
Twelve Months of Time-Restricted Feeding Improves Cognition and Alters Microbiome Composition Independent of Macronutrient Composition

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects and Dietary Interventions
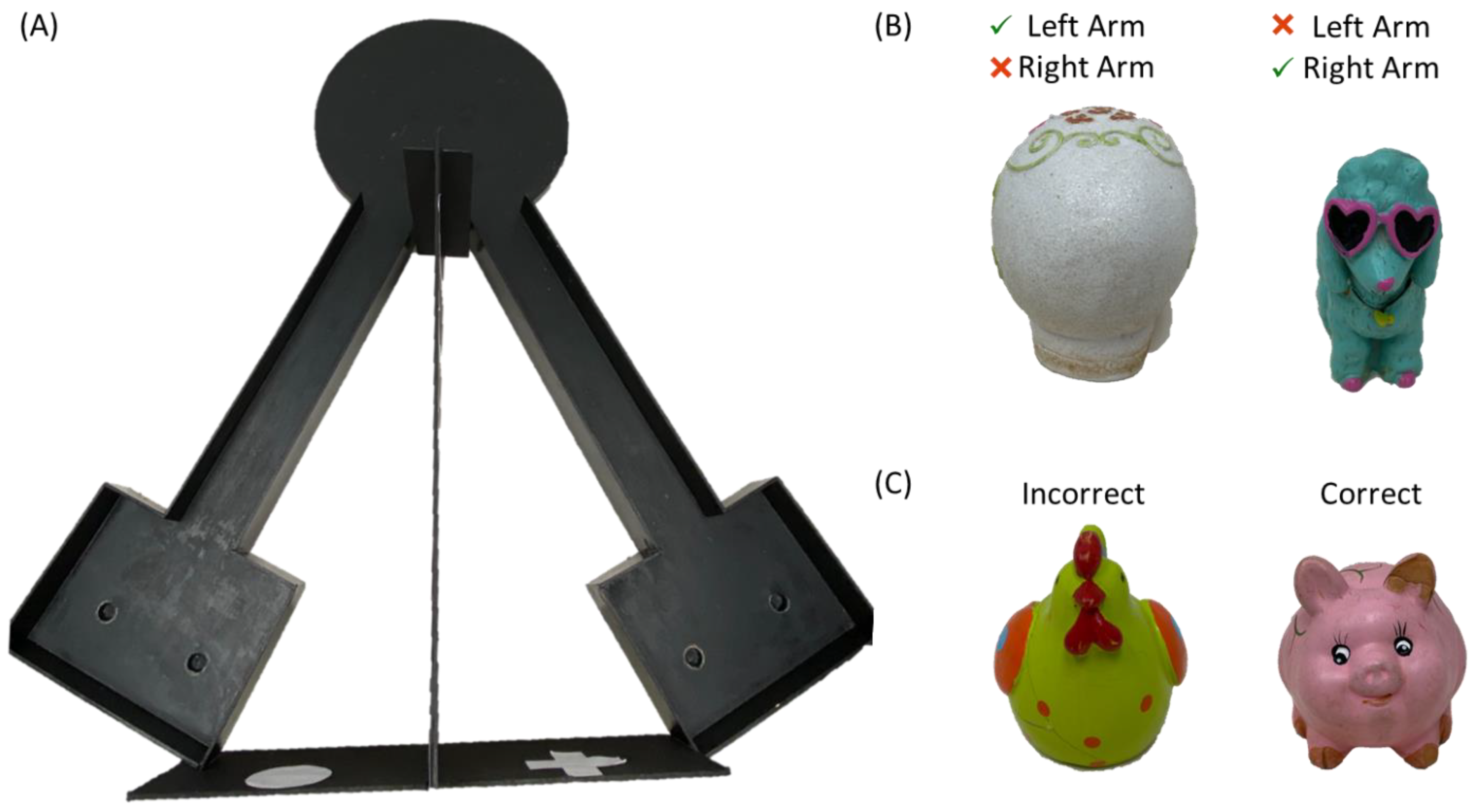
2.2. Behavioral Testing
2.3. Statistical Analysis
2.4. Fecal Microbiome Taxonomy
3. Results
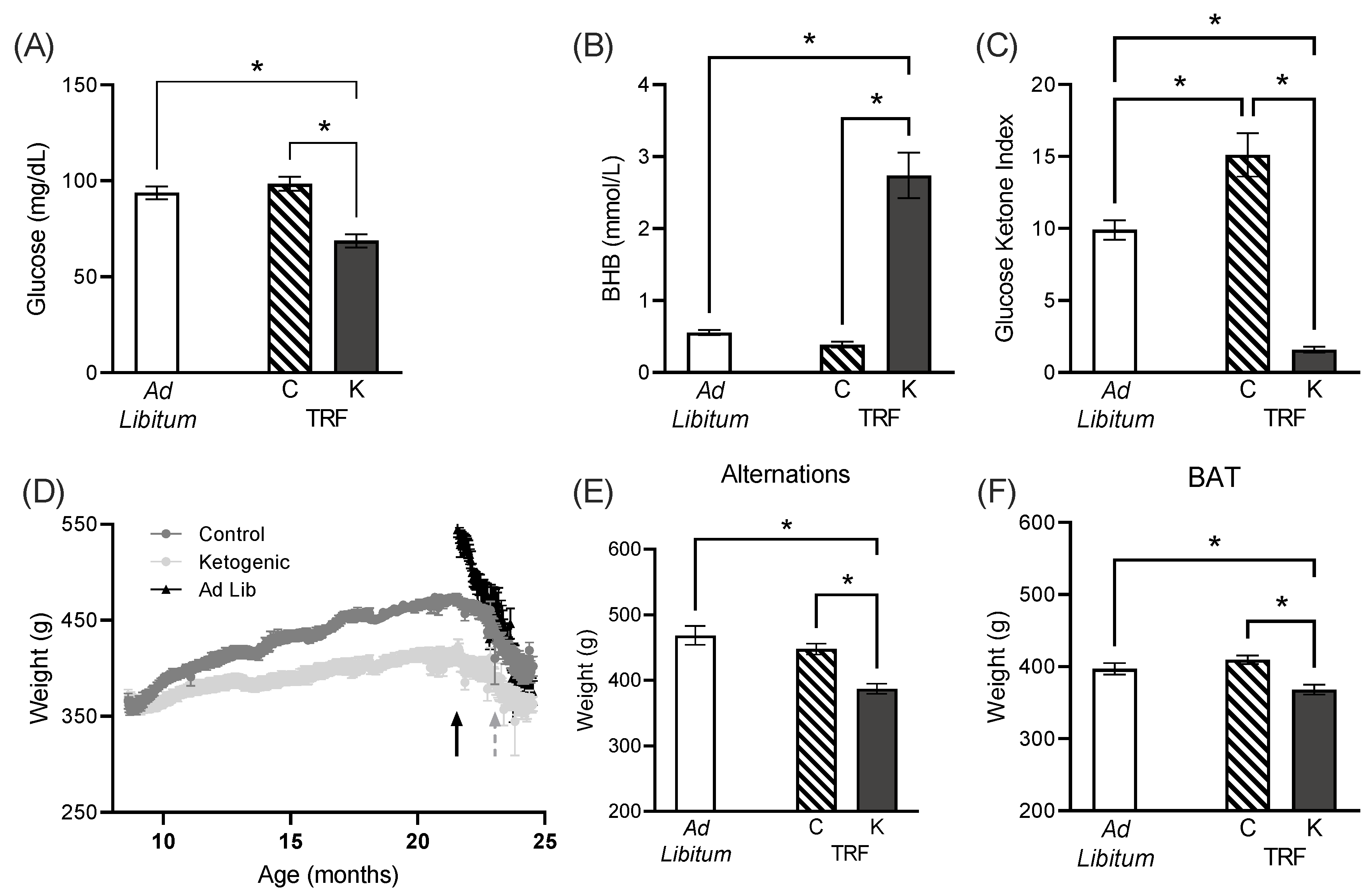
3.1. Peripheral Effects of Feeding Paradigms
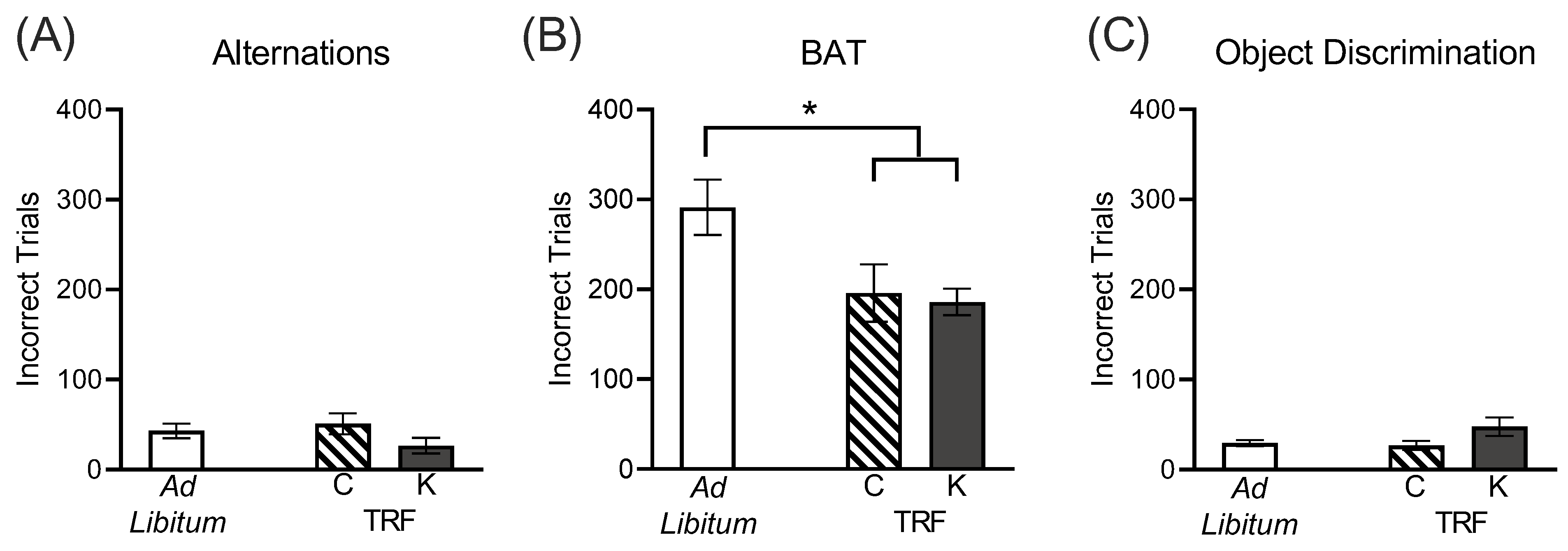
3.2. Time Restricted Feeding, Regardless of Macronutrient Composition, Ameliorated Age-Related Cognitive Impairment on the Biconditional Association Task
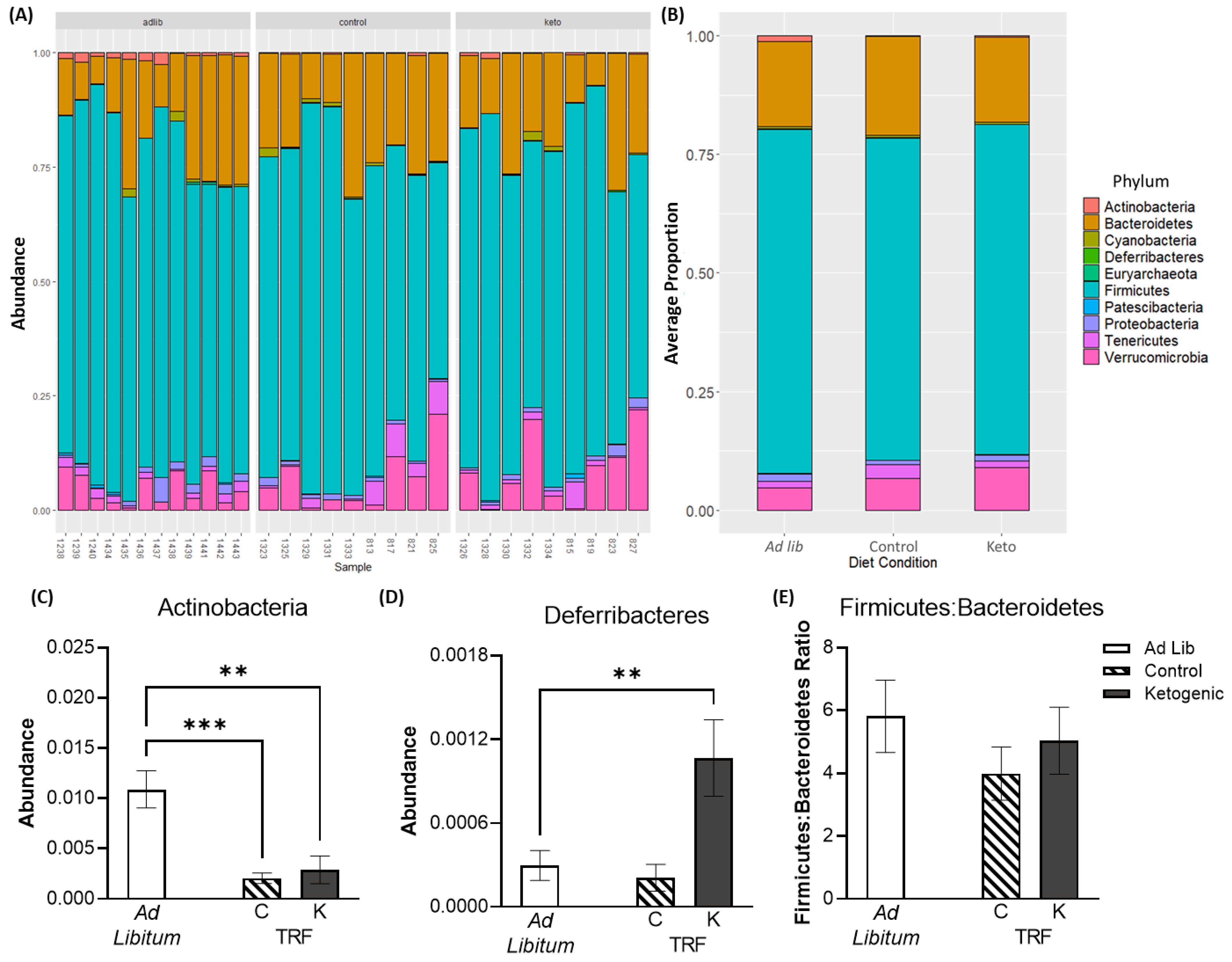
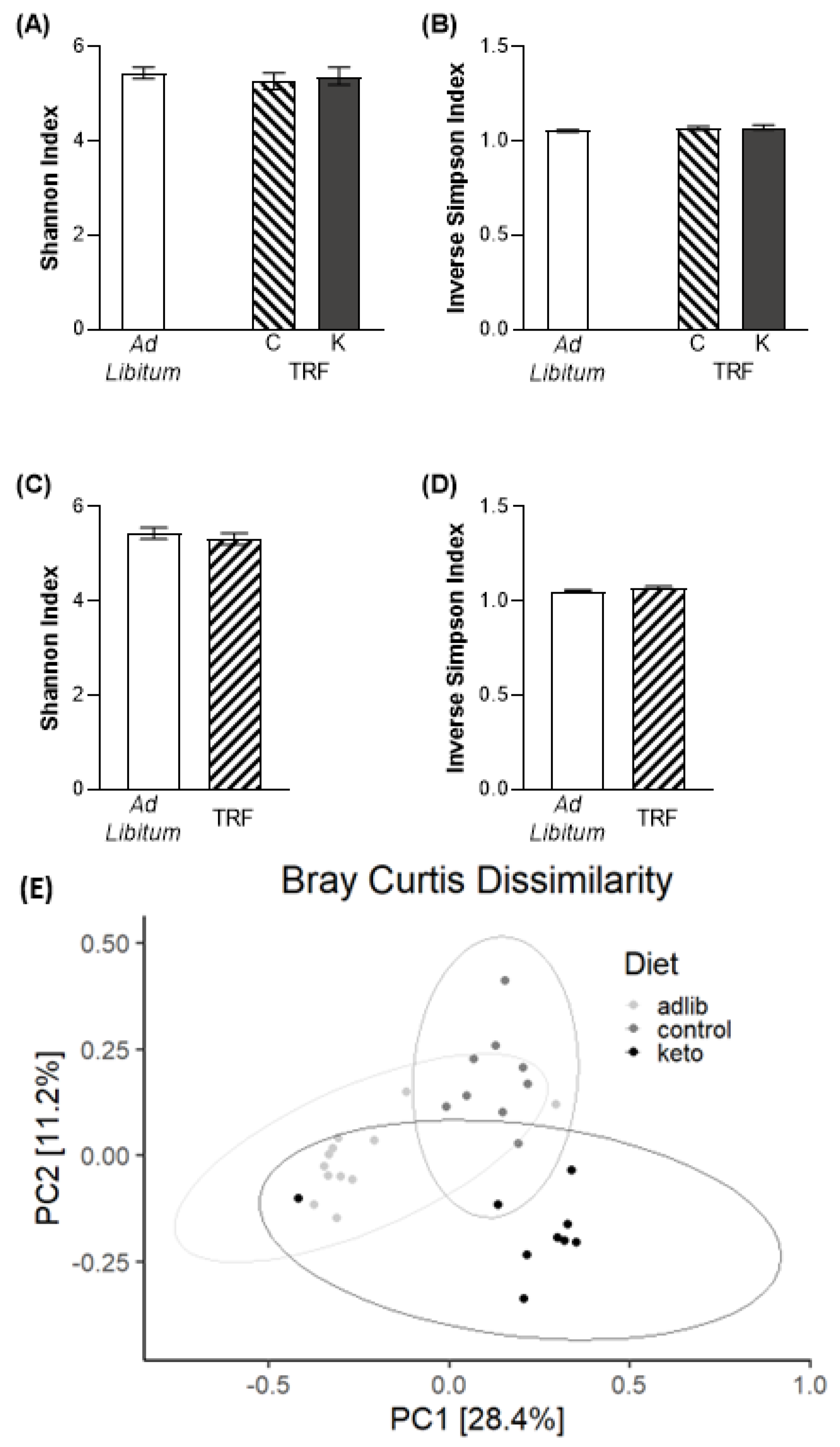
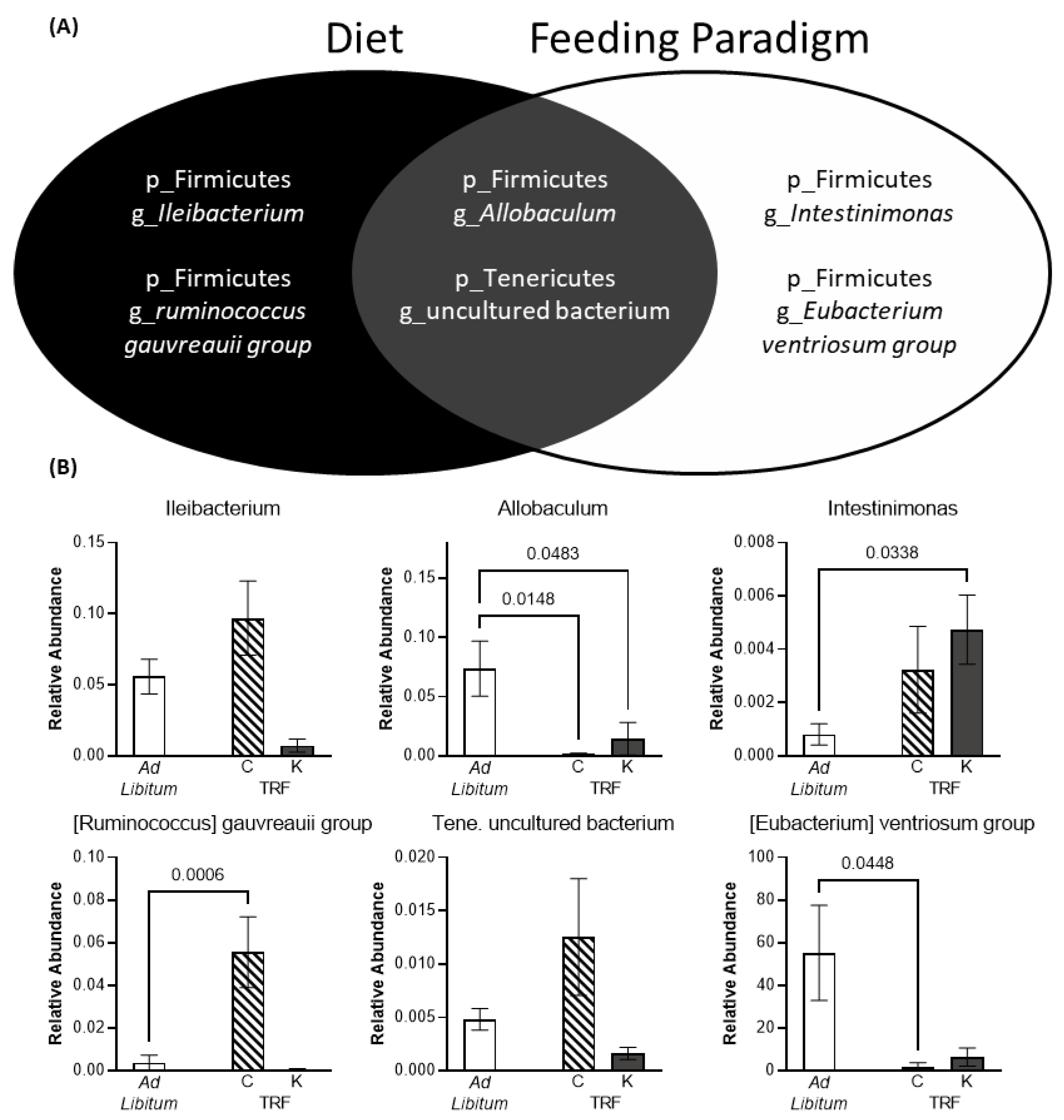
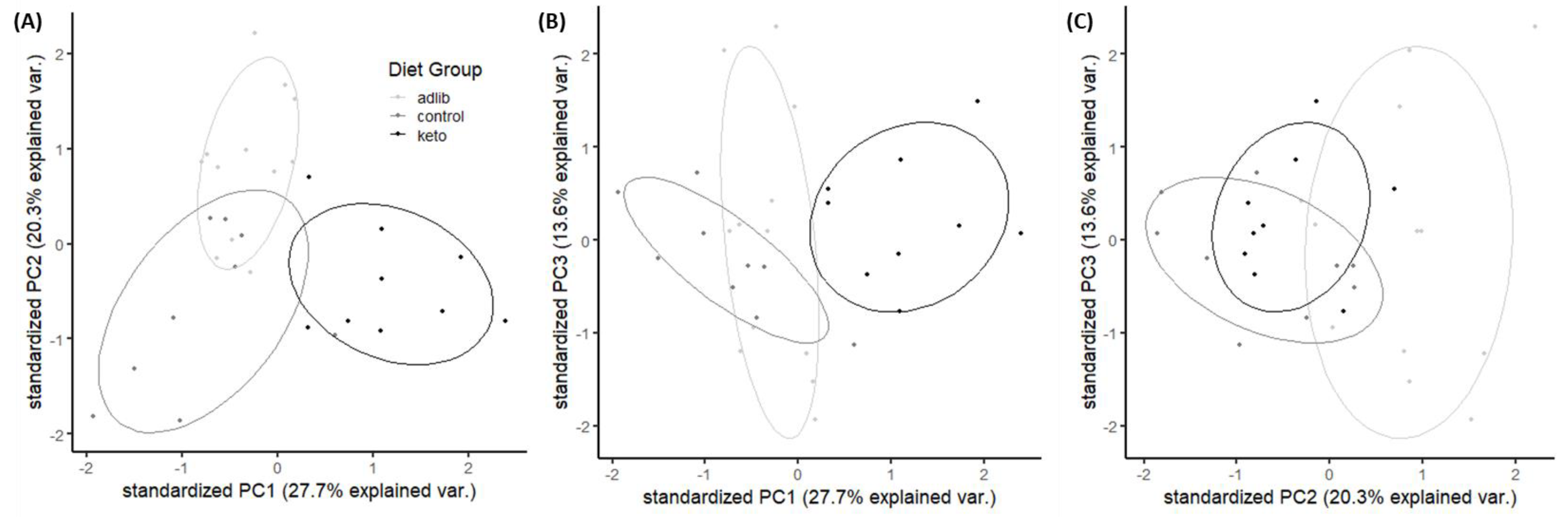
3.3. Time Restricted Feeding and the Ketogenic Diet Influence Gut Microbiome Composition and Beta Diversity
3.4. Alterations in Gut Microbiome Composition Correlate with Behavioral Performance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gold, P.E.; Korol, D.L. Forgetfulness during Aging: An Integrated Biology. Neurobiol. Learn. Mem. 2014, 112, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Trepanowski, J.F.; Canale, R.E.; Marshall, K.E.; Kabir, M.M.; Bloomer, R.J. Impact of Caloric and Dietary Restriction Regimens on Markers of Health and Longevity in Humans and Animals: A Summary of Available Findings. Nutr. J. 2011, 10. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.S.; Leeuwenburgh, C.; Daniels, M.; Foster, T.C. Influence of Calorie Restriction on Measures of Age-Related Cognitive Decline: Role of Increased Physical Activity. J. Gerontol. A. Biol. Sci. Med. Sci. 2009, 64A, 850–859. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Longo, V. Dietary Restriction with and without Caloric Restriction for Healthy Aging. F1000Research 2016, 5, 117. [Google Scholar] [CrossRef]
- Klement, R.J. Mimicking Caloric Restriction: What about Macronutrient Manipulation? A Response to Meynet and Ricci. Trends Mol. Med. 2014, 20, 471–472. [Google Scholar] [CrossRef]
- Newell, C.; Bomhof, M.R.; Reimer, R.A.; Hittel, D.S.; Rho, J.M.; Shearer, J. Ketogenic Diet Modifies the Gut Microbiota in a Murine Model of Autism Spectrum Disorder. Mol. Autism 2016, 7, 37. [Google Scholar] [CrossRef]
- Swidsinski, A.; Dörffel, Y.; Loening-Baucke, V.; Gille, C.; Göktas, Ö.; Reißhauer, A.; Neuhaus, J.; Weylandt, K.-H.; Guschin, A.; Bock, M. Reduced Mass and Diversity of the Colonic Microbiome in Patients with Multiple Sclerosis and Their Improvement with Ketogenic Diet. Front. Microbiol. 2017, 8, 1141. [Google Scholar] [CrossRef]
- Olson, C.A.; Vuong, H.E.; Yano, J.M.; Liang, Q.Y.; Nusbaum, D.J.; Hsiao, E.Y. The Gut Microbiota Mediates the Anti-Seizure Effects of the Ketogenic Diet. Cell 2018, 173, 1728–1741.e13. [Google Scholar] [CrossRef]
- Gutiérrez-Repiso, C.; Hernández-García, C.; García-Almeida, J.M.; Bellido, D.; Martín-Núñez, G.M.; Sánchez-Alcoholado, L.; Alcaide-Torres, J.; Sajoux, I.; Tinahones, F.J.; Moreno-Indias, I. Effect of Synbiotic Supplementation in a Very-Low-Calorie Ketogenic Diet on Weight Loss Achievement and Gut Microbiota: A Randomized Controlled Pilot Study. Mol. Nutr. Food Res. 2019, 63, 1900167. [Google Scholar] [CrossRef]
- Le, H.H.; Johnson, E.L. Going Keto? Say ΒHB-Ye Bye to Your Gut Bifidobacteria. Cell Host Microbe 2020, 28, 3–5. [Google Scholar] [CrossRef]
- Yuan, W.; Lu, W.; Wang, H.; Wu, W.; Zhou, Q.; Chen, Y.; Lee, Y.K.; Zhao, J.; Zhang, H.; Chen, W. A Multiphase Dietetic Protocol Incorporating an Improved Ketogenic Diet Enhances Weight Loss and Alters the Gut Microbiome of Obese People. Int. J. Food Sci. Nutr. 2022, 73, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Zhu, H.; Qiu, P. Aging Progression of Human Gut Microbiota. BMC Microbiol. 2019, 19, 236. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, A.R.; Truckenbrod, L.M.; Federico, Q.P.; Campos, K.T.; Moon, B.M.; Ferekides, N.; Hoppe, M.; D’Agostino, D.; Burke, S.N. Metabolic Switching Is Impaired by Aging and Facilitated by Ketosis Independent of Glycogen. Biochemistry 2019, 12, 7963. [Google Scholar] [CrossRef]
- Kumar, N.K.; Merrill, J.D.; Carlson, S.; German, J.; Yancy, W.S. Adherence to Low-Carbohydrate Diets in Patients with Diabetes: A Narrative Review. Diabetes Metab. Syndr. Obes. Targets Ther. 2022, 15, 477–498. [Google Scholar] [CrossRef]
- Braak, H.; Del Tredici, K. The Preclinical Phase of the Pathological Process Underlying Sporadic Alzheimer’s Disease. Brain 2015, 138, 2814–2833. [Google Scholar] [CrossRef] [PubMed]
- Reiman, E.M.; Chen, K.; Alexander, G.E.; Caselli, R.J.; Bandy, D.; Osborne, D.; Saunders, A.M.; Hardy, J. Functional Brain Abnormalities in Young Adults at Genetic Risk for Late-Onset Alzheimer’s Dementia. Proc. Natl. Acad. Sci. USA 2004, 101, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, A.R.; Hernandez, C.M.; Campos, K.T.; Truckenbrod, L.M.; Sakarya, Y.; McQuail, J.A.; Carter, C.S.; Bizon, J.L.; Maurer, A.P.; Burke, S.N. The Antiepileptic Ketogenic Diet Alters Hippocampal Transporter Levels and Reduces Adiposity in Aged Rats. J. Gerontol. Ser. A 2018, 73, 450–458. [Google Scholar] [CrossRef]
- Hernandez, A.R.; Truckenbrod, L.M.; Campos, K.T.; Williams, S.A.; Burke, S.N. Sex Differences in Age-Related Impairments Vary across Cognitive and Physical Assessments in Rats. Behav. Neurosci. 2020, 134, 69–81. [Google Scholar] [CrossRef]
- Hernandez, A.R.; Maurer, A.P.; Reasor, J.E.; Turner, S.M.; Barthle, S.E.; Johnson, S.A.; Burke, S.N. Age-Related Impairments in Object-Place Associations Are Not Due to Hippocampal Dysfunction. Behav. Neurosci. 2015, 129, 599–610. [Google Scholar] [CrossRef]
- Hernandez, A.R.; Hernandez, C.M.; Campos, K.; Truckenbrod, L.; Federico, Q.; Moon, B.; McQuail, J.A.; Maurer, A.P.; Bizon, J.L.; Burke, S.N. A Ketogenic Diet Improves Cognition and Has Biochemical Effects in Prefrontal Cortex That Are Dissociable From Hippocampus. Front. Aging Neurosci. 2018, 10, 391. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, A.R.; Reasor, J.E.; Truckenbrod, L.M.; Campos, K.T.; Federico, Q.P.; Fertal, K.E.; Lubke, K.N.; Johnson, S.A.; Clark, B.J.; Maurer, A.P.; et al. Dissociable Effects of Advanced Age on Prefrontal Cortical and Medial Temporal Lobe Ensemble Activity. Neurobiol. Aging 2018, 70, 217–232. [Google Scholar] [CrossRef] [PubMed]
- Davidson, G.L.; Cooke, A.C.; Johnson, C.N.; Quinn, J.L. The Gut Microbiome as a Driver of Individual Variation in Cognition and Functional Behaviour. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20170286. [Google Scholar] [CrossRef]
- Gao, W.; Baumgartel, K.L.; Alexander, S.A. The Gut Microbiome as a Component of the Gut–Brain Axis in Cognitive Health. Biol. Res. Nurs. 2020, 22, 485–494. [Google Scholar] [CrossRef]
- Sanz, Y.; Olivares, M.; Moya-Pérez, Á.; Agostoni, C. Understanding the Role of Gut Microbiome in Metabolic Disease Risk. Pediatr. Res. 2015, 77, 236–244. [Google Scholar] [CrossRef] [PubMed]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet Rapidly and Reproducibly Alters the Human Gut Microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef]
- Rosner, B. Fundamentals of Biostatistics, 8th ed.; Cengage Learning: Boston, MA, USA, 2016; ISBN 978-1-305-26892-0. [Google Scholar]
- Hernandez, A.R.; Hernandez, C.M.; Truckenbrod, L.M.; Campos, K.T.; McQuail, J.A.; Bizon, J.L.; Burke, S.N. Age and Ketogenic Diet Have Dissociable Effects on Synapse-Related Gene Expression Between Hippocampal Subregions. Front. Aging Neurosci. 2019, 11, 239. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, A.R.; Reasor, J.E.; Truckenbrod, L.M.; Lubke, K.N.; Johnson, S.A.; Bizon, J.L.; Maurer, A.P.; Burke, S.N. Medial Prefrontal-Perirhinal Cortical Communication Is Necessary for Flexible Response Selection. Neurobiol. Learn. Mem. 2017, 137, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Motulsky, H.J.; Brown, R.E. Detecting Outliers When Fitting Data with Nonlinear Regression–a New Method Based on Robust Nonlinear Regression and the False Discovery Rate. BMC Bioinformatics 2006, 7, 123. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Eipers, P.; Little, R.B.; Crowley, M.; Crossman, D.K.; Lefkowitz, E.J.; Morrow, C.D. Getting Started with Microbiome Analysis: Sample Acquisition to Bioinformatics. Curr. Protoc. Hum. Genet. 2014, 82, 18.8.1–18.8.29. [Google Scholar] [CrossRef] [PubMed]
- Buford, T.W.; Carter, C.S.; VanDerPol, W.J.; Chen, D.; Lefkowitz, E.J.; Eipers, P.; Morrow, C.D.; Bamman, M.M. Composition and Richness of the Serum Microbiome Differ by Age and Link to Systemic Inflammation. GeroScience 2018, 40, 257–268. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, A.R.; Kemp, K.M.; Burke, S.N.; Buford, T.W.; Carter, C.S. Influence of Aging, Macronutrient Composition and Time-Restricted Feeding on the Fischer344 x Brown Norway Rat Gut Microbiota. Nutrients 2022, 14, 1758. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Shetty, S.A.; Lahti, L. Microbiome Data Science. J. Biosci. 2019, 44, 115. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.; Van Treuren, W.; White, R.A.; Eggesbø, M.; Knight, R.; Peddada, S.D. Analysis of Composition of Microbiomes: A Novel Method for Studying Microbial Composition. Microb. Ecol. Health Dis. 2015, 26, 27663. [Google Scholar] [CrossRef] [PubMed]
- Kemp, K.M.; Colson, J.; Lorenz, R.G.; Maynard, C.L.; Pollock, J.S. Early Life Stress in Mice Alters Gut Microbiota Independent of Maternal Microbiota Inheritance. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2021, 320, R663–R674. [Google Scholar] [CrossRef] [PubMed]
- Jena, P.K.; Sheng, L.; Nguyen, M.; Di Lucente, J.; Hu, Y.; Li, Y.; Maezawa, I.; Jin, L.-W.; Wan, Y.-J.Y. Dysregulated Bile Acid Receptor-Mediated Signaling and IL-17A Induction Are Implicated in Diet-Associated Hepatic Health and Cognitive Function. Biomark. Res. 2020, 8, 59. [Google Scholar] [CrossRef] [PubMed]
- Gabel, K.; Marcell, J.; Cares, K.; Kalam, F.; Cienfuegos, S.; Ezpeleta, M.; Varady, K.A. Effect of Time Restricted Feeding on the Gut Microbiome in Adults with Obesity: A Pilot Study. Nutr. Health 2020, 26, 79–85. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Routledge, R.D. Diversity Indices: Which Ones Are Admissible? J. Theor. Biol. 1979, 76, 503–515. [Google Scholar] [CrossRef]
- Meyer, K.; Lulla, A.; Debroy, K.; Shikany, J.M.; Yaffe, K.; Meirelles, O.; Launer, L.J. Association of the Gut Microbiota With Cognitive Function in Midlife. JAMA Netw. Open 2022, 5, e2143941. [Google Scholar] [CrossRef]
- Martin, B.; Ji, S.; Maudsley, S.; Mattson, M.P. “Control” Laboratory Rodents Are Metabolically Morbid: Why It Matters. Proc. Natl. Acad. Sci. USA 2010, 107, 6127–6133. [Google Scholar] [CrossRef] [PubMed]
- Gold, P.E. Glucose and Age-Related Changes in Memory. Neurobiol. Aging 2005, 26, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Ryan, L.; Hay, M.; Huentelman, M.J.; Duarte, A.; Rundek, T.; Levin, B.; Soldan, A.; Pettigrew, C.; Mehl, M.R.; Barnes, C.A. Precision Aging: Applying Precision Medicine to the Field of Cognitive Aging. Front. Aging Neurosci. 2019, 11, 128. [Google Scholar] [CrossRef]
- Zeb, F.; Wu, X.; Chen, L.; Fatima, S.; Haq, I.-U.; Chen, A.; Xu, C.; Jianglei, R.; Feng, Q.; Li, M. Time-Restricted Feeding Is Associated with Changes in Human Gut Microbiota Related to Nutrient Intake. Nutrition 2020, 78, 110797. [Google Scholar] [CrossRef] [PubMed]
- Zeb, F.; Wu, X.; Chen, L.; Fatima, S.; Haq, I.-U.; Chen, A.; Majeed, F.; Feng, Q.; Li, M. Effect of Time-Restricted Feeding on Metabolic Risk and Circadian Rhythm Associated with Gut Microbiome in Healthy Males. Br. J. Nutr. 2020, 123, 1216–1226. [Google Scholar] [CrossRef] [PubMed]
- Janssen, A.W.F.; Katiraei, S.; Bartosinska, B.; Eberhard, D.; Willems van Dijk, K.; Kersten, S. Loss of Angiopoietin-like 4 (ANGPTL4) in Mice with Diet-Induced Obesity Uncouples Visceral Obesity from Glucose Intolerance Partly via the Gut Microbiota. Diabetologia 2018, 61, 1447–1458. [Google Scholar] [CrossRef]
- Li, S.; Qi, Y.; Ren, D.; Qu, D.; Sun, Y. The Structure Features and Improving Effects of Polysaccharide from Astragalus Membranaceus on Antibiotic-Associated Diarrhea. Antibiot. Basel Switz. 2019, 9, 8. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Lyu, W.; Ren, Y.; Li, X.; Zhao, S.; Yang, H.; Xiao, Y. Allobaculum Involves in the Modulation of Intestinal ANGPTLT4 Expression in Mice Treated by High-Fat Diet. Front. Nutr. 2021, 8, 242. [Google Scholar] [CrossRef] [PubMed]
- Jiao, X.; Wang, Y.; Lin, Y.; Lang, Y.; Li, E.; Zhang, X.; Zhang, Q.; Feng, Y.; Meng, X.; Li, B. Blueberry Polyphenols Extract as a Potential Prebiotic with Anti-Obesity Effects on C57BL/6 J Mice by Modulating the Gut Microbiota. J. Nutr. Biochem. 2019, 64, 88–100. [Google Scholar] [CrossRef]
- Wang, G.; Zhou, H.-H.; Luo, L.; Qin, L.-Q.; Yin, J.; Yu, Z.; Zhang, L.; Wan, Z. Voluntary Wheel Running Is Capable of Improving Cognitive Function Only in the Young but Not the Middle-Aged Male APPSwe/PS1De9 Mice. Neurochem. Int. 2021, 145, 105010. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Li, R.; Wang, G.; Chen, J.; Chen, L.; Qin, L.-Q.; Yu, Z.; Wan, Z. Age-Dependent Effects of a High-Fat Diet Combined with Dietary Advanced Glycation End Products on Cognitive Function and Protection with Voluntary Exercise. Food Funct. 2022, 13, 4445–4458. [Google Scholar] [CrossRef] [PubMed]
- Beilharz, J.; Maniam, J.; Morris, M. Diet-Induced Cognitive Deficits: The Role of Fat and Sugar, Potential Mechanisms and Nutritional Interventions. Nutrients 2015, 7, 6719–6738. [Google Scholar] [CrossRef] [PubMed]
- U.S Department of Health and Human Services. National Diabetes Statistics Report 2020. Available online: chrome-extension://efaidnbmnnnibpcajpcglclefindmkaj/https://oversight.house.gov/sites/democrats.oversight.house.gov/files/2022.09.14%20FINAL%20COR%20Supplemental%20Memo.pdf (accessed on 13 August 2022).
- Liu, Z.; Dai, X.; Zhang, H.; Shi, R.; Hui, Y.; Jin, X.; Zhang, W.; Wang, L.; Wang, Q.; Wang, D.; et al. Gut Microbiota Mediates Intermittent-Fasting Alleviation of Diabetes-Induced Cognitive Impairment. Nat. Commun. 2020, 11, 855. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernandez, A.R.; Watson, C.; Federico, Q.P.; Fletcher, R.; Brotgandel, A.; Buford, T.W.; Carter, C.S.; Burke, S.N. Twelve Months of Time-Restricted Feeding Improves Cognition and Alters Microbiome Composition Independent of Macronutrient Composition. Nutrients 2022, 14, 3977. https://doi.org/10.3390/nu14193977
Hernandez AR, Watson C, Federico QP, Fletcher R, Brotgandel A, Buford TW, Carter CS, Burke SN. Twelve Months of Time-Restricted Feeding Improves Cognition and Alters Microbiome Composition Independent of Macronutrient Composition. Nutrients. 2022; 14(19):3977. https://doi.org/10.3390/nu14193977
Chicago/Turabian StyleHernandez, Abbi R., Cory Watson, Quinten P. Federico, Rachel Fletcher, Armen Brotgandel, Thomas W. Buford, Christy S. Carter, and Sara N. Burke. 2022. "Twelve Months of Time-Restricted Feeding Improves Cognition and Alters Microbiome Composition Independent of Macronutrient Composition" Nutrients 14, no. 19: 3977. https://doi.org/10.3390/nu14193977