
Associating Inulin with a Pea Protein Improves Fast-Twitch Skeletal Muscle Mass and Muscle Mitochondrial Activities in Old Rats
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experiment
2.2. Body Composition Measurement
2.3. Protein Quality Evaluation
2.4. Plasma Analyses
2.5. Protein Synthesis Measurement
2.6. Quantitative RT-PCR Analysis
2.7. Western Blot Analysis
2.8. Mitochondrial Enzymatic Assays
2.9. Statistics
3. Results
3.1. Body Weight and Composition Changes and Final Relative Tissue Weights
3.2. Protein Quality Evaluation
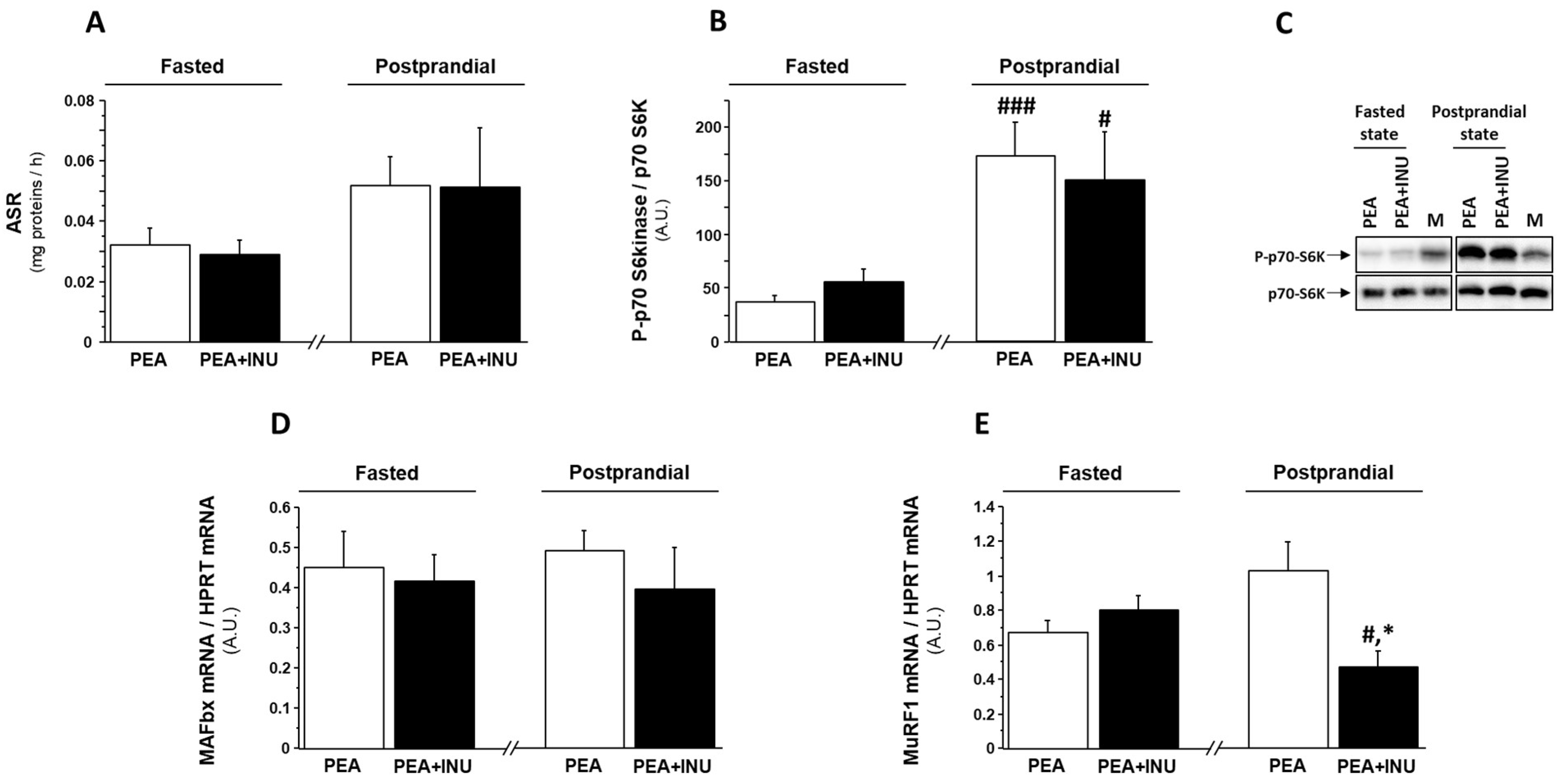
3.3. Muscle Protein Synthesis in Fasted and Postprandial State and Markers of Muscle Proteolysis
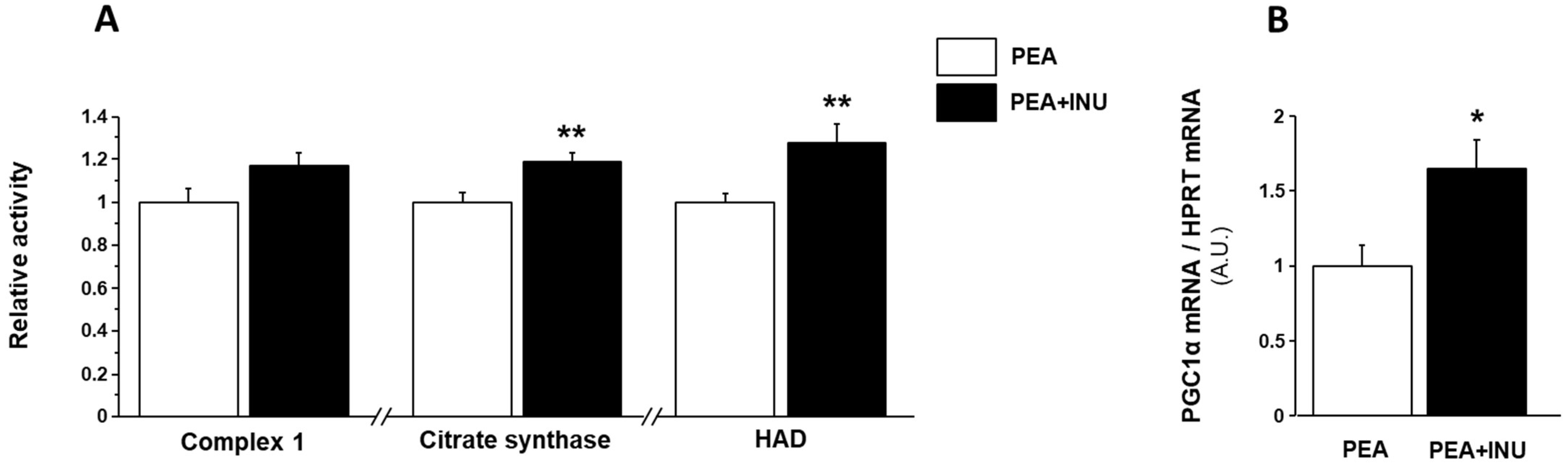
3.4. Muscle Mitochondrial Activity
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Floret, C.; Monnet, A.F.; Micard, V.; Walrand, S.; Michon, C. Replacement of animal proteins in food: How to take advantage of nutritional and gelling properties of alternative protein sources. Crit. Rev. Food Sci. Nutr. 2023, 63, 920–946. [Google Scholar] [CrossRef] [PubMed]
- FAO. The state of food and agriculture. In Livestock in the Balance; Food and Agriculture Organization of the United Nations: Rome, Italy, 2009; Available online: http://www.fao.org/3/a-i0680e.pdf (accessed on 1 May 2023).
- FAO. World agriculture: Towards 2030/2050. In ESA Working Paper No. 12-03, 160; Food and Agriculture Organization of the United Nations: Rome, Italy, 2012; Available online: http://www.fao.org/fileadmin/templates/esa/Global_persepctives/world_ag_2030_50_2012_rev.pdf (accessed on 1 May 2023).
- Berrazaga, I.; Micard, V.; Gueugneau, M.; Walrand, S. The Role of the Anabolic Properties of Plant-versus Animal-Based Protein Sources in Supporting Muscle Mass Maintenance: A Critical Review. Nutrients 2019, 11, 1825. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, L.; Panaite, S.A.; Bertazzo, A.; Visioli, F. Animal- and Plant-Based Protein Sources: A Scoping Review of Human Health Outcomes and Environmental Impact. Nutrients 2022, 14, 5115. [Google Scholar] [CrossRef] [PubMed]
- Walrand, S.; Boirie, Y. Optimizing protein intake in aging. Curr. Opin. Clin. Nutr. Metab. Care 2005, 8, 89–94. [Google Scholar] [CrossRef]
- Daily, J.W.; Park, S. Sarcopenia Is a Cause and Consequence of Metabolic Dysregulation in Aging Humans: Effects of Gut Dysbiosis, Glucose Dysregulation, Diet and Lifestyle. Cells 2022, 11, 338. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Larsson, S.C. Epidemiology of sarcopenia: Prevalence, risk factors, and consequences. Metabolism 2023, 144, 155533. [Google Scholar] [CrossRef]
- Berrazaga, I.; Salles, J.; Laleg, K.; Guillet, C.; Patrac, V.; Giraudet, C.; Le Bacquer, O.; Gueugneau, M.; Denis, P.; Pouyet, C.; et al. Anabolic Properties of Mixed Wheat-Legume Pasta Products in Old Rats: Impact on Whole-Body Protein Retention and Skeletal Muscle Protein Synthesis. Nutrients 2020, 12, 1596. [Google Scholar] [CrossRef]
- Marzani, B.; Balage, M.; Vénien, A.; Astruc, T.; Papet, I.; Dardevet, D.; Mosoni, L. Antioxidant supplementation restores defective leucine stimulation of protein synthesis in skeletal muscle from old rats. J. Nutr. 2008, 138, 2205–2211. [Google Scholar] [CrossRef]
- El Hajj, C.; Fares, S.; Chardigny, J.M.; Boirie, Y.; Walrand, S. Vitamin D supplementation and muscle strength in pre-sarcopenic elderly Lebanese people: A randomized controlled trial. Arch. Osteoporos. 2018, 14, 4. [Google Scholar] [CrossRef]
- Chanet, A.; Salles, J.; Guillet, C.; Giraudet, C.; Berry, A.; Patrac, V.; Domingues-Faria, C.; Tagliaferri, C.; Bouton, K.; Bertrand-Michel, J.; et al. Vitamin D supplementation restores the blunted muscle protein synthesis response in deficient old rats through an impact on ectopic fat deposition. J. Nutr. Biochem. 2017, 46, 30–38. [Google Scholar] [CrossRef]
- Walrand, S. Effect of vitamin D on skeletal muscle. Geriatr. Psychol. Neuropsychiatr. Vieil. 2016, 14, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, K.; Tsuchiya, A.; Nakano, O.; Kuroki, Y.; Oka, K.; Minemura, A.; Matsumoto, A.; Takahashi, M.; Kadota, Y.; Tochio, T.; et al. Increase in muscle mass associated with the prebiotic effects of 1-kestose in super-elderly patients with sarcopenia. Biosci. Microbiota Food Health 2021, 40, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Akram, W.; Garud, N.; Joshi, R. Role of inulin as prebiotics on inflammatory bowel disease. Drug Discov. Ther. 2019, 13, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Salles, J.; Guillet, C.; Le Bacquer, O.; Malnero-Fernandez, C.; Giraudet, C.; Patrac, V.; Berry, A.; Denis, P.; Pouyet, C.; Gueugneau, M.; et al. Pea Proteins Have Anabolic Effects Comparable to Milk Proteins on Whole Body Protein Retention and Muscle Protein Metabolism in Old Rats. Nutrients 2021, 13, 4234. [Google Scholar] [CrossRef] [PubMed]
- Anthony, J.C.; Reiter, A.K.; Anthony, T.G.; Crozier, S.J.; Lang, C.H.; MacLean, D.A.; Kimball, S.R.; Jefferson, L.S. Orally administered leucine enhances protein synthesis in skeletal muscle of diabetic rats in the absence of increases in 4E-BP1 or S6K1 phosphorylation. Diabetes 2002, 51, 928–936. [Google Scholar] [CrossRef]
- Balage, M.; Dupont, J.; Mothe-Satney, I.; Tesseraud, S.; Mosoni, L.; Dardevet, D. Leucine supplementation in rats induced a delay in muscle IR/PI3K signaling pathway associated with overall impaired glucose tolerance. J. Nutr. Biochem. 2011, 22, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Dijk, F.J.; van Dijk, M.; Walrand, S.; van Loon, L.J.C.; van Norren, K.; Luiking, Y.C. Differential effects of leucine and leucine-enriched whey protein on skeletal muscle protein synthesis in aged mice. Clin. Nutr. ESPEN 2018, 24, 127–133. [Google Scholar] [CrossRef]
- Dumas, A. [N-Determination According to Dumas]. Stickstoffbestimmung Nach Dumas. Die Praxis des Org. Chemikers, 41st ed.; Schrag: Nuremberg, Germany, 1962. [Google Scholar]
- Proll, J.; Petzke, K.J.; Ezeagu, I.E.; Metges, C.C. Low nutritional quality of unconventional tropical crop seeds in rats. J. Nutr. 1998, 128, 2014–2022. [Google Scholar] [CrossRef]
- Zangarelli, A.; Chanseaume, E.; Morio, B.; Brugère, C.; Mosoni, L.; Rousset, P.; Giraudet, C.; Patrac, V.; Gachon, P.; Boirie, Y.; et al. Synergistic effects of caloric restriction with maintained protein intake on skeletal muscle performance in 21-month-old rats: A mitochondria-mediated pathway. FASEB J. 2006, 20, 2439–2450. [Google Scholar] [CrossRef]
- Salles, J.; Chanet, A.; Berry, A.; Giraudet, C.; Patrac, V.; Domingues-Faria, C.; Rocher, C.; Guillet, C.; Denis, P.; Pouyet, C.; et al. Fast digestive, leucine-rich, soluble milk proteins improve muscle protein anabolism, and mitochondrial function in undernourished old rats. Mol. Nutr. Food Res. 2017, 61, 1700287. [Google Scholar] [CrossRef]
- Salles, J.; Cardinault, N.; Patrac, V.; Berry, A.; Giraudet, C.; Collin, M.-L.; Chanet, A.; Tagliaferri, C.; Denis, P.; Pouyet, C.; et al. Bee pollen improves muscle protein and energy metabolism in malnourished old rats through interfering with the Mtor signaling pathway and mitochondrial activity. Nutrients 2014, 6, 5500–5516. [Google Scholar] [CrossRef] [PubMed]
- Salles, J.; Chanet, A.; Guillet, C.; Vaes, A.M.; Brouwer-Brolsma, E.M.; Rocher, C.; Giraudet, C.; Patrac, V.; Meugnier, E.; Montaurier, C.; et al. Vitamin D status modulates mitochondrial oxidative capacities in skeletal muscle: Role in sarcopenia. Commun. Biol. 2022, 5, 1288. [Google Scholar] [CrossRef] [PubMed]
- Salles, J.; Chanet, A.; Giraudet, C.; Patrac, V.; Pierre, P.; Jourdan, M.; Luiking, Y.C.; Verlaan, S.; Migné, C.; Boirie, Y.; et al. 1, 25(OH)2-vitamin D3 enhances the stimulating effect of leucine and insulin on protein synthesis rate through Akt/PKB and mTOR mediated pathways in murine C2C12 skeletal myotubes. Mol. Nutr. Food Res. 2013, 57, 2137–2146. [Google Scholar] [CrossRef]
- Medja, F.; Allouche, S.; Frachon, P.; Jardel, C.; Malgat, M.; Mousson de Camaret, B.; Slama, A.; Lunardi, J.; Mazat, J.P.; Lombès, A. Development and implementation of standardized respiratory chain spectrophotometric assays for clinical diagnosis. Mitochondrion 2009, 9, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Cortés, N.G.; Pertuiset, C.; Dumon, E.; Börlin, M.; Hebert-Chatelain, E.; Pierron, D.; Feldmann, D.; Jonard, L.; Marlin, S.; Letellier, T.; et al. Novel mitochondrial DNA mutations responsible for maternally inherited nonsyndromic hearing loss. Hum. Mutat. 2012, 33, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Tardif, N.; Salles, J.; Guillet, C.; Tordjman, J.; Reggio, S.; Landrier, J.F.; Giraudet, C.; Patrac, V.; Bertrand-Michel, J.; Migne, C.; et al. Muscle ectopic fat deposition contributes to anabolic resistance in obese sarcopenic old rats through eIF2alpha activation. Aging Cell 2014, 13, 1001–1011. [Google Scholar] [CrossRef]
- Du, M.; Cheng, X.; Qian, L.; Huo, A.; Chen, J.; Sun, Y. Extraction, Physicochemical Properties, Functional Activities and Applications of Inulin Polysaccharide: A Review. Plant Foods Hum. Nutr. 2023, 78, 243–252. [Google Scholar] [CrossRef]
- Ikegami, S.; Tsuchihashi, F.; Harada, H.; Tsuchihashi, N.; Nishide, E.; Innami, S. Effect of viscous indigestible polysaccharides on pancreatic-biliary secretion and digestive organs in rats. J. Nutr. 1990, 120, 353–360. [Google Scholar] [CrossRef]
- Partridge, I.G.; Low, A.G.; Sambrook, I.E.; Corring, T. The influence of diet on the exocrine pancreatic secretion of growing pigs. Br. J. Nutr. 1982, 48, 137–145. [Google Scholar] [CrossRef]
- Cassidy, M.M.; Lightfoot, F.G.; Grau, L.E.; Story, J.A.; Kritchevsky, D.; Vahouny, G.V. Effect of chronic intake of dietary fibers on the ultrastructural topography of rat jejunum and colon: A scanning electron microscopy study. Am. J. Clin. Nutr. 1981, 34, 218–228. [Google Scholar] [CrossRef]
- Libao-Mercado, A.J.; Yin, Y.; van Eys, J.; de Lange, C.F. True ileal amino acid digestibility and endogenous ileal amino acid losses in growing pigs fed wheat shorts- or casein-based diets. J. Anim. Sci. 2006, 84, 1351–1361. [Google Scholar] [CrossRef] [PubMed]
- Bergner, H.; Simon, O.; Zimmer, M. [Influence of crude fibers in the diet of rats on the absorption of amino acids]. Einfluss des Gehaltes nativer Rohfaser in Diaten von Ratten auf die Aminosaurenresorption. Arch Tierernahr. 1975, 25, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.A.; Marcos, R.; Macarulla, M.T.; Larralde, J. Growth, hormonal status and protein turnover in rats fed on a diet containing peas (Pisum sativum L.) as the source of protein. Plant Foods Hum. Nutr. 1995, 47, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, H.; Nakanishi, N.; Miyoshi, T.; Okamura, T.; Hashimoto, Y.; Senmaru, T.; Majima, S.; Ushigome, E.; Asano, M.; Yamaguchi, M.; et al. Inulin reduces visceral adipose tissue mass and improves glucose tolerance through altering gut metabolites. Nutr. Metab. 2022, 19, 50. [Google Scholar] [CrossRef]
- Guillet, C.; Masgrau, A.; Walrand, S.; Boirie, Y. Impaired protein metabolism: Interlinks between obesity, insulin resistance and inflammation. Obes. Rev. 2012, 13 (Suppl. S2), 51–57. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Chen, D.; Yu, B.; Huang, Z.; Luo, Y.; Zheng, P.; Mao, X.; Yu, J.; Luo, J.; He, J. Effect of Dietary Inulin Supplementation on Growth Performance, Carcass Traits, and Meat Quality in Growing-Finishing Pigs. Animals 2019, 9, 840. [Google Scholar] [CrossRef] [PubMed]
- Buigues, C.; Fernández-Garrido, J.; Pruimboom, L.; Hoogland, A.J.; Navarro-Martínez, R.; Martínez-Martínez, M.; Verdejo, Y.; Carmen Mascarós, M.; Peris, C.; Cauli, O. Effect of a Prebiotic Formulation on Frailty Syndrome: A Randomized, Double-Blind Clinical Trial. Int. J. Mol. Sci. 2016, 17, 932. [Google Scholar] [CrossRef]
- Cani, P.D.; Possemiers, S.; Van de Wiele, T.; Guiot, Y.; Everard, A.; Rottier, O.; Geurts, L.; Naslain, D.; Neyrinck, A.; Lambert, D.M.; et al. Changes in gut microbiota control inflammation in obese mice through a mechanism involving GLP-2-driven improvement of gut permeability. Gut 2009, 58, 1091–1103. [Google Scholar] [CrossRef]
- Everard, A.; Lazarevic, V.; Derrien, M.; Girard, M.; Muccioli, G.G.; Neyrinck, A.M.; Possemiers, S.; Van Holle, A.; François, P.; de Vos, W.M.; et al. Responses of gut microbiota and glucose and lipid metabolism to prebiotics in genetic obese and diet-induced leptin-resistant mice. Diabetes 2011, 60, 2775–2786. [Google Scholar] [CrossRef]
- Giron, M.; Thomas, M.; Jarzaguet, M.; Mayeur, C.; Ferrere, G.; Noordine, M.L.; Bornes, S.; Dardevet, D.; Chassard, C.; Savary-Auzeloux, I. Lacticaseibacillus casei CNCM I-5663 supplementation maintained muscle mass in a model of frail rodents. Front. Nutr. 2022, 9, 928798. [Google Scholar] [CrossRef]
- Enck, P.; Zimmermann, K.; Rusch, K.; Schwiertz, A.; Klosterhalfen, S.; Frick, J.S. The effects of maturation on the colonic microflora in infancy and childhood. Gastroenterol. Res. Pract. 2009, 2009, 752401. [Google Scholar] [CrossRef] [PubMed]
- Morales, M.G.; Olguin, H.; Di Capua, G.; Brandan, E.; Simon, F.; Cabello-Verrugio, C. Endotoxin-induced skeletal muscle wasting is prevented by angiotensin-(1-7) through a p38 MAPK-dependent mechanism. Clin. Sci. 2015, 129, 461–476. [Google Scholar] [CrossRef] [PubMed]
- Picca, A.; Fanelli, F.; Calvani, R.; Mulè, G.; Pesce, V.; Sisto, A.; Pantanelli, C.; Bernabei, R.; Landi, F.; Marzetti, E. Gut Dysbiosis and Muscle Aging: Searching for Novel Targets against Sarcopenia. Mediat. Inflamm. 2018, 2018, 7026198. [Google Scholar] [CrossRef]
- Schaafsma, G.; Slavin, J.L. Significance of Inulin Fructans in the Human Diet. Compr. Rev. Food Sci. Food Saf. 2015, 14, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Byrne, T.; Cooke, J.; Bambrick, P.; McNeela, E.; Harrison, M. Circulating inflammatory biomarker responses in intervention trials in frail and sarcopenic older adults: A systematic review and meta-analysis. Exp. Gerontol. 2023, 177, 112199. [Google Scholar] [CrossRef]
- Macfarlane, S.; Cleary, S.; Bahrami, B.; Reynolds, N.; Macfarlane, G.T. Synbiotic consumption changes the metabolism and composition of the gut microbiota in older people and modifies inflammatory processes: A randomised, double-blind, placebo-controlled crossover study. Aliment Pharmacol. Ther. 2013, 38, 804–816. [Google Scholar] [CrossRef]
- Bellanti, F.; Lo Buglio, A.; Vendemiale, G. Hepatic Mitochondria-Gut Microbiota Interactions in Metabolism-Associated Fatty Liver Disease. Metabolites 2023, 13, 322. [Google Scholar] [CrossRef]
- Colangeli, L.; Escobar Marcillo, D.I.; Simonelli, V.; Iorio, E.; Rinaldi, T.; Sbraccia, P.; Fortini, P.; Guglielmi, V. The Crosstalk between Gut Microbiota and White Adipose Tissue Mitochondria in Obesity. Nutrients 2023, 15, 1723. [Google Scholar] [CrossRef]
- Righetto, I.; Gasparotto, M.; Casalino, L.; Vacca, M.; Filippini, F. Exogenous Players in Mitochondria-Related CNS Disorders: Viral Pathogens and Unbalanced Microbiota in the Gut-Brain Axis. Biomolecules 2023, 13, 169. [Google Scholar] [CrossRef]
- Gu, X.; Wang, W.; Yang, Y.; Lei, Y.; Liu, D.; Wang, X.; Wu, T. The Effect of Metabolites on Mitochondrial Functions in the Pathogenesis of Skeletal Muscle Aging. Clin. Interv. Aging 2022, 17, 1275–1295. [Google Scholar] [CrossRef]
- Murphy, E.A.; Velazquez, K.T.; Herbert, K.M. Influence of high-fat diet on gut microbiota: A driving force for chronic disease risk. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 515–520. [Google Scholar] [CrossRef] [PubMed]
- van Krimpen, S.J.; Jansen, F.A.C.; Ottenheim, V.L.; Belzer, C.; van der Ende, M.; van Norren, K. The Effects of Pro-, Pre-, and Synbiotics on Muscle Wasting, a Systematic Review-Gut Permeability as Potential Treatment Target. Nutrients 2021, 13, 1115. [Google Scholar] [CrossRef] [PubMed]
- Vrieze, A.; Van Nood, E.; Holleman, F.; Salojärvi, J.; Kootte, R.S.; Bartelsman, J.F.; Dallinga-Thie, G.M.; Ackermans, M.T.; Serlie, M.J.; Oozeer, R.; et al. Transfer of intestinal microbiota from lean donors increases insulin sensitivity in individuals with metabolic syndrome. Gastroenterology 2012, 143, 913–916.e7. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Chang, S.; Chang, H.; Wu, C.; Pan, C.; Chang, C.; Chan, C.; Huang, H. Probiotic supplementation attenuates age-related sarcopenia via the gut-muscle axis in SAMP8 mice. J. Cachexia Sarcopenia Muscle 2022, 13, 515–531. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, S.; Wang, J.; Liu, W.; Gong, H.; Zhang, Z.; Lyu, B.; Yu, H. Insoluble Dietary Fiber from Soybean Residue (Okara) Exerts Anti-Obesity Effects by Promoting Hepatic Mitochondrial Fatty Acid Oxidation. Foods 2023, 12, 2081. [Google Scholar] [CrossRef] [PubMed]
- Sridharan, G.V.; Choi, K.; Klemashevich, C.; Wu, C.; Prabakaran, D.; Pan, L.B.; Steinmeyer, S.; Mueller, C.; Yousofshahi, M.; Alaniz, R.C.; et al. Prediction and quantification of bioactive microbiota metabolites in the mouse gut. Nat. Commun. 2014, 5, 5492. [Google Scholar] [CrossRef]
- Kimura, I.; Inoue, D.; Hirano, K.; Tsujimoto, G. The SCFA Receptor GPR43 and Energy Metabolism. Front. Endocrinol. 2014, 5, 85. [Google Scholar] [CrossRef]
- Gao, Z.; Yin, J.; Zhang, J.; Ward, R.E.; Martin, R.J.; Lefevre, M.; Cefalu, W.T.; Ye, J. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes 2009, 58, 1509–1517. [Google Scholar] [CrossRef]
- Nucci, R.A.B.; Filho, V.A.N.; Jacob-Filho, W.; Otoch, J.P.; Pessoa, A.F.M. Role of Nutritional Supplements on Gut-Muscle Axis Across Age: A Mini-Review. Cell. Physiol. Biochem. 2023, 57, 161–168. [Google Scholar]
- Mao, B.; Li, D.; Zhao, J.; Liu, X.; Gu, Z.; Chen, Y.Q.; Zhang, H.; Chen, W. Metagenomic insights into the effects of fructo-oligosaccharides (FOS) on the composition of fecal microbiota in mice. J. Agric. Food Chem. 2015, 63, 856–863. [Google Scholar] [CrossRef]
- Miralles-Pérez, B.; Nogués, M.R.; Sánchez-Martos, V.; Fortuño-Mar, À.; Ramos-Romero, S.; Torres, J.L.; Ponomarenko, J.; Amézqueta, S.; Zhang, X.; Romeu, M. Influence of Dietary Inulin on Fecal Microbiota, Cardiometabolic Risk Factors, Eicosanoids, and Oxidative Stress in Rats Fed a High-Fat Diet. Foods 2022, 11, 4072. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| PEA | PEA + INU | |
|---|---|---|
| Diet composition (g/100 g) | ||
| Pea protein | 14 | 14 |
| Fat (soybean oil) | 6 | 6 |
| Carbohydrates | 68 | 68 |
| Cellulose | 7.5 | 0 |
| Inulin | 0 | 7.5 |
| Vitamin and mineral mix | 4.5 | 4.5 |
| Component | Final Concentration or Volume |
|---|---|
| Leucine | 16 g/L |
| Sucrose | 191 g/L |
| Glucose | 191 g/L |
| Glutamine | 0.3 g/L |
| Amino acid solution (M5550–Sigma Aldrich) | 2 mL |
| Distilled water | q.s. 50 mL |
| Gene Name | Forward and Reverse Primers |
|---|---|
| MAFbx Muscle atrophy F-box | For 5′-AGTGAAGACCGGCTACTGTGGAA-3′ Rev 5′-TTGCAAAGCTGCAGGGTGAC-3′ |
| MuRF1 Muscle RING finger-1 | For 5′-GTGAAGTTGCCCCCTTACAA-3′ Rev 5′-TGGAGATGCAATTGCTCAGT-3′ |
| PGC1α Peroxisome proliferator-activated receptor gamma coactivator 1-alpha | For 5′-AGTTTTTGGTGAAATTGAGGAAT-3′ Rev 5′-TCATACTTGCTCTTGGTGGAAGC-3′ |
| HPRT Hypoxanthine-guanine phosphoribosyltransferase | For 5′-AGTTGAGAGATCATCTCCAC-3′ Rev 5′-TTGCTGACCTGCTGGATTAC-3′ |
| PEA (n = 14) | PEA + INU (n = 12) | p | |
|---|---|---|---|
| Body weight (g) | |||
| initial | 598 ± 19 | 577 ± 18 | 0.4 |
| final | 612 ± 27 | 588 ± 21 | 0.5 |
| change | +13 ± 16 | +12 ± 12 | 0.9 |
| Fat mass (g) | |||
| initial | 108 ± 8 | 89 ± 8 | 0.1 |
| final | 140 ± 16 # | 115 ± 12 ## | 0.2 |
| change | +33 ± 12 | +26 ± 7 | 0.6 |
| Lean mass (g) | |||
| initial | 438 ± 12 | 436 ± 13 | 0.9 |
| final | 414 ± 13 ### | 418 ± 14 ## | 0.8 |
| change | −23 ± 4 | −18 ± 5 | 0.4 |
| PEA (n = 14) | PEA + INU (n = 12) | |
|---|---|---|
| Plantaris (mg/100 g body weight) | 46.0 ± 1.6 | 53.5 ± 3.4 * |
| Tibialis (mg/100 g body weight) | 70.6 ± 2.7 | 82.0 ± 4.1 * |
| Gastrocnemius (mg/100 g body weight) | 202.1 ± 9.0 | 238.9 ± 15.4 * |
| Soleus (mg/100 g body weight) | 29.8 ± 1.9 | 33.9 ± 2.9 |
| Perirenal adipose tissue (g/100 g body weight) | 3.0 ± 0.3 | 2.2 ± 0.2 |
| Subcutaneous adipose tissue (g/100 g body weight) | 2.3 ± 0.2 | 1.9 ± 0.3 |
| Liver (g/100 g body weight) | 2.3 ± 0.1 | 2.2 ± 0.1 |
| Heart (g/100 g body weight) | 0.3 ± 0.0 | 0.3 ± 0.0 |
| PEA (n = 14) | PEA + INU (n = 12) | |
|---|---|---|
| Nitrogen intake (g) | 1.65 ± 0.09 | 1.86 ± 0.11 |
| Fecal nitrogen (g) | 0.13 ± 0.01 | 0.18 ± 0.01 * |
| Urinary nitrogen (g) | 0.92 ± 0.07 | 0.86 ± 0.06 |
| FN/NI | 0.08 ± 0.01 | 0.10 ± 0.01 * |
| UN/NI | 0.56 ± 0.03 | 0.47 ± 0.03 |
| Nitrogen balance (g) | 0.60 ± 0.07 | 0.81 ± 0.09 |
| PEA | PEA + INU | |||
|---|---|---|---|---|
| Fasted State (n = 8) | Postprandial State (n = 6) | Fasted State (n = 7) | Postprandial State (n = 5) | |
| Glucose (g/L) | 0.970 ± 0.108 | 1.574 ± 0.072 *** | 0.795 ± 0.112 | 1.445 ± 0.083 *** |
| Insulin (ng/mL) | 0.553 ± 0.102 | 2.531 ± 0.456 *** | 0.564 ± 0.125 | 2.037 ± 1.066 * |
| Triglycerides (g/L) | 0.604 ± 0.176 | 0.745 ± 0.133 | 0.682 ± 0.139 | 0.486 ± 0.036 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salles, J.; Gueugneau, M.; Patrac, V.; Malnero-Fernandez, C.; Guillet, C.; Le Bacquer, O.; Giraudet, C.; Sanchez, P.; Collin, M.-L.; Hermet, J.; et al. Associating Inulin with a Pea Protein Improves Fast-Twitch Skeletal Muscle Mass and Muscle Mitochondrial Activities in Old Rats. Nutrients 2023, 15, 3766. https://doi.org/10.3390/nu15173766
Salles J, Gueugneau M, Patrac V, Malnero-Fernandez C, Guillet C, Le Bacquer O, Giraudet C, Sanchez P, Collin M-L, Hermet J, et al. Associating Inulin with a Pea Protein Improves Fast-Twitch Skeletal Muscle Mass and Muscle Mitochondrial Activities in Old Rats. Nutrients. 2023; 15(17):3766. https://doi.org/10.3390/nu15173766
Chicago/Turabian StyleSalles, Jérôme, Marine Gueugneau, Véronique Patrac, Carmen Malnero-Fernandez, Christelle Guillet, Olivier Le Bacquer, Christophe Giraudet, Phelipe Sanchez, Marie-Laure Collin, Julien Hermet, and et al. 2023. "Associating Inulin with a Pea Protein Improves Fast-Twitch Skeletal Muscle Mass and Muscle Mitochondrial Activities in Old Rats" Nutrients 15, no. 17: 3766. https://doi.org/10.3390/nu15173766