
The Hepatic Antioxidant System Damage Induced with the Cafeteria (CAF) Diet Is Largely Counteracted Using SCD Probiotics during Development of Male Wistar Rats


Abstract
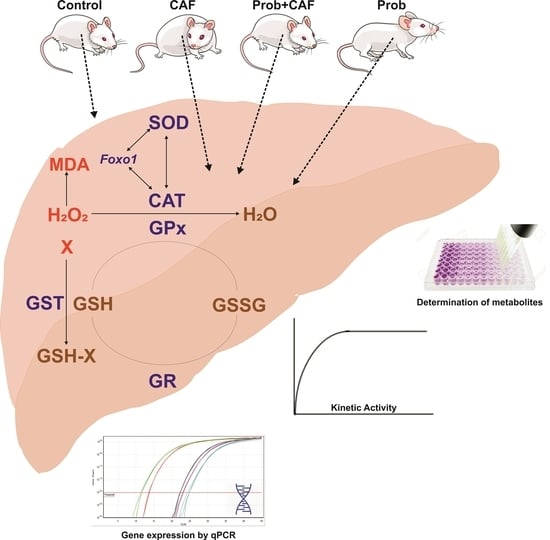
:
1. Introduction
2. Material and Methods
2.1. Experimental Design and Animal Care
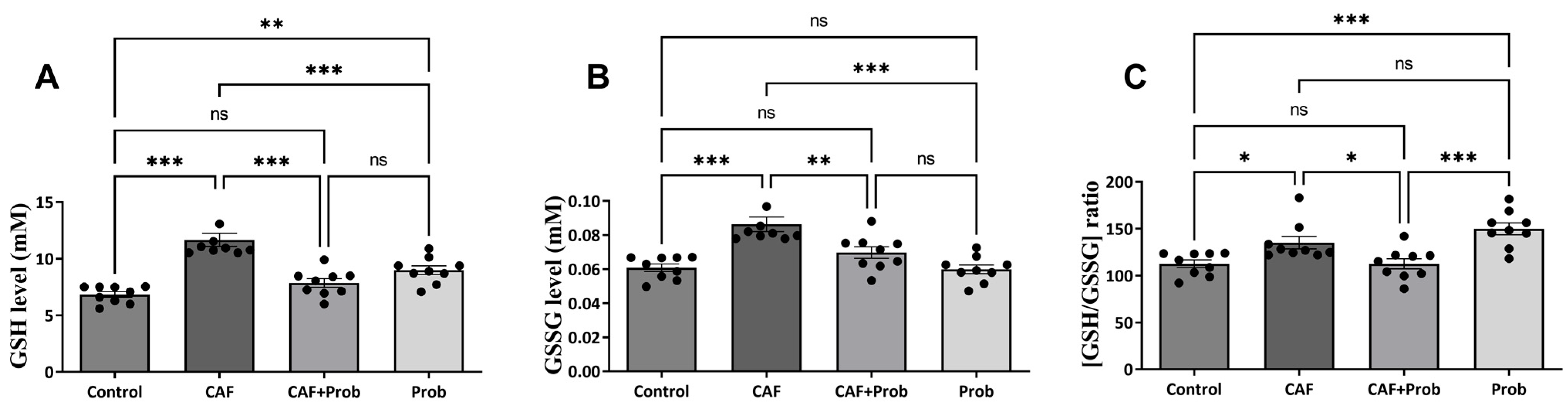
2.2. Determination of the Quantitative GSH Level and GSH/GSSG Ratio
2.3. Determination of Lipid Peroxidation
2.4. RNA Isolation and cDNA Synthesis
2.5. Primers and Gene Expression Analysis
2.6. Homogenate Preparation and Protein Determination Assay
2.7. Enzyme Activity Assays
2.8. Data Collection from Databases and the Validation of Obtained Data
2.9. Statistical Analysis
3. Results
3.1. Effects of Different Diet Combinations on Quantitative Hepatic Metabolite Levels
3.2. Effect of Diet Practices on Hepatic Antioxidant Gene Expressions
3.3. The Effect of Dietary Practices on the Enzyme Activities of Hepatic Antioxidant System
3.4. Correlation of Hepatic Antioxidant System Based on AWC
3.5. Identification of LIHC Prevalence Based on Obesity
3.6. Identification of LIHC Prevalence Based on Patient Weight
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- La Russa, D.; Giordano, F.; Marrone, A.; Parafati, M.; Janda, E.; Pellegrino, D. Oxidative Imbalance and Kidney Damage in Cafeteria Diet-Induced Rat Model of Metabolic Syndrome: Effect of Bergamot Polyphenolic Fraction. Antioxidants 2019, 8, 66. [Google Scholar] [CrossRef]
- Lalanza, J.F.; Snoeren, E.M.S. The cafeteria diet: A standardized protocol and its effects on behavior. Neurosci. Biobehav. Rev. 2021, 122, 92–119. [Google Scholar] [CrossRef] [PubMed]
- Buyukdere, Y.; Gulec, A.; Akyol, A. Cafeteria diet increased adiposity in comparison to high fat diet in young male rats. PeerJ 2019, 7, e6656. [Google Scholar] [CrossRef]
- Subias-Gusils, A.; Boque, N.; Caimari, A.; Del Bas, J.M.; Marine-Casado, R.; Solanas, M.; Escorihuela, R.M. A restricted cafeteria diet ameliorates biometric and metabolic profile in a rat diet-induced obesity model. Int. J. Food Sci. Nutr. 2021, 72, 767–780. [Google Scholar] [CrossRef]
- De Oliveira, F.A.; Costa, W.S.; FJ, B.S.; Gregorio, B.M. Resveratrol attenuates metabolic, sperm, and testicular changes in adult Wistar rats fed a diet rich in lipids and simple carbohydrates. Asian J. Androl. 2019, 21, 201–207. [Google Scholar] [CrossRef]
- Lang, P.; Hasselwander, S.; Li, H.; Xia, N. Effects of different diets used in diet-induced obesity models on insulin resistance and vascular dysfunction in C57BL/6 mice. Sci. Rep. 2019, 9, 19556. [Google Scholar] [CrossRef] [PubMed]
- Bortolin, R.; Vargas, A.; Gasparotto, J.; Chaves, P.; Schnorr, C.E.; Martinello, K.B.; Silveira, A.; Rabelo, T.K.; Gelain, D.; Moreira, J. A new animal diet based on human Western diet is a robust diet-induced obesity model: Comparison to high-fat and cafeteria diets in term of metabolic and gut microbiota disruption. Int. J. Obes. 2018, 42, 525–534. [Google Scholar] [CrossRef]
- Gual-Grau, A.; Guirro, M.; Mayneris-Perxachs, J.; Arola, L.; Boque, N. Impact of different hypercaloric diets on obesity features in rats: A metagenomics and metabolomics integrative approach. J. Nutr. Biochem. 2019, 71, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Crew, R.C.; Waddell, B.J.; Mark, P.J. Maternal obesity induced by a ‘cafeteria’ diet in the rat does not increase inflammation in maternal, placental or fetal tissues in late gestation. Placenta 2016, 39, 33–40. [Google Scholar] [CrossRef]
- Do Bonfim, T.H.F.; Tavares, R.L.; de Vasconcelos, M.H.A.; Gouveia, M.; Nunes, P.C.; Soares, N.L.; Alves, R.C.; de Carvalho, J.L.P.; Alves, A.F.; de Alencar Pereira, R. Potentially obesogenic diets alter metabolic and neurobehavioural parameters in Wistar rats: A comparison between two dietary models. J. Affect. Disord. 2021, 279, 451–461. [Google Scholar] [CrossRef]
- Gil-Cardoso, K.; Gines, I.; Pinent, M.; Ardevol, A.; Terra, X.; Blay, M. A cafeteria diet triggers intestinal inflammation and oxidative stress in obese rats. Br. J. Nutr. 2017, 117, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Beilharz, J.; Kaakoush, N.; Maniam, J.; Morris, M. Cafeteria diet and probiotic therapy: Cross talk among memory, neuroplasticity, serotonin receptors and gut microbiota in the rat. Mol. Psychiatry 2018, 23, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Rajoka, M.; Thirumdas, R.; Mehwish, H.; Umair, M.; Khurshid, M.; Hayat, H.; Phimolsiripol, Y.; Pallarés, N.; Martí-Quijal, F. Role of Food Antioxidants in Modulating Gut Microbial Com-Munities: Novel Understandings in Intestinal Oxidative Stress Damage and Their Impact on Host Health. Food Sci. Nutr. 2019, 59, 1927–1936. [Google Scholar]
- Alan, Y.; Savcı, A.; Koçpınar, E.F.; Ertaş, M. Postbiotic metabolites, antioxidant and anticancer activities of probiotic Leuconostoc pseudomesenteroides strains in natural pickles. Arch. Microbiol. 2022, 204, 571. [Google Scholar] [CrossRef] [PubMed]
- Budak, H.; Kocpinar, E.F.; Gonul, N.; Ceylan, H.; Erol, H.S.; Erdogan, O. Stimulation of gene expression and activity of antioxidant related enzyme in Sprague Dawley rat kidney induced by long-term iron toxicity. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2014, 166, 44–50. [Google Scholar] [CrossRef]
- Kocpinar, E.F.; Baltaci, N.G.; Akkemik, E.; Budak, H. Depletion of Tip60/Kat5 affects the hepatic antioxidant system in mice. J. Cell Biochem. 2023, 124, 103–117. [Google Scholar] [CrossRef]
- Ceylani, T. Effect of SCD probiotics supplemented with tauroursodeoxycholic acid (TUDCA) application on the aged rat gut microbiota composition. J. Appl. Microbiol. 2023, 134, lxad092. [Google Scholar] [CrossRef]
- Griffith, O.W. Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vinylpyridine. Anal. Biochem. 1980, 106, 207–212. [Google Scholar] [CrossRef]
- Sonmez Aydin, F.; Hukkamli, B.; Budak, H. Coaction of hepatic thioredoxin and glutathione systems in iron overload-induced oxidative stress. J. Biochem. Mol. Toxicol. 2021, 35, e22704. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Sun, Y.; Oberley, L.W.; Li, Y. A simple method for clinical assay of superoxide dismutase. Clin. Chem. 1988, 34, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Carlberg, I.; Mannervik, B. Glutathione reductase. Methods Enzymol. 1985, 113, 484–490. [Google Scholar] [CrossRef] [PubMed]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases. The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Tang, Z.; Kang, B.; Li, C.; Chen, T.; Zhang, Z. GEPIA2: An enhanced web server for large-scale expression profiling and interactive analysis. Nucleic Acids Res. 2019, 47, W556–W560. [Google Scholar] [CrossRef]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.; Varambally, S. UALCAN: A Portal for Facilitating Tumor Subgroup Gene Expression and Survival Analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef]
- Dequanter, D.; Dok, R.; Nuyts, S. Basal oxidative stress ratio of head and neck squamous cell carcinomas correlates with nodal metastatic spread in patients under therapy. OncoTargets Ther. 2017, 10, 259–263. [Google Scholar] [CrossRef]
- Alkazemi, D.; Rahman, A.; Habra, B. Alterations in glutathione redox homeostasis among adolescents with obesity and anemia. Sci. Rep. 2021, 11, 3034. [Google Scholar] [CrossRef]
- Wang, Y.; Yen, F.S.; Zhu, X.G.; Timson, R.C.; Weber, R.; Xing, C.; Liu, Y.; Allwein, B.; Luo, H.; Yeh, H.W.; et al. SLC25A39 is necessary for mitochondrial glutathione import in mammalian cells. Nature 2021, 599, 136–140. [Google Scholar] [CrossRef]
- Ceylan, H. Identification of hub genes associated with obesity-induced hepatocellular carcinoma risk based on integrated bioinformatics analysis. Med. Oncol. 2021, 38, 63. [Google Scholar] [CrossRef] [PubMed]
- Brahma, M.K.; Gilglioni, E.H.; Zhou, L.; Trépo, E.; Chen, P.; Gurzov, E.N. Oxidative stress in obesity-associated hepatocellular carcinoma: Sources, signaling and therapeutic challenges. Oncogene 2021, 40, 5155–5167. [Google Scholar] [CrossRef]
- Wang, S.; Song, Z.; Tan, B.; Zhang, J.; Zhang, J.; Liu, S. Identification and Validation of Hub Genes Associated With Hepatocellular Carcinoma Via Integrated Bioinformatics Analysis. Front. Oncol. 2021, 11, 614531. [Google Scholar] [CrossRef] [PubMed]
- Galasso, M.; Gambino, S.; Romanelli, M.G.; Donadelli, M.; Scupoli, M.T. Browsing the oldest antioxidant enzyme: Catalase and its multiple regulation in cancer. Free. Radic. Biol. Med. 2021, 172, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, Y.; Wang, S.; Li, Q.; Cao, B.; Huang, B.; Wang, T.; Guo, R.; Liu, N. SP1-induced lncRNA ZFPM2 antisense RNA 1 (ZFPM2-AS1) aggravates glioma progression via the miR-515-5p/Superoxide dismutase 2 (SOD2) axis. Bioengineered 2021, 12, 2299–2310. [Google Scholar] [CrossRef]
- El-Benhawy, S.A.; Morsi, M.I.; El-Tahan, R.A.; Matar, N.A.; Ehmaida, H.M. Radioprotective Effect of Thymoquinone in X-irradiated Rats. Asian Pac. J. Cancer Prev. APJCP 2021, 22, 3005–3015. [Google Scholar] [CrossRef]
- Umar Ijaz, M.; Rauf, A.; Mustafa, S.; Ahmed, H.; Ashraf, A.; Al-Ghanim, K.; Swamy Mruthinti, S.; Mahboob, S. Pachypodol attenuates Perfluorooctane sulphonate-induced testicular damage by reducing oxidative stress. Saudi J. Biol. Sci. 2022, 29, 1380–1385. [Google Scholar] [CrossRef]
- Borrelli, A.; Bonelli, P.; Tuccillo, F.M.; Goldfine, I.D.; Evans, J.L.; Buonaguro, F.M.; Mancini, A. Role of gut microbiota and oxidative stress in the progression of non-alcoholic fatty liver disease to hepatocarcinoma: Current and innovative therapeutic approaches. Redox Biol. 2018, 15, 467–479. [Google Scholar] [CrossRef] [PubMed]
- Mishra, V.; Shah, C.; Mokashe, N.; Chavan, R.; Yadav, H.; Prajapati, J. Probiotics as potential antioxidants: A systematic review. J. Agric. Food Chem. 2015, 63, 3615–3626. [Google Scholar] [CrossRef]
- Lee, J.Y.; Kang, C.H. Probiotics Alleviate Oxidative Stress in H2O2-Exposed Hepatocytes and t-BHP-Induced C57BL/6 Mice. Microorganisms 2022, 10, 234. [Google Scholar] [CrossRef] [PubMed]
- Teker, H.T.; Ceylani, T. Intermittent fasting supports the balance of the gut microbiota composition. Int. Microbiol. 2023, 26, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Ceylani, T.; Teker, H.T.; Samgane, G.; Gurbanov, R. Intermittent fasting-induced biomolecular modifications in rat tissues detected by ATR-FTIR spectroscopy and machine learning algorithms. Anal. Biochem. 2022, 654, 114825. [Google Scholar] [CrossRef]
- Ardahanli, I.; Ozkan, H.I.; Ozel, F.; Gurbanov, R.; Teker, H.T.; Ceylani, T. Infrared spectrochemical findings on intermittent fasting-associated gross molecular modifications in rat myocardium. Biophys. Chem. 2022, 289, 106873. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions, and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions, or products referred to in the content. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Onset | Week 1 | Week 2 | Week 3 | Week 4 | Week 5 | ||
|---|---|---|---|---|---|---|---|
| Energy (kcal) | Control | 1792 | 1803 | 2483 | 3247 | 3686 | 3476 |
| CAF | 1792 | 2354 | 3740 | 5848 | 6471 | 6312 | |
| Prob + CAF | 1792 | 3264 | 3669 | 5863 | 5110 | 6140 | |
| Prob | 1792 | 2093 | 3056 | 3629 | 3575 | 3285 | |
| Rodent nutrition (g) | Control | 67 | 67 | 93 | 121 | 138 | 130 |
| CAF | 67 | 61 | 89 | 136 | 134 | 123 | |
| Prob + CAF | 67 | 38 | 55 | 57 | 50 | 65 | |
| Prob | 67 | 41 | 53 | 58 | 39 | 47 | |
| AWC | Control | 79.9 ± 5.24 | 105.8 ± 5.61 | 134.8 ± 7.91 | 165.6 ± 7.87 | 196.7 ± 7.26 | 235.7 ± 6.98 |
| CAF | 79.3 ± 5.24 | 105.1 ± 5.43 | 128.6 ± 5.76 | 173.7 ± 5.96 | 207.8 ± 7.84 | 239.3 ± 9.36 | |
| Prob + CAF | 79.3 ± 5.69 | 106.1 ± 5.65 | 133.1 ± 6.69 | 172.7 ± 6.05 | 203.4 ± 7.06 | 227.3 ± 6.24 | |
| Prob | 79.7 ± 5.39 | 108.1 ± 5.26 | 130.6 ± 5.87 | 165.6 ± 7.51 | 203.7 ± 10.09 | 232.8 ± 8.78 |
| Gene Name | Accession Number | Elongation Position | Sequence (5′-3′) |
|---|---|---|---|
| Superoxide dismutase (Sod) | NM_017050.1 | Forward | GCTTCTGTCGTCTCCTTGCT |
| Reverse | CTCGAAGTGAATGACGCCCT | ||
| Catalase (Cat) | NM_012520.2 | Forward | GCGAATGGAGAGGCAGTGTA |
| Reverse | GTGCAAGTCTTCCTGCCTCT | ||
| Glutathione reductase (Gr) | NM_053906.2 | Forward | AGTTCACTGCTCCACACATCC |
| Reverse | TCCAGCTGAAAGAACCCATC | ||
| Glutathione peroxidase (Gpx) | NM_183403.2 | Forward | TGGCTTACATCGCCAAGTC |
| Reverse | CCGGGTAGTTGTTCCTCAGA | ||
| Glutathione S-transferase (Gst) | NM_017013.4 | Forward | AGACGGGAATTTGATGTTTGAC |
| Reverse | TGTCAATCAGGGCTCTCTCC | ||
| Forkhead box protein O1 (Foxo1) | NM_001285835.1 | Forward | ACCGTATCTGTGTGTGTGTGTG |
| Reverse | ACAGCCAAGTCCATCAAGAC | ||
| Glyceraldehyde-3-phosphate dehydrogenase (Gapdh) | NM_007393.5 | Forward | TGGACCTCATGGCCTACATG |
| Reverse | AGGGAGATGCTCAGTGTTGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aba, N.; Koçpınar, E.F.; Ceylani, T. The Hepatic Antioxidant System Damage Induced with the Cafeteria (CAF) Diet Is Largely Counteracted Using SCD Probiotics during Development of Male Wistar Rats. Nutrients 2023, 15, 4557. https://doi.org/10.3390/nu15214557
Aba N, Koçpınar EF, Ceylani T. The Hepatic Antioxidant System Damage Induced with the Cafeteria (CAF) Diet Is Largely Counteracted Using SCD Probiotics during Development of Male Wistar Rats. Nutrients. 2023; 15(21):4557. https://doi.org/10.3390/nu15214557
Chicago/Turabian StyleAba, Nurdan, Enver Fehim Koçpınar, and Taha Ceylani. 2023. "The Hepatic Antioxidant System Damage Induced with the Cafeteria (CAF) Diet Is Largely Counteracted Using SCD Probiotics during Development of Male Wistar Rats" Nutrients 15, no. 21: 4557. https://doi.org/10.3390/nu15214557