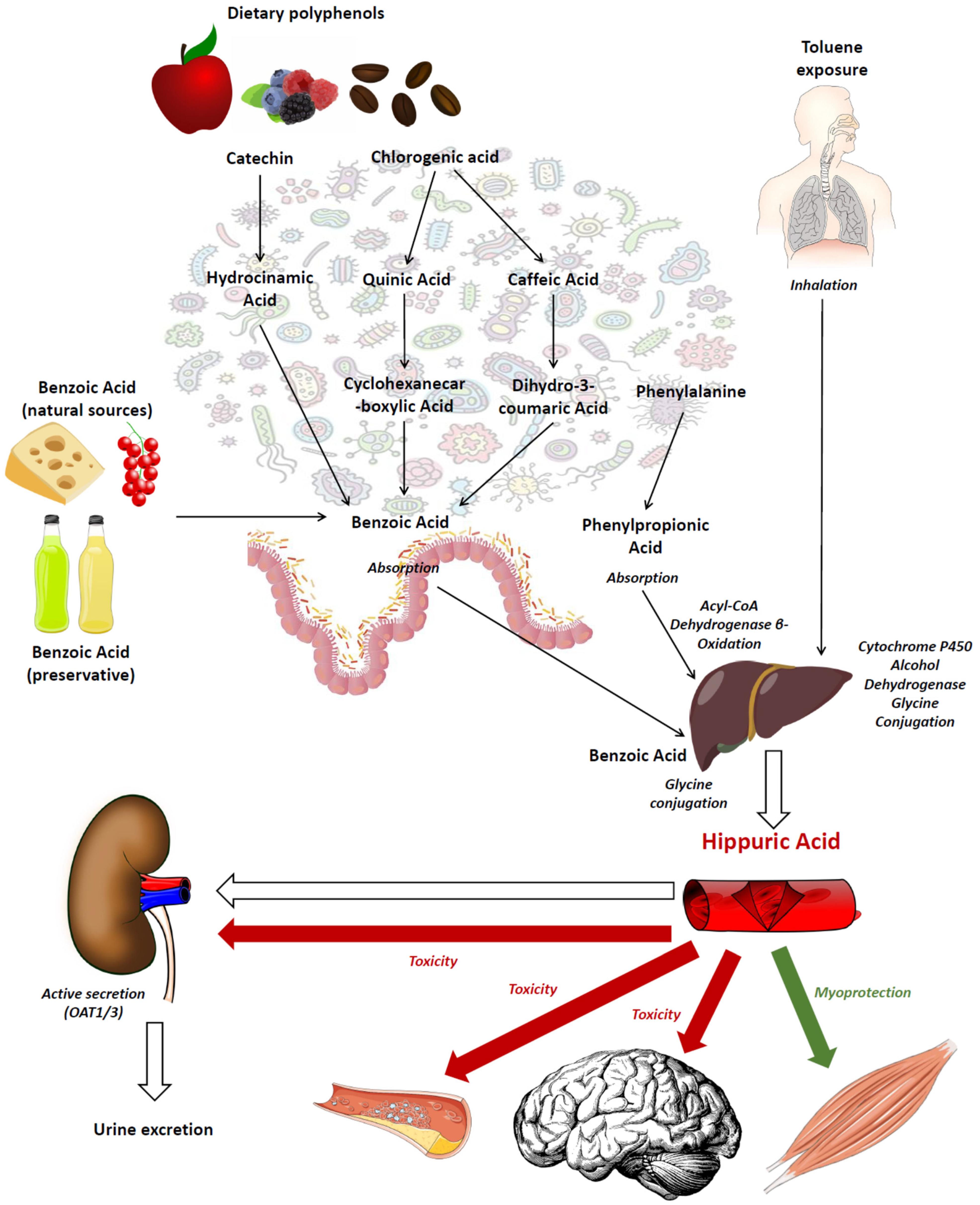{kind=link}
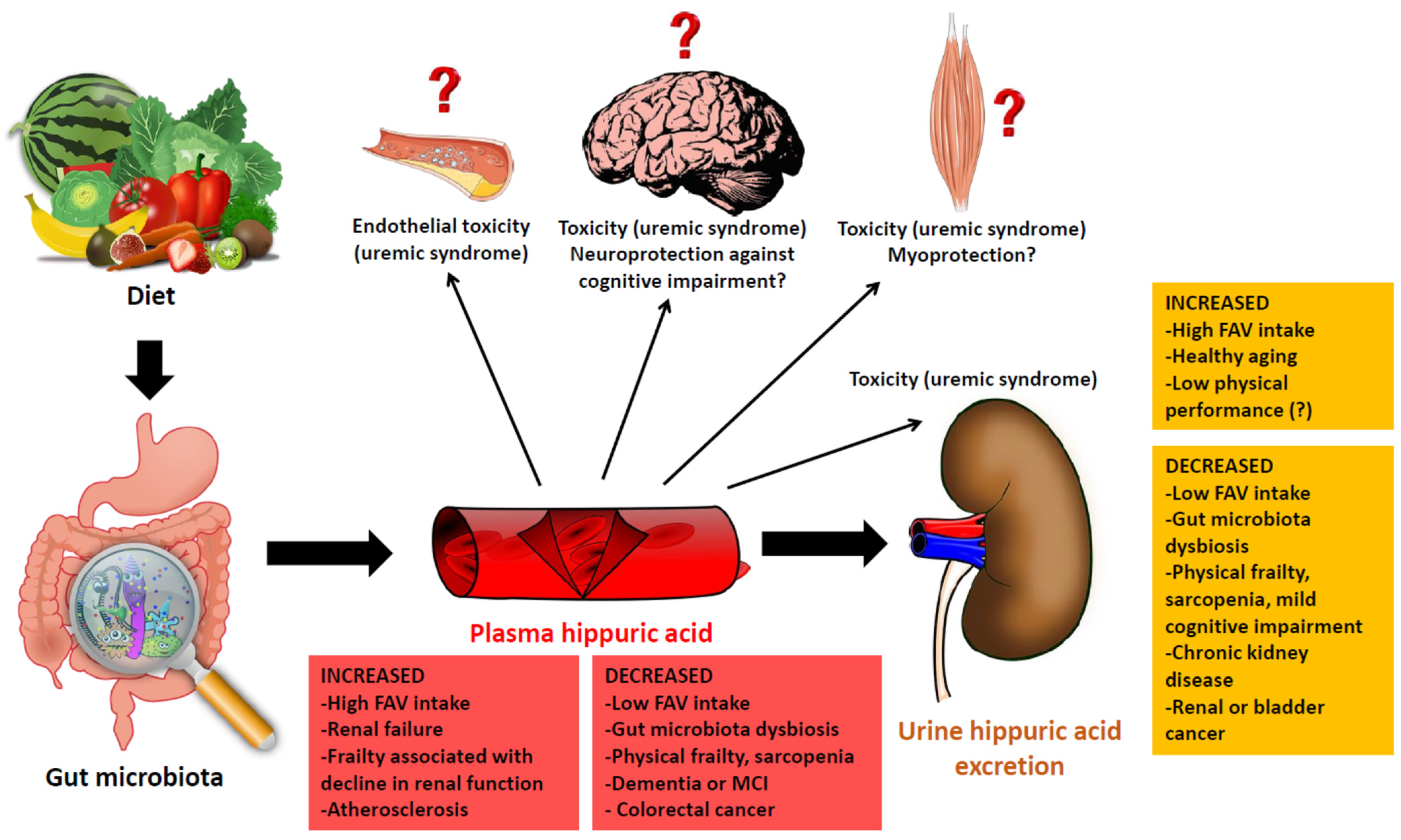{kind=link}
{kind=link}
| Author, Year (Ref) | Country | Study Design | Sample Size and Characteristics | Age | Method of Frailty Assessment | Key Findings |
|---|---|---|---|---|---|---|
| Saoi et al., 2019 [103] | Canada | Intervention (two weeks of inactivity with <1000 steps per day) | 17 overweight pre-diabetic older adults at risk for sarcopenia | 69 ± 17 | Not described | Physical inactivity was associated with indirect signs of muscle wasting and reduced plasma levels of uremic toxins, including HA. Resuming normal physical activity was not associated with recovery of baseline HA levels. |
| Brunelli et al., 2021 [104] | Italy | Longitudinal population-based | Profiling cohort: 65 fit and 65 frail Validation cohort: 124 fit, 59 pre-frail and 81 frail | Range 76–78 y.o. for all cohorts | Frailty Index based on 32 health variables or deficits | HA was the only marker, detected by an untargeted metabolomic approach, significantly lower in the plasma of the frail with respect to that of fit subjects and linearly associated with the FI. HA levels predicted incident frailty. |
| Kameda et al., 2021 [105] | Japan | Cross-sectional | 19 older community dwellers | 86 ± 7 | Skeletal Muscle Index measured by bioimpedance analysis; gait speed on 10 m straight walkway; grip strength | HA was among 22 plasma markers of sarcopenia with significantly lower levels in patients with low SMI detected by bioimpedance analysis, but was not associated with frailty. |
| Harmsen et al., 2022 [106] | Netherlands | Cross-sectional | 12 older metabolically compromised men 12 young fit adults | 65 ± 9 (older) 22 ± 2 (young) | Altered glucose metabolism | HA was significantly reduced in the plasma of older men and associated with a marker of altered skeletal muscle metabolism. The amplitude of plasma HA variations over the day was reduced in older men. |
| Douzi et al., 2022 [107] | Finland | RCT | 33 patients undergoing rehabilitation after hip fracture surgery | 80 ± 8 | Fried phenotypical criteria of frailty | Urinary HA was significantly increased in patients with low physical performance, measured by the SPPB score, and in patients who died during hospital stay. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ticinesi, A.; Guerra, A.; Nouvenne, A.; Meschi, T.; Maggi, S. Disentangling the Complexity of Nutrition, Frailty and Gut Microbial Pathways during Aging: A Focus on Hippuric Acid. Nutrients 2023, 15, 1138. https://doi.org/10.3390/nu15051138
Ticinesi A, Guerra A, Nouvenne A, Meschi T, Maggi S. Disentangling the Complexity of Nutrition, Frailty and Gut Microbial Pathways during Aging: A Focus on Hippuric Acid. Nutrients. 2023; 15(5):1138. https://doi.org/10.3390/nu15051138
Chicago/Turabian StyleTicinesi, Andrea, Angela Guerra, Antonio Nouvenne, Tiziana Meschi, and Stefania Maggi. 2023. "Disentangling the Complexity of Nutrition, Frailty and Gut Microbial Pathways during Aging: A Focus on Hippuric Acid" Nutrients 15, no. 5: 1138. https://doi.org/10.3390/nu15051138