
Dried Plum’s Polyphenolic Compounds and Carbohydrates Contribute to Its Osteoprotective Effects and Exhibit Prebiotic Activity in Estrogen Deficient C57BL/6 Mice

 , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
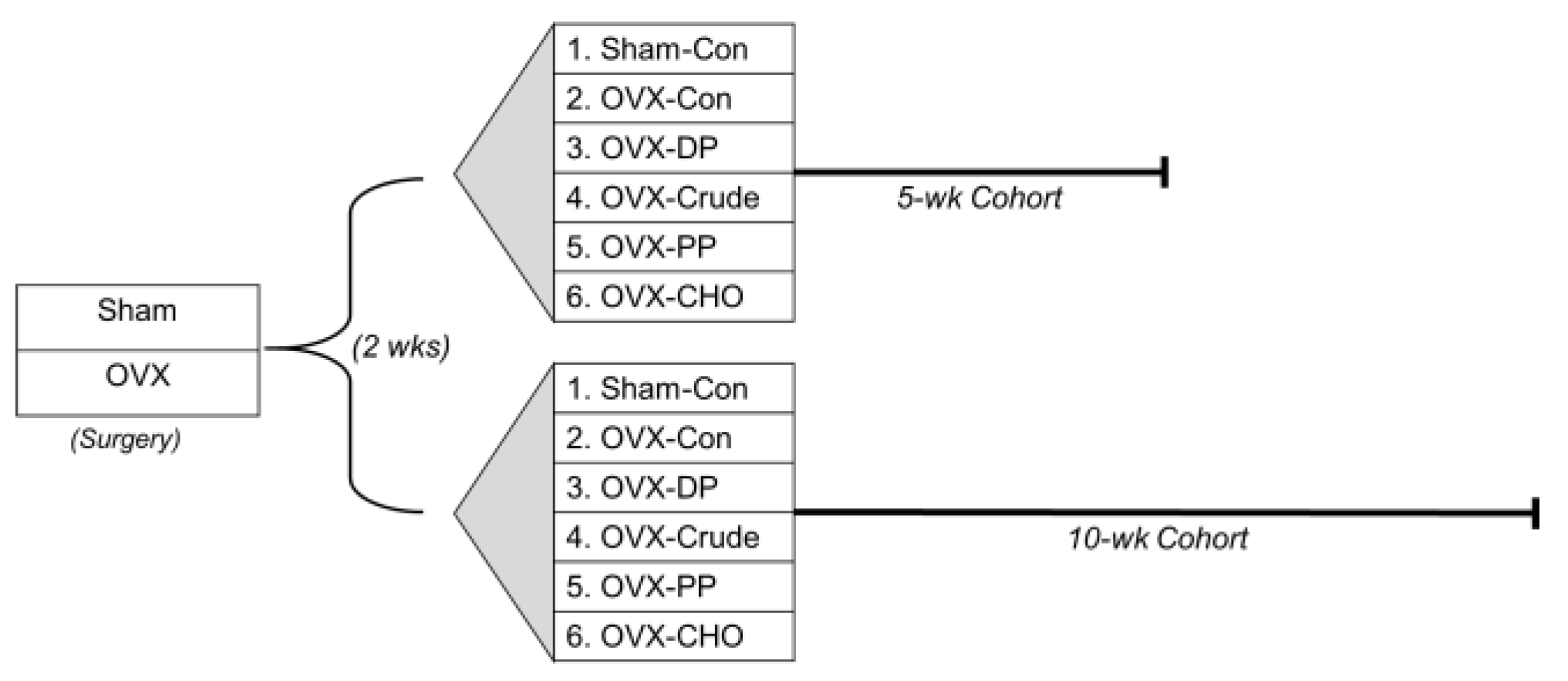
2.1. Animal Care and Experimental Design
2.2. Preparation of the Polyphenol and Carbohydrate Fractions
2.3. Micro-Computed Tomography (MicroCT) Analyses
2.4. Serum Bone Biomarkers
2.5. Short Chain Fatty Acids
2.6. Gut Microbiota Profiling
2.7. Statistical Analyses
3. Results
3.1. Dried Plum and Crude Extract Influence on Body Weight, Bone Density and Bone Biomarkers of Ovariectomized Mice
3.2. Dried Plum and Crude Extract Diets Improve Trabecular and Cortical Bone Microarchitecture of Ovariectomized Mice
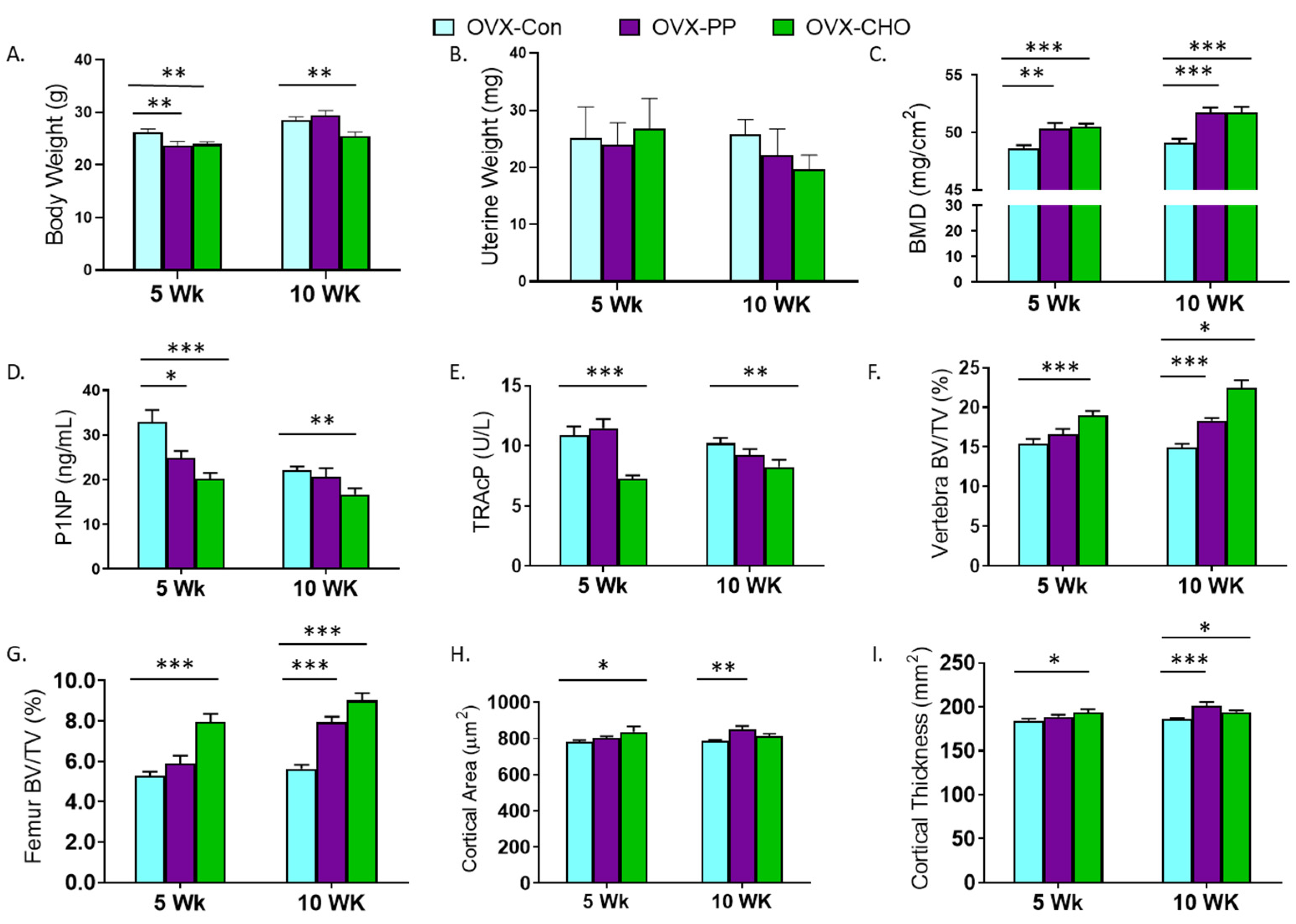
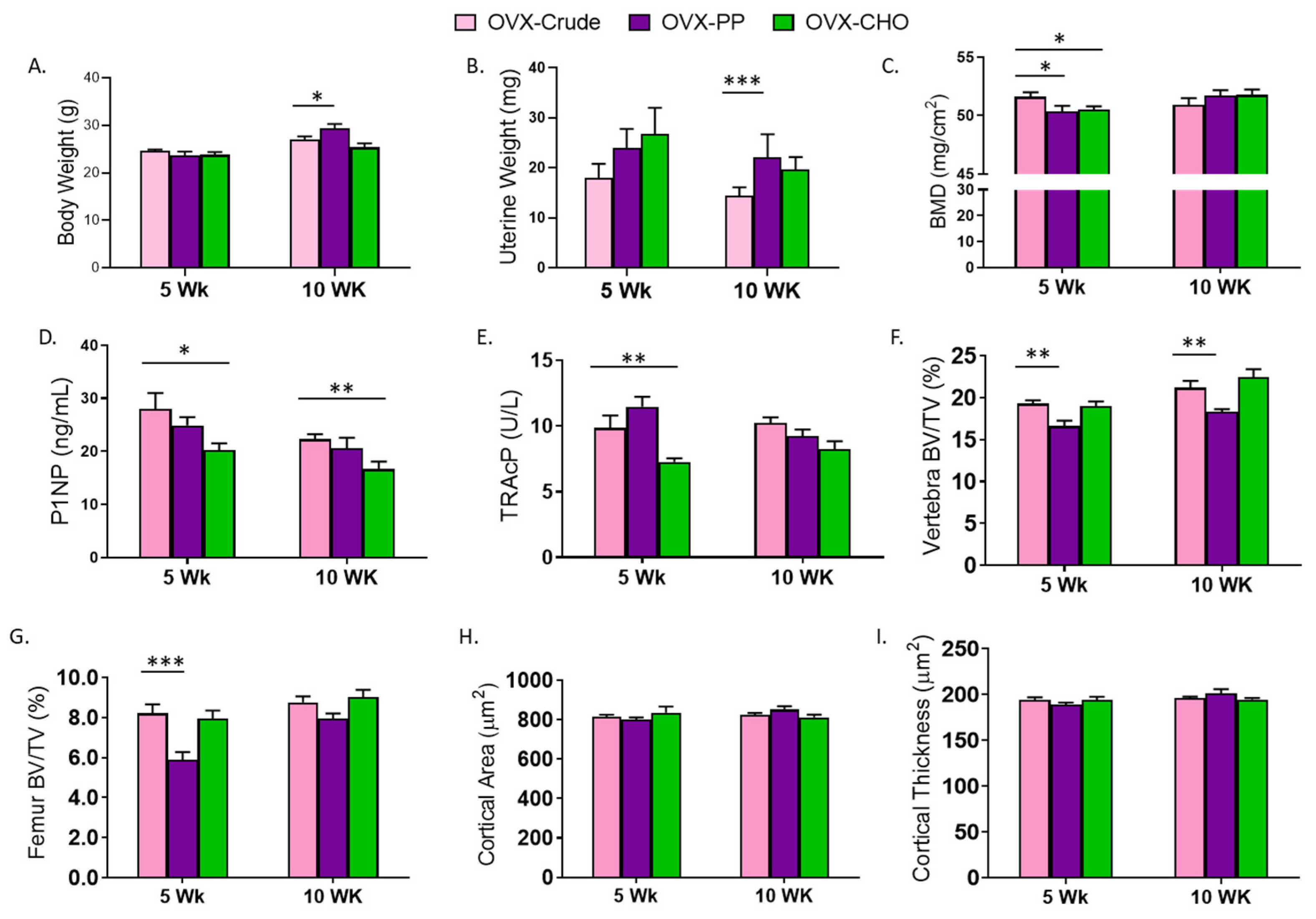
3.3. PP and CHO Fractions Alter Body Weight, Bone Density, Biomarkers, and Trabecular and Cortical Bone Microarchitecture
3.4. Comparison of PP and CHO Effects to the Crude Extract on Bone Structural and Metabolic Outcomes
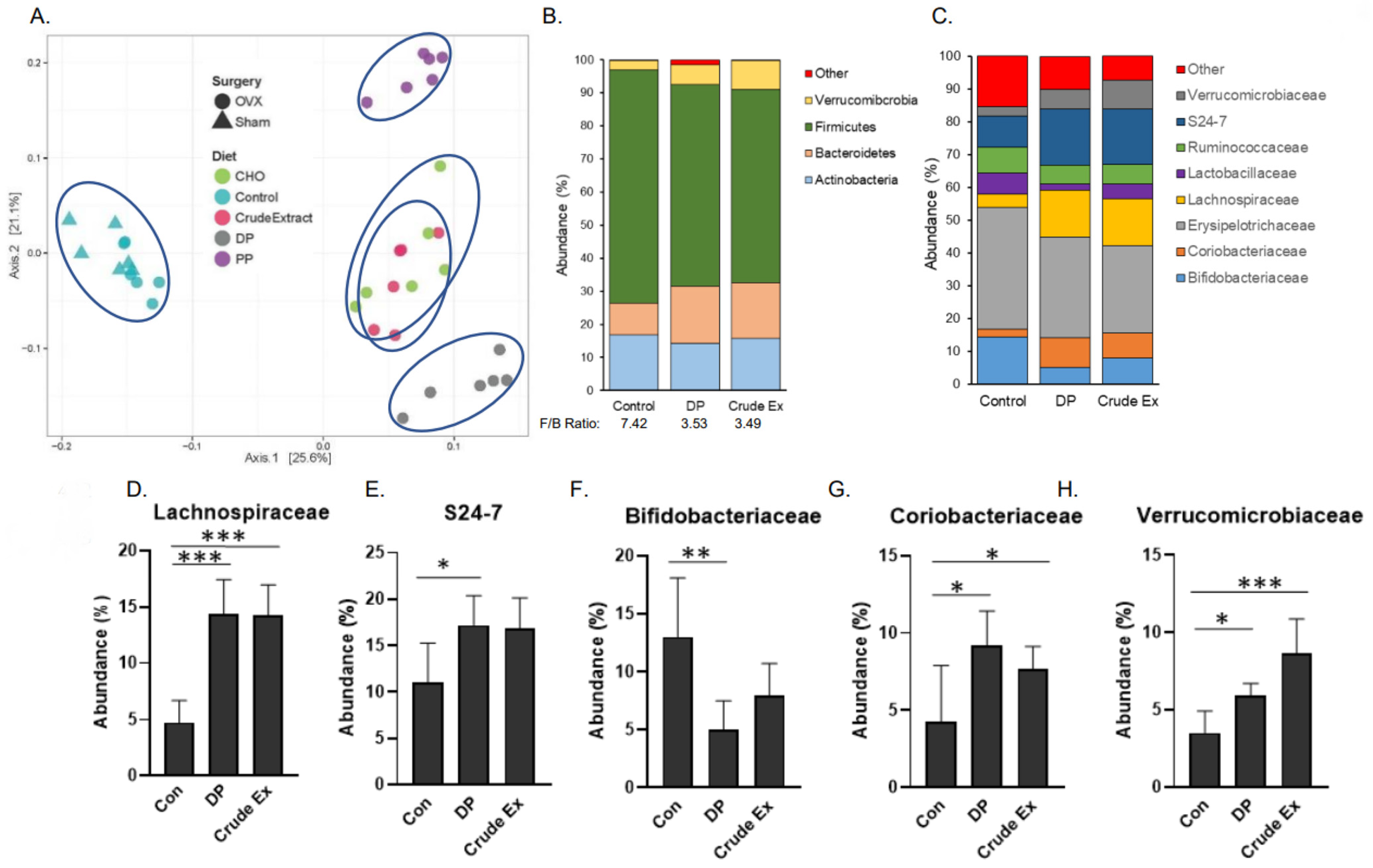
3.5. Gut Microbiota and Dietary Supplementation with Dried Plum and Its Bioactive Components
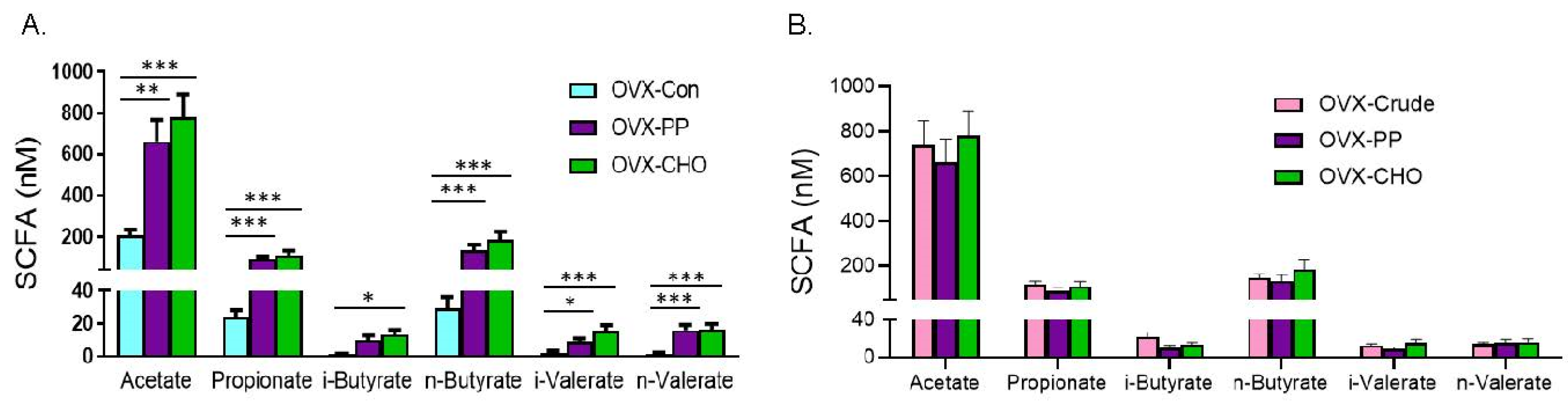
3.6. Short Chain Fatty Acids Production Induced by Dried Plum and Its Bioactive Components
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rizzoli, R.; Biver, E.; Brennan-Speranza, T.C. Nutritional intake and bone health. Lancet Diabetes Endocrinol. 2021, 9, 606–621. [Google Scholar] [CrossRef]
- Whisner, C.M.; Castillo, L.F. Prebiotics, Bone and Mineral Metabolism. Calcif. Tissue Int. 2018, 102, 443–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Sire, A.; de Sire, R.; Curci, C.; Castiglione, F.; Wahli, W. Role of Dietary Supplements and Probiotics in Modulating Microbiota and Bone Health: The Gut-Bone Axis. Cells 2022, 11, 743. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.O.; Heo, H.J.; Kim, Y.J.; Yang, H.S.; Lee, C.Y. Sweet and sour cherry phenolics and their protective effects on neuronal cells. J. Agric. Food Chem. 2005, 53, 9921–9927. [Google Scholar] [CrossRef]
- Fang, N.; Yu, S.; Prior, R.L. LC/MS/MS characterization of phenolic constituents in dried plums. J. Agric. Food Chem. 2002, 50, 3579–3585. [Google Scholar] [CrossRef]
- Almeida, M.; Han, L.; Martin-Millan, M.; Plotkin, L.I.; Stewart, S.A.; Roberson, P.K.; Kousteni, S.; O’Brien, C.A.; Bellido, T.; Parfitt, A.M.; et al. Skeletal involution by age-associated oxidative stress and its acceleration by loss of sex steroids. J. Biol. Chem. 2007, 282, 27285–27297. [Google Scholar] [CrossRef] [Green Version]
- Grassi, F.; Tell, G.; Robbie-Ryan, M.; Gao, Y.; Terauchi, M.; Yang, X.; Romanello, M.; Jones, D.P.; Weitzmann, M.N.; Pacifici, R. Oxidative stress causes bone loss in estrogen-deficient mice through enhanced bone marrow dendritic cell activation. Proc. Natl. Acad. Sci. USA 2007, 104, 15087–15092. [Google Scholar] [CrossRef] [Green Version]
- Hooshmand, S.; Chai, S.C.; Saadat, R.L.; Payton, M.E.; Brummel-Smith, K.; Arjmandi, B.H. Comparative effects of dried plum and dried apple on bone in postmenopausal women. Br. J. Nutr. 2011, 106, 923–930. [Google Scholar] [CrossRef]
- Arjmandi, B.H.; Khalil, D.A.; Lucas, E.A.; Georgis, A.; Stoecker, B.J.; Hardin, C.; Payton, M.E.; Wild, R.A. Dried plums improve indices of bone formation in postmenopausal women. J. Women’s Health Gend. Based Med. 2002, 11, 61–68. [Google Scholar] [CrossRef]
- Hooshmand, S.; Kern, M.; Metti, D.; Shamloufard, P.; Chai, S.C.; Johnson, S.A.; Payton, M.E.; Arjmandi, B.H. The effect of two doses of dried plum on bone density and bone biomarkers in osteopenic postmenopausal women: A randomized, controlled trial. Osteoporos. Int. 2016, 27, 2271–2279. [Google Scholar] [CrossRef]
- Hooshmand, S.; Gaffen, D.; Eisner, A.; Fajardo, J.; Payton, M.; Kern, M. Effects of 12 Months Consumption of 100 g Dried Plum (Prunes) on Bone Biomarkers, Density, and Strength in Men. J. Med. Food 2022, 25, 40–47. [Google Scholar] [CrossRef]
- George, K.S.; Munoz, J.; Ormsbee, L.T.; Akhavan, N.S.; Foley, E.M.; Siebert, S.C.; Kim, J.S.; Hickner, R.C.; Arjmandi, B.H. The Short-Term Effect of Prunes in Improving Bone in Men. Nutrients 2022, 14, 276. [Google Scholar] [CrossRef]
- Arjmandi, B.H.; Lucas, E.A.; Juma, S.; Soliman, A.; Stoecker, B.J.; Khalil, D.A.; Smith, B.J.; Wang, C. Dried plums prevent ovariectomy-induced bone loss in rats. JANA 2001, 4, 50–56. [Google Scholar]
- Bu, S.Y.; Lucas, E.A.; Franklin, M.; Marlow, D.; Brackett, D.J.; Boldrin, E.A.; Devareddy, L.; Arjmandi, B.H.; Smith, B.J. Restoration of bone mass and microarchitecture by dietary dried plum is comparable to PTH in osteopenic orchidectomized rats. Osteoporos. Int. 2007, 18, 931–942. [Google Scholar] [CrossRef]
- Halloran, B.P.; Wronski, T.J.; VonHerzen, D.C.; Chu, V.; Xia, X.; Pingel, J.E.; Williams, A.A.; Smith, B.J. Dietary Dried Plum Increases Bone Mass in Adult and Aged Male Mice. J. Nutr. 2010, 140, 1781–1787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, B.J.; Bu, S.Y.; Wang, Y.; Rendina, E.; Lim, Y.F.; Marlow, D.; Clarke, S.L.; Cullen, D.M.; Lucas, E.A. A comparative study of the bone metabolic response to dried plum supplementation and PTH treatment in adult, osteopenic ovariectomized rat. Bone 2014, 58, 151–159. [Google Scholar] [CrossRef]
- Mirza, F.; Lorenzo, J.; Drissi, H.; Lee, F.Y.; Soung, D.Y. Dried plum alleviates symptoms of inflammatory arthritis in TNF transgenic mice. J. Nutr. Biochem. 2018, 52, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.Y.; Kern, M.; Nakamichi-Lee, M.; Abbaspour, N.; Far, A.A.; Hooshmand, S. Dried Plum Consumption Improves Total Cholesterol and Antioxidant Capacity and Reduces Inflammation in Healthy Postmenopausal Women. J. Med. Food 2021, 24, 1161–1168. [Google Scholar] [CrossRef] [PubMed]
- Rendina, E.; Lim, Y.F.; Marlow, D.; Wang, Y.; Clarke, S.L.; Kuvibidila, S.; Lucas, E.A.; Smith, B.J. Dietary Supplementation with Dried Plum Prevents Ovariectomy-Induced Bone Loss in C57BL/6 Mice and Modulates the Immune Response. J. Nutr. Biochem. 2012, 23, 60–68. [Google Scholar] [CrossRef]
- Graef, J.L.; Rendina-Ruedy, E.; Crockett, E.K.; Ouyang, P.; Wu, L.; King, J.B.; Cichewicz, R.H.; Lin, D.; Lucas, E.A.; Smith, B.J. Osteoclast Differentiation is Downregulated by Select Polyphenolic Fractions from Dried Plum via Suppression of MAPKs and Nfatc1in Mouse C57BL/6 Primary Bone Marrow Cells. Curr. Dev. Nutr. 2017, 1, e000406. [Google Scholar]
- Graef, J.L.; Rendina-Ruedy, E.; Crockett, E.K.; Ouyang, P.; King, J.B.; Cichewicz, R.H.; Lucas, E.A.; Smith, B.J. Select Phenolic Fractions from Dried Plum Enhance Osteoblast Activity Through BMP-2 Signaling. J. Nutr. Biochem. 2017, 55, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Kimura, Y.; Ito, H.; Kawaji, M.; Ikami, T.; Hatano, T. Characterization and antioxidative properties of oligomeric proanthocyanidin from prunes, dried fruit of Prunus domestica L. Biosci. Biotechnol. Biochem. 2008, 72, 1615–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rendina, E.; Hembree, K.D.; Davis, M.R.; Marlow, D.; Clarke, S.L.; Halloran, B.P.; Lucas, E.A.; Smith, B.J. Dried plum’s unique capacity to reverse bone loss and alter bone metabolism in postmenopausal osteoporosis model. PLoS ONE 2013, 8, e60569. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.S.; Sang, S.; Lambert, J.D.; Lee, M.J. Bioavailability issues in studying the health effects of plant polyphenolic compounds. Mol. Nutr. Food Res. 2008, 52, S139–S151. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Scalbert, A.; Morand, C.; Remesy, C.; Jimenez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [Green Version]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Remesy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [CrossRef] [Green Version]
- Graef, J.L.; Ouyang, P.; Wang, Y.; Rendina-Ruedy, E.; Lerner, M.; Marlow, D.; Lucas, E.A.; Smith, B.J. Dried plum polyphenolic extract combined with vitamin K and potassium restores trabecular and cortical bone in osteopenic model of postmenopausal bone loss. J. Funct. Foods 2018, 42, 262–270. [Google Scholar] [CrossRef]
- Yan, J.; Herzog, J.W.; Tsang, K.; Brennan, C.A.; Bower, M.A.; Garrett, W.S.; Sartor, B.R.; Aliprantis, A.O.; Charles, J.F. Gut microbiota induce IGF-1 and promote bone formation and growth. Proc. Natl. Acad. Sci. USA 2016, 113, E7554–E7563. [Google Scholar] [CrossRef] [Green Version]
- Tyagi, A.M.; Yu, M.; Darby, T.M.; Vaccaro, C.; Li, J.Y.; Owens, J.A.; Hsu, E.; Adams, J.; Weitzmann, M.N.; Jones, R.M.; et al. The Microbial Metabolite Butyrate Stimulates Bone Formation via T Regulatory Cell-Mediated Regulation of WNT10B Expression. Immunity 2018, 49, 1116–1131. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Pal, S.; Paterson, C.W.; Li, J.Y.; Tyagi, A.M.; Adams, J.; Coopersmith, C.M.; Weitzmann, M.N.; Pacifici, R. Ovariectomy induces bone loss via microbial-dependent trafficking of intestinal TNF+ T cells and Th17 cells. J. Clin. Investig. 2021, 131, e143137. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-Ciocalteu reagent. Oxid. Antioxid. 1999, 299, 152–178. [Google Scholar]
- Ojo, B.; El-Rassi, G.D.; Payton, M.E.; Perkins-Veazie, P.; Clarke, S.; Smith, B.J.; Lucas, E.A. Mango Supplementation Modulates Gut Microbial Dysbiosis and Short-Chain Fatty Acid Production Independent of Body Weight Reduction in C57BL/6 Mice Fed a High-Fat Diet. J. Nutr. 2016, 146, 1483–1491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An Ordination of the Upland Forest Communities in Southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R package for reproducible interactive analysis and graphicsof microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Deyhim, F.; Stoecker, B.J.; Brusewitz, G.H.; Devareddy, L.; Arjmandi, B.H. Dried plum reverses bone loss in an osteopenic rat model of osteoporosis. Menopause 2005, 12, 755–762. [Google Scholar] [CrossRef]
- Glatt, V.; Canalis, E.; Stadmeyer, L.; Bouxsein, M.L. Age-related changes in trabecular architecture differ in female and male C57BL/6J mice. J. Bone Miner. Res. 2007, 22, 1197–1207. [Google Scholar] [CrossRef]
- Smith, B.J.; Graef, J.L.; Wronski, T.J.; Rendina, E.; Williams, A.A.; Clark, K.A.; Clarke, S.L.; Lucas, E.A.; Halloran, B.P. Effects of dried plum supplementation on bone metabolism in adult C57BL/6 male mice. Calcif. Tissue Int. 2014, 94, 442–453. [Google Scholar] [CrossRef] [Green Version]
- Bu, S.Y.; Lerner, M.; Stoecker, B.J.; Boldrin, E.; Brackett, D.J.; Lucas, E.A.; Smith, B.J. Dried Plum Polyphenols Inhibit Osteoclastogenesis by Downregulating NFATc1 and Inflammatory Mediators. Calcif. Tissue Int. 2008, 82, 475–488. [Google Scholar] [CrossRef] [PubMed]
- Vacca, M.; Celano, G.; Calabrese, F.M.; Portincasa, P.; Gobbetti, M.; De Angelis, M. The Controversial Role of Human Gut Lachnospiraceae. Microorganisms 2020, 8, 573. [Google Scholar] [CrossRef] [PubMed]
- Guglielmetti, S.; Fracassetti, D.; Taverniti, V.; Del Bo, C.; Vendrame, S.; Klimis-Zacas, D.; Arioli, S.; Riso, P.; Porrini, M. Differential modulation of human intestinal bifidobacterium populations after consumption of a wild blueberry (Vaccinium angustifolium) drink. J. Agric. Food Chem. 2013, 61, 8134–8140. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Daza, M.C.; Daoust, L.; Boutkrabt, L.; Pilon, G.; Varin, T.; Dudonne, S.; Levy, E.; Marette, A.; Roy, D.; Desjardins, Y. Wild blueberry proanthocyanidins shape distinct gut microbiota profile and influence glucose homeostasis and intestinal phenotypes in high-fat high-sucrose fed mice. Sci. Rep. 2020, 10, 2217. [Google Scholar] [CrossRef] [Green Version]
- Louis, P.; Flint, H.J. Formation of propionate and butyrate by the human colonic microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef] [Green Version]
- Lagkouvardos, I.; Lesker, T.R.; Hitch, T.C.A.; Galvez, E.J.C.; Smit, N.; Neuhaus, K.; Wang, J.; Baines, J.F.; Abt, B.; Stecher, B.; et al. Sequence and cultivation study of Muribaculaceae reveals novel species, host preference, and functional potential of this yet undescribed family. Microbiome 2019, 7, 28. [Google Scholar] [CrossRef] [Green Version]
- Lever, E.; Scott, S.M.; Louis, P.; Emery, P.W.; Whelan, K. The effect of prunes on stool output, gut transit time and gastrointestinal microbiota: A randomised controlled trial. Clin. Nutr. 2019, 38, 165–173. [Google Scholar] [CrossRef] [Green Version]
- Kaakoush, N.O. Insights into the Role of Erysipelotrichaceae in the Human Host. Front. Cell. Infect. Microbiol. 2015, 5, 84. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Wu, N.; Wang, X.; Chi, Y.; Zhang, Y.; Qiu, X.; Hu, Y.; Li, J.; Liu, Y. Dysbiosis gut microbiota associated with inflammation and impaired mucosal immune function in intestine of humans with non-alcoholic fatty liver disease. Sci. Rep. 2015, 5, 8096. [Google Scholar] [CrossRef]
- Lucas, S.; Omata, Y.; Hofmann, J.; Bottcher, M.; Iljazovic, A.; Sarter, K.; Albrecht, O.; Schulz, O.; Krishnacoumar, B.; Kronke, G.; et al. Short-chain fatty acids regulate systemic bone mass and protect from pathological bone loss. Nat. Commun. 2018, 9, 55. [Google Scholar] [CrossRef] [Green Version]
- Li, J.Y.; Chassaing, B.; Tyagi, A.M.; Vaccaro, C.; Luo, T.; Adams, J.; Darby, T.M.; Weitzmann, M.N.; Mulle, J.G.; Gewirtz, A.T.; et al. Sex steroid deficiency-associated bone loss is microbiota dependent and prevented by probiotics. J. Clin. Investig. 2016, 126, 2049–2063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dikeman, C.L.; Bauer, L.L.; Fahey, G.C., Jr. Carbohydrate composition of selected plum/prune preparations. J. Agric. Food Chem. 2004, 52, 853–859. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sham-Con | OVX-Con | OVX-DP | OVX-Crude | p-Value | |
|---|---|---|---|---|---|
| 5 Weeks | |||||
| Body weight | |||||
| Pre-treatment (g) | 20.80 ± 0.27 b | 22.30 ± 0.29 a | 21.75 ± 0.34 a | 22.25 ± 0.20 a | 0.0011 |
| Final (g) | 21.52 ± 0.30 d | 26.18 ± 0.65 a | 23.13 ± 0.73 c | 24.65 ± 0.29 b | <0.0001 |
| Uterine weight (mg) | 112.12 ± 3.73 a | 24.38 ± 5.10 b | 26.54 ± 5.72 b | 21.93 ± 4.70 b | 0.0016 |
| Whole body BMD (mg/cm2) | 51.65 ± 0.46 a | 48.62 ± 0.29 c | 50.55 ± 0.34 b | 51.61 ± 0.38 a | <0.0001 |
| Bone biomarkers | |||||
| P1NP (ng/mL) | 56.91 ± 7.80 a | 32.97 ± 2.62 b | 26.87 ± 1.70 b | 27.97 ± 3.06 b | <0.0001 |
| TRAcP (U/L) | 11.28 ± 1.01 a | 10.89 ± 0.72 a | 7.72 ± 0.39 b | 9.85 ± 0.96 ab | 0.0119 |
| Cecal content weight (g) | 0.48 ± 0.04 c | 0.48 ± 0.03 c | 0.91 ± 0.07 a | 0.67 ± 0.05 b | <0.0001 |
| 10 Weeks | |||||
| Body weight | |||||
| Pre-treatment (g) | 20.15 ± 0.23 b | 21.76 ± 0.19 a | 21.71 ± 0.39 a | 22.07 ± 0.25 a | <0.0001 |
| Final (g) | 21.93 ± 0.40 c | 28.50 ± 0.63 a | 24.40 ± 0.62 b | 27.02 ± 0.68 a | <0.0001 |
| Uterine weight (mg) | 80.86 ± 5.94 a | 31.86 ± 6.54 b | 28.85 ± 6.30 b | 14.43 ± 1.66 b | <0.0001 |
| Whole body BMD (mg/cm2) | 52.00 ± 0.29 a | 49.09 ± 0.37 b | 51.16 ± 0.53 a | 50.92 ± 0.57 a | 0.0001 |
| Bone biomarkers | |||||
| P1NP (ng/mL) | 27.67 ± 2.27 a | 22.15 ± 0.77 ab | 17.73 ± 1.38 b | 22.32 ± 0.94 ab | 0.0003 |
| TRAcP (U/L) | 8.09 ± 0.21 b | 10.22 ± 0.44 a | 7.03 ± 0.33 c | 7.96 ± 0.44 bc | <0.0001 |
| Cecal content weight (g) | 0.53 ± 0.04 c | 0.42 ± 0.04 c | 1.08 ± 0.05 a | 0.73 ± 0.04 b | <0.0001 |
| Sham-Con | OVX-Con | OVX-DP | OVX-Crude | p-Value | |
|---|---|---|---|---|---|
| 5 weeks | |||||
| Distal femur metaphysis | |||||
| BV/TV (%) | 8.49 ± 0.74 a | 5.28 ± 0.21 b | 8.30 ± 0.83 a | 8.21 ± 0.45 a | 0.0044 |
| TbTh (mm) | 0.048 ± 0.001 a | 0.041 ± 0.001 b | 0.039 ± 0.001 b | 0.041 ± 0.001 b | <0.0001 |
| TbN (1/mm2) | 3.43 ± 0.07 b | 3.09 ± 0.10 c | 3.83 ± 0.13 a | 3.81 ± 0.04 a | <0.0001 |
| TbSp (mm) | 0.29 ± 0.01 b | 0.32 ± 0.01 a | 0.26 ± 0.01 c | 0.26 ± 0.003 c | <0.0001 |
| Lumbar vertebra | |||||
| BV/TV (%) | 20.31 ± 0.69 a | 15.38 ± 0.64 b | 20.54 ± 0.41 a | 19.31 ± 0.39 a | <0.0001 |
| TbTh (mm) | 0.048 ± 0.001 a | 0.043 ± 0.001 b | 0.049 ± 0.001 a | 0.048 ± 0.0004 a | <0.0001 |
| TbN (1/mm2) | 4.41 ± 0.08 a | 3.94 ± 0.09 b | 4.10 ± 0.05 b | 3.95 ± 0.06 b | 0.0034 |
| TbSp (mm) | 0.228 ± 0.005 b | 0.260 ± 0.01 a | 0.245 ± 0.003 a | 0.255 ± 0.004 a | 0.0002 |
| Femur mid-diaphysis | |||||
| Cortical area (μm2) | 813.69 ± 15.66 | 780.03 ± 10.22 | 775.52 ± 14.54 | 815.34 ± 10.25 | 0.0601 |
| Cortical thickness (μm) | 193.11 ± 2.55 a | 184.44 ± 1.94 b | 186.00 ± 1.35 b | 193.90 ± 2.71 a | 0.0093 |
| 10 weeks | |||||
| Distal femur metaphysis | |||||
| BV/TV (%) | 6.99 ± 0.37 c | 5.63 ± 0.20 d | 10.17 ± 0.58 a | 8.76 ± 0.30 b | <0.0001 |
| TbTh (mm) | 0.044 ± 0.001 | 0.045 ± 0.001 | 0.043 ± 0.001 | 0.045 ± 0.001 | 0.6237 |
| TbN (1/mm2) | 3.94 ± 0.13 b | 2.81 ± 0.08 c | 3.66 ± 0.14 a | 3.28 ± 0.08 b | <0.0001 |
| TbSp (mm) | 0.32 ± 0.01 b | 0.36 ± 0.01 a | 0.27 ± 0.01 c | 0.30 ± 0.01 b | <0.0001 |
| Lumbar vertebra | |||||
| BV/TV (%) | 21.96 ± 0.95 a | 14.98 ± 0.43 b | 22.18 ± 0.95 a | 21.19 ± 0.84a | <0.0001 |
| TbTh (mm) | 0.053 ± 0.001 a | 0.047 ±0.0004 c | 0.050 ± 0.001 ab | 0.050 ± 0.001 bc | 0.0010 |
| TbN (1/mm2) | 3.94 ± 0.13 a | 3.37 ± 0.08 b | 4.00 ± 0.13 a | 3.88 ± 0.19 a | 0.0244 |
| TbSp (mm) | 0.26 ± 0.01 b | 0.30 ± 0.01 a | 0.25 ± 0.01 b | 0.26 ± 0.01 b | 0.0284 |
| Femur mid-diaphysis | |||||
| Cortical area (um2) | 823.59 ± 7.53 a | 785.12 ± 7.28 ab | 745.48 ± 48.03 b | 825.00 ± 9.80 a | 0.0390 |
| Cortical thickness (um) | 196.88 ± 1.79 a | 186.00 ± 1.18 ab | 174.40 ± 12.20 b | 196.00 ± 1.6 a | 0.0162 |
| Sham-Con | OVX-Con | OVX-DP | OVX-Crude | p Value | |
|---|---|---|---|---|---|
| Total SCFA (nM) | 341.15 ± 73.05 c | 264.92 ± 38.68 c | 3092.90 ± 550.92 a | 1054.95 ± 115.04 b | <0.0001 |
| Acetate (nM) | 270.92 ± 56.99 c | 206.96 ± 27.08 c | 2259.18 ± 437.21 a | 739.25 ± 107.79 b | <0.0001 |
| Propionate (nM) | 29.86 ± 6.87 c | 24.08 ± 4.19 c | 374.30 ± 64.38 a | 119.16 ± 13.27 b | <0.0001 |
| i-butyrate (nM) | 1.98 ± 1.30 b | 1.01 ± 0.69 b | 6.61 ± 3.27 b | 21.69 ± 5.49 a | 0.0090 |
| n-butyrate (nM) | 31.76 ± 6.62 c | 29.25 ± 6.90 c | 421.53 ± 70.79 a | 147.40 ± 17.15 b | <0.0001 |
| i-valerate (nM) | 4.65 ± 1.45 bc | 2.12 ± 1.27 c | 9.10 ± 2.95 ab | 12.80 ± 1.92 a | 0.0109 |
| n-valerate (nM) | 1.98 ± 1.30 c | 1.49 ± 0.75 c | 22.18 ± 4.01 a | 14.65 ± 1.64 b | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smith, B.J.; Hatter, B.; Washburn, K.; Graef-Downard, J.; Ojo, B.A.; El-Rassi, G.D.; Cichewicz, R.H.; Payton, M.; Lucas, E.A. Dried Plum’s Polyphenolic Compounds and Carbohydrates Contribute to Its Osteoprotective Effects and Exhibit Prebiotic Activity in Estrogen Deficient C57BL/6 Mice. Nutrients 2022, 14, 1685. https://doi.org/10.3390/nu14091685
Smith BJ, Hatter B, Washburn K, Graef-Downard J, Ojo BA, El-Rassi GD, Cichewicz RH, Payton M, Lucas EA. Dried Plum’s Polyphenolic Compounds and Carbohydrates Contribute to Its Osteoprotective Effects and Exhibit Prebiotic Activity in Estrogen Deficient C57BL/6 Mice. Nutrients. 2022; 14(9):1685. https://doi.org/10.3390/nu14091685
Chicago/Turabian StyleSmith, Brenda J., Bethany Hatter, Karley Washburn, Jennifer Graef-Downard, Babajide A. Ojo, Guadalupe Davila El-Rassi, Robert H. Cichewicz, Mark Payton, and Edralin A. Lucas. 2022. "Dried Plum’s Polyphenolic Compounds and Carbohydrates Contribute to Its Osteoprotective Effects and Exhibit Prebiotic Activity in Estrogen Deficient C57BL/6 Mice" Nutrients 14, no. 9: 1685. https://doi.org/10.3390/nu14091685