
Integrative Metabolomic Characterization Reveals the Mediating Effect of Bifidobacterium breve on Amino Acid Metabolism in a Mouse Model of Alzheimer’s Disease

 , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Bacterial Treatment
2.2. Animals
2.3. Serum and Hippocampus Collection
2.4. Metabolomics
2.4.1. Metabolite Sample Preparation
2.4.2. Ultraperformance Liquid Chromatography-Mass Spectrometry (UPLC-MS) Analysis Parameters
2.4.3. Metabolomic Data Analysis
3. Results
3.1. Quality Assessment and Annotated Metabolites of the Overall Metabolome
3.2. B. breve CCFM1025 Supplementation Modulated the Aβ-Induced Serum Metabolome
3.3. B. breve CCFM1025 Supplementation Restored the Aβ-Induced Hippocampal Metabolome
3.4. Metabolic Alterations Were Identified in CCFM1025 Mice Compared with Model Mice
3.5. B. breve CCFM1025 Inhibited Aβ-Induced Neuroinflammation by Regulating Amino Acid Metabolism
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ballard, C.; Gauthier, S.; Corbett, A.; Brayne, C.; Aarsland, D.; Jones, E. Alzheimer’s disease. Lancet 2011, 377, 1019–1031. [Google Scholar] [CrossRef]
- De Strooper, B.; Karran, E. The Cellular Phase of Alzheimer’s Disease. Cell 2016, 164, 603–615. [Google Scholar] [CrossRef] [Green Version]
- Scheltens, P.; Blennow, K.; Breteler, M.M.B.; de Strooper, B.; Frisoni, G.B.; Salloway, S.; Van der Flier, W.M. Alzheimer’s disease. Lancet 2016, 388, 505–517. [Google Scholar] [CrossRef]
- Sanborn, V.; Azcarate-Peril, M.A.; Updegraff, J.; Manderino, L.; Gunstad, J. Randomized Clinical Trial Examining the Impact of Lactobacillus rhamnosus GG Probiotic Supplementation on Cognitive Functioning in Middle-aged and Older Adults. Neuropsychiatr. Dis. Treat. 2020, 16, 2765–2777. [Google Scholar] [CrossRef]
- Xiao, J.; Katsumata, N.; Bernier, F.; Ohno, K.; Kaneko, T. Probiotic Bifidobacterium breve in Improving Cognitive Functions of Older Adults with Suspected Mild Cognitive Impairment: A Randomized, Double-Blind, Placebo-Controlled Trial. J. Alzheimer’s Dis. 2020, 77, 139–147. [Google Scholar] [CrossRef]
- Zhu, G.; Zhao, J.; Zhang, H.; Chen, W.; Wang, G. Administration of Bifidobacterium breve Improves the Brain Function of Aβ(1-42)-Treated Mice via the Modulation of the Gut Microbiome. Nutrients 2021, 13, 1602. [Google Scholar] [CrossRef]
- Modern analytical techniques in metabolomics analysis. Analyst 2011, 137, 293–300.
- Ding, J.; Ji, J.; Rabow, Z.; Shen, T.; Folz, J.; Brydges, C.R.; Fan, S.; Lu, X.; Mehta, S.; Showalter, M.R.; et al. A metabolome atlas of the aging mouse brain. Nat. Commun. 2021, 12, 6021. [Google Scholar] [CrossRef]
- Lavelle, A.; Sokol, H. Gut microbiota-derived metabolites as key actors in inflammatory bowel disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Nasaruddin, M.B.; Elliott, C.T.; McGuinness, B. Alzheimer’s disease–like pathology has transient effects on the brain and blood metabolome. Neurobiol. Aging 2016, 38, 151–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Domínguez, R.; García-Barrera, T.; Vitorica, J.; Gómez-Ariza, J.L. Metabolomic screening of regional brain alterations in the APP/PS1 transgenic model of Alzheimer’s disease by direct infusion mass spectrometry. J. Pharm. Biomed. Anal. 2015, 102, 425–435. [Google Scholar] [CrossRef] [PubMed]
- González-Domínguez, R.; García-Barrera, T.; Gómez-Ariza, J. Metabolomic study of lipids in serum for biomarker discovery in Alzheimer’s disease using direct infusion mass spectrometry. J. Pharm. Biomed. Anal. 2014, 98, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Guiraud, S.P.; Montoliu, I.; Da Silva, L.; Dayon, L.; Galindo, A.N.; Corthésy, J.; Kussmann, M.; Martin, F.P. High-throughput and simultaneous quantitative analysis of homocysteine-methionine cycle metabolites and co-factors in blood plasma and cerebrospinal fluid by isotope dilution LC-MS/MS. Anal. Bioanal. Chem. 2017, 409, 295–305. [Google Scholar] [CrossRef] [Green Version]
- Yuan, M.; Breitkopf, S.B.; Yang, X.; Asara, J.M. A positive/negative ion-switching, targeted mass spectrometry-based metabolomics platform for bodily fluids, cells, and fresh and fixed tissue. Nat. Protoc. 2012, 7, 872–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, C.A.; Vuong, H.E.; Yano, J.M.; Liang, Q.Y.; Nusbaum, D.J.; Hsiao, E.Y. The Gut Microbiota Mediates the Anti-Seizure Effects of the Ketogenic Diet. Cell 2018, 173, 1728–1741.e1713. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Geng, W.; Sun, H.; Liu, C.; Huang, F.; Cao, J.; Xia, L.; Zhao, H.; Zhai, J.; Li, Q.; et al. Integrative metabolomic characterisation identifies altered portal vein serum metabolome contributing to human hepatocellular carcinoma. Gut 2021, 1–11. [Google Scholar] [CrossRef]
- Sreekumar, A.; Poisson, L.M.; Rajendiran, T.M.; Khan, A.P.; Cao, Q.; Yu, J.; Laxman, B.; Mehra, R.; Lonigro, R.J.; Li, Y. Metabolomic profiles delineate potential role for sarcosine in prostate cancer progression. Nature 2009, 457, 910–914. [Google Scholar] [CrossRef] [Green Version]
- Boehme, M.; Guzzetta, K.E.; Bastiaanssen, T.F.S.; van de Wouw, M.; Moloney, G.M.; Gual-Grau, A.; Spichak, S.; Olavarría-Ramírez, L.; Fitzgerald, P.; Morillas, E.; et al. Microbiota from young mice counteracts selective age-associated behavioral deficits. Nat. Aging 2021, 1, 666–676. [Google Scholar] [CrossRef]
- Canfield, C.A.; Bradshaw, P.C. Amino acids in the regulation of aging and aging-related diseases. Transl. Med. Aging 2019, 3, 70–89. [Google Scholar] [CrossRef]
- Krishnan, S.; Ding, Y.; Saedi, N.; Choi, M.; Sridharan, G.V.; Sherr, D.H.; Yarmush, M.L.; Alaniz, R.C.; Jayaraman, A.; Lee, K. Gut Microbiota-Derived Tryptophan Metabolites Modulate Inflammatory Response in Hepatocytes and Macrophages. Cell Rep. 2018, 23, 1099–1111. [Google Scholar] [CrossRef]
- Wilkins, J.M.; Trushina, E. Application of Metabolomics in Alzheimer’s Disease. Front. Neurol. 2017, 8, 719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McQuail, J.A.; Frazier, C.J.; Bizon, J.L. Molecular aspects of age-related cognitive decline: The role of GABA signaling. Trends Mol. Med. 2015, 21, 450–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braniste, V.; Al-Asmakh, M.; Kowal, C.; Anuar, F.; Abbaspour, A.; Toth, M.; Korecka, A.; Bakocevic, N.; Ng, L.G.; Kundu, P. The gut microbiota influences blood-brain barrier permeability in mice. Sci. Transl. Med. 2014, 6, 263ra158. [Google Scholar] [CrossRef] [Green Version]
- Puigdellívol, M.; Allendorf, D.H.; Brown, G.C. Sialylation and Galectin-3 in Microglia-Mediated Neuroinflammation and Neurodegeneration. Front. Cell. Neurosci. 2020, 14, 162. [Google Scholar] [CrossRef] [PubMed]
- Malinski, T. Nitric oxide and nitroxidative stress in Alzheimer’s disease. J. Alzheimers Dis. 2007, 11, 207–218. [Google Scholar] [CrossRef]
- Baird, A.L.; Westwood, S.; Lovestone, S. Blood-Based Proteomic Biomarkers of Alzheimer’s Disease Pathology. Front. Neurol. 2015, 6, 236. [Google Scholar] [CrossRef] [Green Version]
- Rosas, H.D.; Doros, G.; Bhasin, S.; Thomas, B.; Gevorkian, S.; Malarick, K.; Matson, W.; Hersch, S.M. A systems-level “misunderstanding”: The plasma metabolome in Huntington’s disease. Ann. Clin. Transl. Neurol. 2015, 2, 756–768. [Google Scholar] [CrossRef]
- Wang, X.; Sun, G.; Feng, T.; Zhang, J.; Huang, X.; Wang, T.; Xie, Z.; Chu, X.; Yang, J.; Wang, H.; et al. Sodium oligomannate therapeutically remodels gut microbiota and suppresses gut bacterial amino acids-shaped neuroinflammation to inhibit Alzheimer’s disease progression. Cell Res. 2019, 29, 787–803. [Google Scholar] [CrossRef]
- Tian, P.; Bastiaanssen, T.F.S.; Song, L.; Jiang, B.; Zhang, X.; Zhao, J.; Zhang, H.; Chen, W.; Cryan, J.F.; Wang, G. Unraveling the Microbial Mechanisms Underlying the Psychobiotic Potential of a Bifidobacterium breve Strain. Mol. Nutr. Food Res. 2021, 65, e2000704. [Google Scholar] [CrossRef] [PubMed]
- Janik, R.; Thomason, L.A.M.; Stanisz, A.M.; Forsythe, P.; Bienenstock, J.; Stanisz, G.J. Magnetic resonance spectroscopy reveals oral Lactobacillus promotion of increases in brain GABA, N-acetyl aspartate and glutamate. Neuroimage 2016, 125, 988–995. [Google Scholar] [CrossRef]
- Yudkoff, M.; Daikhin, Y.; Nissim, I.; Lazarow, A.; Nissim, I. Ketogenic diet, amino acid metabolism, and seizure control. J. Neurosci. Res. 2001, 66, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Dalangin, R.; Kim, A.; Campbell, R.E. The Role of Amino Acids in Neurotransmission and Fluorescent Tools for Their Detection. Int. J. Mol. Sci. 2020, 21, 6197. [Google Scholar] [CrossRef] [PubMed]
- González-Domínguez, R.; González-Domínguez, Á.; Sayago, A.; González-Sanz, J.D.; Lechuga-Sancho, A.M.; Fernández-Recamales, Á. Mechanistic Insights into Alzheimer’s Disease Unveiled through the Investigation of Disturbances in Central Metabolites and Metabolic Pathways. Biomedicines 2021, 9, 298. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, A.E.; Pinkard, O.; Coller, J.M. tRNA Metabolism and Neurodevelopmental Disorders. Annu. Rev. Genom. Hum. Genet. 2019, 20, 359–387. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
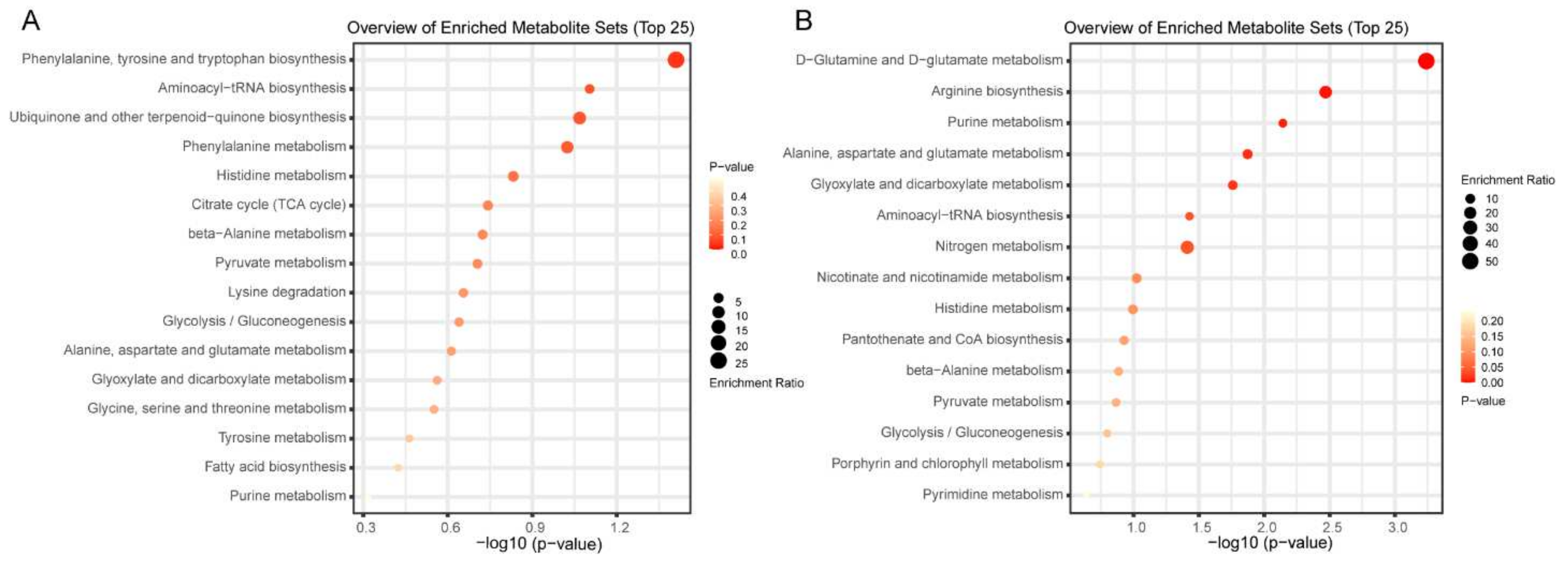{kind=link}
{kind=link}
| Metab_id | Metabolite | OPLS-DA VIP | p Value | Fold Change | HMDB ID | KEGG ID |
|---|---|---|---|---|---|---|
| N173 | Plumbagin | 2.09326 | 0.00678447 | 1.58203 | HMDB0035291 | - |
| N232 | Ascorbicacid-2-sulfate | 1.86579 | 0.0157301 | 0.761099 | - | - |
| N187 | Decanoic acid | 1.82526 | 0.0209732 | 0.770978 | HMDB0000511 | C01571 |
| N96 | No-noic acid | 1.84173 | 0.021134 | 0.729044 | HMDB0000847 | C01601 |
| N219 | L-Tyrosine | 1.70911 | 0.0292742 | 1.21693 | HMDB0000158 | C00082 |
| N289 | 2-(2-Acetoxy-2-oxoethyl)-2-hydroxysuccinic acid | 1.71639 | 0.0326245 | 2.0757 | - | - |
| N271 | 5-[(2Z,8Z)-2,8-Pentadecadien-1-yl]-1,3-benzenediol | 1.64572 | 0.0396629 | 0.662185 | - | - |
| N6 | L-(+)-Lactic acid | 1.6948 | 0.0423889 | 1.35085 | HMDB0000190 | C00186 |
| N55 | Citric acid | 1.7116 | 0.0433744 | 0.563898 | HMDB0000094 | C00158 |
| N98 | Cytidine;1-beta-delta-Ribofuranosyl-Cytosine | 1.59184 | 0.0450476 | 0.738744 | - | - |
| N135 | [FA(18:4)]6Z_9Z_12Z_15Z-octadecatetraenoicacid | 1.6114 | 0.046132 | 0.605545 | - | - |
| P93 | Hexanoylglycine | 2.19498 | 0.00415879 | 0.57031 | HMDB0000701 | - |
| P132 | (4R)-5-Hydroxy-L-leucine | 2.13034 | 0.00443283 | 0.766452 | HMDB0000450 | C16741 |
| P15 | Betaine | 1.99501 | 0.00767035 | 1.43377 | HMDB0000043 | C00719 |
| P95 | L-Histidine | 1.88292 | 0.0125969 | 1.42264 | HMDB0000177 | C00135 |
| P280 | Capsidiol | 1.90628 | 0.0165168 | 0.627435 | HMDB0002352 | C09627 |
| P209 | 4121 | 1.90749 | 0.0171203 | 0.579715 | HMDB0000462 | C01551 |
| P55 | N,N-Dimethyldecylamine N-oxide | 1.8191 | 0.0239789 | 1.96321 | HMDB0001466 | C01183 |
| P189 | 1-Methylhistidine | 1.74198 | 0.0325697 | 0.846794 | HMDB0000001 | C01152 |
| P113 | 2475675 | 1.69582 | 0.0366034 | 0.556372 | HMDB0015593 | - |
| Metab_id | Metabolite | OPLS-DAVIP | p Value | Foldchange | HMDB ID | KEGGID |
|---|---|---|---|---|---|---|
| N80 | Abieticacid | 3.69022 | 4.98E-11 | 845.997 | HMDB0000042 | C00033 |
| N92 | Hypoxanthine | 2.82776 | 0.00092 | 3.21625 | HMDB0000157 | C00262 |
| N74 | 1-Stearoyl-2-hydroxy-sn-glycero-3-PE | 2.52581 | 0.003066 | 0.751745 | - | - |
| N239 | 2-(2-Carboxyethyl)-4-methyl-5-pentyl-3-furoic acid | 2.48185 | 0.00567 | 0.435581 | - | - |
| N85 | D-(-)-Glutamine | 2.47921 | 0.00346 | 0.352861 | HMDB0000641 | C00064 |
| N23 | 2,4-di-tert-Butylphenol | 2.4217 | 0.005509 | 0.685152 | HMDB0013816 | - |
| N292 | 1-Palmitoyl-2-hydroxy-sn-glycero-3-PE | 2.34582 | 0.006262 | 0.834934 | - | - |
| N14 | L-Aspartic acid | 2.21739 | 0.011711 | 0.867782 | HMDB0000191 | C00049 |
| N3 | Dodecyl sulfate | 2.0113 | 0.031644 | 0.562855 | - | - |
| N238 | Cytidine;1-beta-delta-Ribofuranosyl-Cytosine | 1.96217 | 0.040588 | 1.23977 | - | - |
| N16 | Myristyl sulfate | 1.90313 | 0.042206 | 0.704926 | - | - |
| P1146 | Diethyl phthalate | 1.92305 | 0.046523 | 0.624987 | - | - |
| P743 | Dichloromethane | 1.81003 | 0.044936 | 1.67818 | HMDB0031548 | C02271 |
| P145 | Adenosine 5’-monophosphate | 2.00506 | 0.041507 | 1.67312 | HMDB0000045 | C00020 |
| P215 | 1-Oleoyl-2-hydroxy-sn-glycero-3-PE | 1.86473 | 0.041022 | 1.50993 | - | - |
| P628 | PAF C-18:1 | 1.9106 | 0.039774 | 1.54938 | - | - |
| P1068 | methyl 2,8-dihydroxy-6-(hydroxymethyl)-9-oxo-2,9-dihydro-1H-xanthene-1-carboxylate | 1.89251 | 0.039566 | 1.45483 | - | - |
| P109 | Adenosine 5’-monophosphate | 2.00566 | 0.034384 | 1.63448 | ||
| P1049 | Docosahexaenoyl Ethanolamide | 1.93602 | 0.032247 | 1.547 | HMDB0013658 | - |
| P144 | Sangivamycin | 2.03425 | 0.031676 | 1.63017 | - | - |
| P912 | 11-Nitro-1-undecene | 2.05666 | 0.025121 | 1.76411 | - | - |
| P1156 | 4049 | 2.05316 | 0.02406 | 1.65415 | ||
| P489 | Biliverdin | 1.97892 | 0.023273 | 7.11057 | HMDB0015624 | - |
| P1071 | mebutamate | 2.19096 | 0.018291 | 1.56646 | HMDB0001008 | C00500 |
| P800 | 1-Stearoyl-2-hydroxy-sn-glycero-3-PE | 2.12975 | 0.017473 | 1.70019 | - | - |
| P393 | 1-stearoyl-lysophosphatidylcholine | 2.24008 | 0.012303 | 1.67575 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, G.; Guo, M.; Zhao, J.; Zhang, H.; Wang, G.; Chen, W. Integrative Metabolomic Characterization Reveals the Mediating Effect of Bifidobacterium breve on Amino Acid Metabolism in a Mouse Model of Alzheimer’s Disease. Nutrients 2022, 14, 735. https://doi.org/10.3390/nu14040735
Zhu G, Guo M, Zhao J, Zhang H, Wang G, Chen W. Integrative Metabolomic Characterization Reveals the Mediating Effect of Bifidobacterium breve on Amino Acid Metabolism in a Mouse Model of Alzheimer’s Disease. Nutrients. 2022; 14(4):735. https://doi.org/10.3390/nu14040735
Chicago/Turabian StyleZhu, Guangsu, Min Guo, Jianxin Zhao, Hao Zhang, Gang Wang, and Wei Chen. 2022. "Integrative Metabolomic Characterization Reveals the Mediating Effect of Bifidobacterium breve on Amino Acid Metabolism in a Mouse Model of Alzheimer’s Disease" Nutrients 14, no. 4: 735. https://doi.org/10.3390/nu14040735