
Supplementation with Queen Bee Larva Powder Extended the Longevity of Caenorhabditis elegans
Abstract
:1. Introduction
2. Materials and Methods
2.1. C. elegans Strains, Growth Conditions, and QBLP Treatment
2.2. C. elegans Lifespan Assay
2.3. C. elegans Locomotion Assay
2.4. RNA Extraction and Transcriptome Analysis
2.5. Functional Analyses
2.6. Quantitative Real-Time PCR Validation
2.7. Statistical Analysis
3. Results
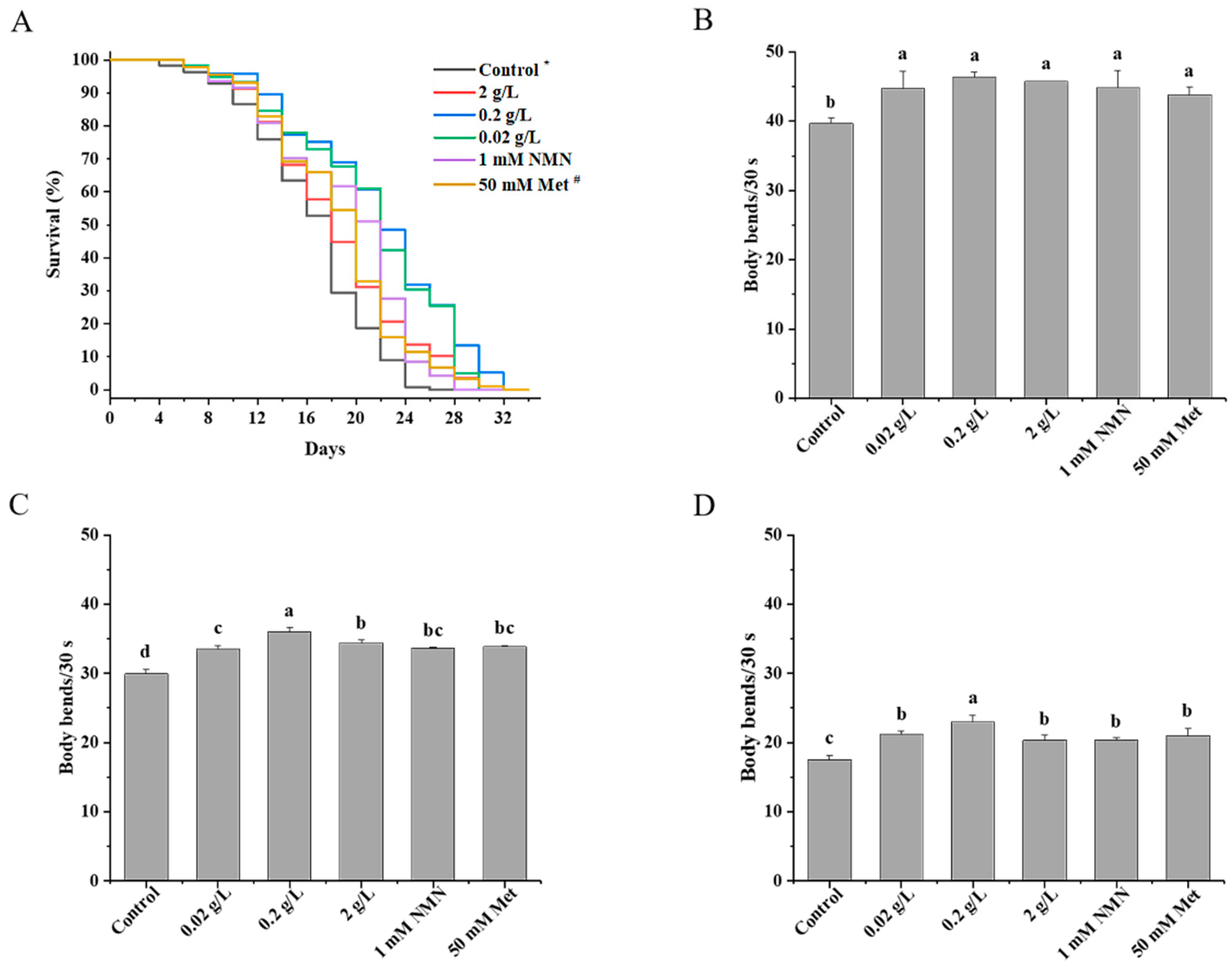
3.1. Effects of QBLP Supplementation on the Lifespan of C. elegans
3.2. Effects of QBLP Supplementation on the Locomotion of C. elegans
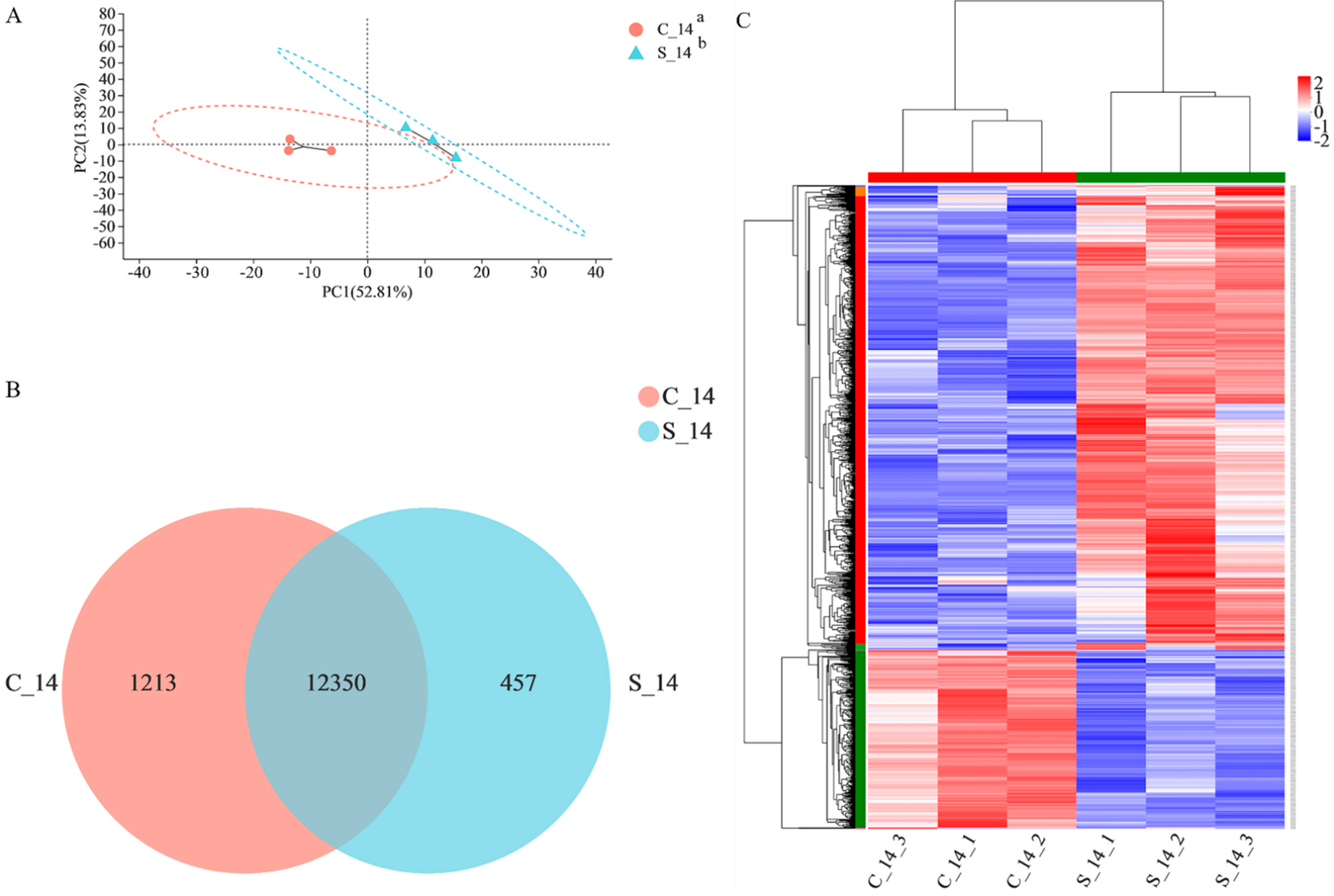
3.3. Overview of RNA-Sequencing
3.4. Differential Expression mRNA Analysis
3.5. Gene Ontology Functional Analysis of mRNAs
3.6. KEGG Pathway Analysis of mRNAs
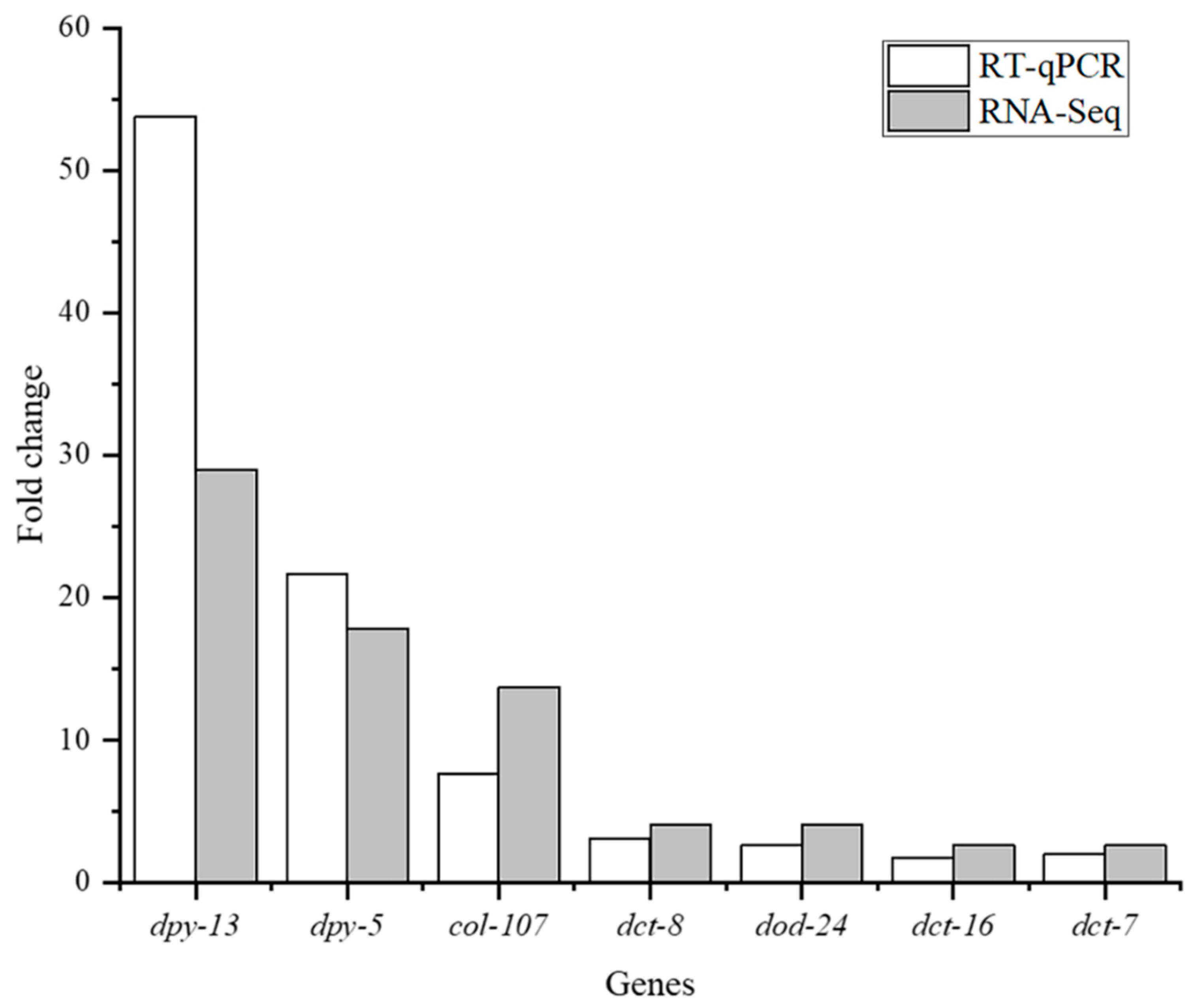
3.7. Validation of mRNA Expression
3.8. Effects of QBLP Supplementation on the Lifespan of daf-16 Mutants
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Culig, L.; Chu, X.; Bohr, V.A. Neurogenesis in aging and age-related neurodegenerative diseases. Ageing Res. Rev. 2022, 78, 101636. [Google Scholar] [CrossRef] [PubMed]
- Kritsilis, M.; Rizou, S.V.; Koutsoudaki, P.N.; Evangelou, K.; Gorgoulis, V.G.; Papadopoulos, D. Ageing, Cellular Senescence and Neurodegenerative Disease. Int. J. Mol. Sci. 2018, 19, 2937. [Google Scholar] [CrossRef] [PubMed]
- Nadeeshani, H.; Li, J.Y.; Ying, T.L.; Zhang, B.H.; Lu, J. Nicotinamide mononucleotide (NMN) as an anti-aging health product-Promises and safety concerns. J. Adv. Res. 2022, 37, 267–278. [Google Scholar] [CrossRef]
- United Nations: Global Issues, Ageing. 2019. Available online: https://www.un.org/en/sections/issues-depth/ageing/ (accessed on 10 May 2022).
- Li, Z.H.; Cheng, J.; Huang, L.; Li, W.X.; Zhao, Y.P.; Lin, W.Y. Aging Diagnostic Probe for Research on Aging and Evaluation of Anti-aging Drug Efficacy. Anal. Chem. 2021, 93, 13800–13806. [Google Scholar] [CrossRef] [PubMed]
- Dong, D.; Dong, M.; Liu, K.; Yu, B. Antioxidant activity of queen bee larvae processed by enzymatic hydrolysis. J. Food Processing Preserv. 2018, 42, e13461. [Google Scholar] [CrossRef]
- Hartfelder, K.; Guidugli-Lazzarini, K.R.; Cervoni, M.S.; Santos, D.E.; Humann, F.C. Old threads make new tapestry-rewiring of signalling pathways underlies caste phenotypic plasticity in the honey bee, Apis mellifera L. Adv. Insect Physiol. 2015, 48, 1–36. [Google Scholar] [CrossRef]
- Tang, Q.H.; Xiong, J.; Wang, J.X.; Cao, Z.; Liao, S.Q.; Xiao, Y.; Tian, W.L.; Guo, J. Queen bee larva consumption improves sleep disorder and regulates gut microbiota in mice with PCPA-induced insomnia. Food Biosci. 2021, 43, 101256. [Google Scholar] [CrossRef]
- Haber, M.; Mishyna, M.; Martinez, J.J.I.; Benjamin, O. Edible larvae and pupae of honey bee (Apis mellifera): Odor and nutritional characterization as a function of diet. Food Chem. 2019, 292, 197–203. [Google Scholar] [CrossRef]
- Wu, X.L.; He, K.; Velickovic, T.C.; Liu, Z.G. Nutritional, functional, and allergenic properties of silkworm pupae. Food Sci. Nutr. 2021, 9, 4655–4665. [Google Scholar] [CrossRef]
- Gao, Y.; Zhao, Y.J.; Xu, M.L.; Shi, S.S. Clanis bilineata tsingtauica: A Sustainable Edible Insect Resource. Sustainability 2021, 13, 12533. [Google Scholar] [CrossRef]
- del Hierro, J.N.; Gutierrez-Docio, A.; Otero, P.; Reglero, G.; Martin, D. Characterization, antioxidant activity, and inhibitory effect on pancreatic lipase of extracts from the edible insects Acheta domesticus and Tenebrio molitor. Food Chem. 2020, 309, 125742. [Google Scholar] [CrossRef] [PubMed]
- Hall, F.; Johnson, P.E.; Liceaga, A. Effect of enzymatic hydrolysis on bioactive properties and allergenicity of cricket (Gryllodes sigillatus) protein. Food Chem. 2018, 262, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.D.; Min, H.J.; Won, Y.S.; Ahn, H.Y.; Cho, Y.S.; Seo, K.I. Solid state fermentation process with Aspergillus kawachii enhances the cancer-suppressive potential of silkworm larva in hepatocellular carcinoma cells. BMC Complementary Altern. Med. 2019, 19, 241. [Google Scholar] [CrossRef]
- Seo, M.; Goo, T.W.; Chung, M.Y.; Baek, M.; Hwang, J.S.; Kim, M.A.; Yun, E.Y. Tenebrio molitor Larvae Inhibit Adipogenesis through AMPK and MAPKs Signaling in 3T3-L1 Adipocytes and Obesity in High-Fat Diet-Induced Obese Mice. Int. J. Mol. Sci. 2017, 18, 518. [Google Scholar] [CrossRef]
- Lacroix, I.M.E.; Teran, I.D.; Fogliano, V.; Wichers, H.J. Investigation into the potential of commercially available lesser mealworm (A. diaperinus) protein to serve as sources of peptides with DPP-IV inhibitory activity. Int. J. Food Sci. Technol. 2019, 54, 696–704. [Google Scholar] [CrossRef]
- Reigada, I.; Kapp, K.; Maynard, C.; Weinkove, D.; Valero, M.S.; Langa, E.; Hanski, L.; Gomez-Rincon, C. Alterations in Bacterial Metabolism Contribute to the Lifespan Extension Exerted by Guarana in Caenorhabditis elegans. Nutrients 2022, 14, 1986. [Google Scholar] [CrossRef]
- Liao, V.H.C. Use of Caenorhabditis elegans To Study the Potential Bioactivity of Natural Compounds. J. Agric. Food Chem. 2018, 66, 1737–1742. [Google Scholar] [CrossRef]
- Li, S.T.; Zhao, H.Q.; Zhang, P.; Liang, C.Y.; Zhang, Y.P.; Hsu, A.L.; Dong, M.Q. DAF-16 stabilizes the aging transcriptome and is activated in mid-aged Caenorhabditis elegans to cope with internal stress. Aging Cell 2019, 18, e12896. [Google Scholar] [CrossRef]
- Ye, Y.L.; Gu, Q.Y.; Sun, X.L. Potential of Caenorhabditis elegansas an antiaging evaluation model for dietary phytochemicals: A review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 3084–3105. [Google Scholar] [CrossRef]
- Peng, Y.; Sun, Q.C.; Gao, R.C.; Park, Y. AAK-2 and SKN-1 Are Involved in Chicoric-Acid-Induced Lifespan Extension in Caenorhabditis elegans. J. Agric. Food Chem. 2019, 67, 9178–9186. [Google Scholar] [CrossRef]
- Lopez-Garcia, G.; Cilla, A.; Barbera, R.; Genoves, S.; Martorell, P.; Alegria, A. Effect of plant sterol and galactooligosaccharides enriched beverages on oxidative stress and longevity in Caenorhabditis elegans. J. Funct. Foods 2020, 65, 103747. [Google Scholar] [CrossRef]
- Deng, Y.F.; Liu, H.; Huang, Q.; Tu, L.Y.; Hu, L.; Zheng, B.S.; Sun, H.Q.; Lu, D.J.; Guo, C.W.; Zhou, L. Mechanism of Longevity Extension of Caenorhabditis elegans Induced by Schizophyllum commune Fermented Supernatant With Added Radix Puerariae. Front. Nutr. 2022, 9, 847064. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, Y.; Gan, Q.W.; Xu, M.; Ding, X.; Tang, G.H.; Liang, J.J.; Liu, K.; Liu, X.Z.; Wang, X.; et al. C. elegans-based screen identifies lysosome-damaging alkaloids that induce STAT3-dependent lysosomal cell death. Protein Cell 2018, 9, 1013–1026. [Google Scholar] [CrossRef]
- Li, P.; Wang, Z.H.; Lam, S.M.; Shui, G.H. Rebaudioside A Enhances Resistance to Oxidative Stress and Extends Lifespan and Healthspan in Caenorhabditis elegans. Antioxidants 2021, 10, 262. [Google Scholar] [CrossRef] [PubMed]
- Fang, E.F.; Waltz, T.B.; Kassahun, H.; Lu, Q.P.; Kerr, J.S.; Morevati, M.; Fivenson, E.M.; Wollman, B.N.; Marosi, K.; Wilson, M.A.; et al. Tomatidine enhances lifespan and healthspan in C. elegans through mitophagy induction via the SKN-1/Nrf2 pathway. Sci. Rep. 2017, 7, 46208. [Google Scholar] [CrossRef] [PubMed]
- Bansal, A.; Zhu, L.H.J.; Yen, K.; Tissenbaum, H.A. Uncoupling lifespan and healthspan in Caenorhabditis elegans longevity mutants. Proc. Natl. Acad. Sci. USA 2015, 112, E277–E286. [Google Scholar] [CrossRef] [PubMed]
- Serna, E.; Mastaloudis, A.; Martorell, P.; Wood, S.M.; Hester, S.N.; Bartlett, M.; Prolla, T.A.; Vina, J. A Novel Micronutrient Blend Mimics Calorie Restriction Transcriptomics in Multiple Tissues of Mice and Increases Lifespan and Mobility in C. elegans. Nutrients 2020, 12, 486. [Google Scholar] [CrossRef]
- Park, S.; Kim, B.K.; Park, S.K. Supplementation with phosphatidylethanolamine confers anti-oxidant and anti-aging effects via hormesis and reduced insulin/IGF-1-like signaling in C. elegans. Mech. Ageing Dev. 2021, 197, 111498. [Google Scholar] [CrossRef]
- Kim, H.M.; Long, N.P.; Yoon, S.J.; Nguyen, H.T.; Kwon, S.W. Metabolomics and phenotype assessment reveal cellular toxicity of triclosan in Caenorhabditis elegans. Chemosphere 2019, 236, 124306. [Google Scholar] [CrossRef]
- Machalinski, B.; Roginska, D.; Wilk, A.; Szumilas, K.; Prowans, P.; Paczkowska, E.; Szumilas, P.; Stecewicz, I.; Zawodny, P.; Zietek, M.; et al. Global Gene Expression of Cultured Human Dermal Fibroblasts: Focus on Cell Cycle and Proliferation Status in Improving the Condition of Face Skin. Int. J. Med. Sci. 2021, 18, 1519–1531. [Google Scholar] [CrossRef]
- Guha, S.; Cao, M.; Kane, R.M.; Savino, A.M.; Zou, S.G.; Dong, Y.Q. The longevity effect of cranberry extract in Caenorhabditis elegans is modulated by daf-16 and osr-1. Age 2013, 35, 1559–1574. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.X.; Cook, L.F.; Grasso, L.M.; Cao, M.; Dong, Y.Q. Royal jelly-mediated prolongevity and stress resistance in Caenorhabditis elegans is possibly modulated by the interplays of DAF-16, SIR-2.1, HCF-1, and 14-3-3 proteins. J. Gerontol. Biol. Sci. 2015, 70, 827–838. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.L.; Lui, E.Y.; Ip, Y.K.; Lam, S.H. RNA sequencing, de novo assembly and differential analysis of the gill transcriptome of freshwater climbing perch Anabas testudineus after 6 days of seawater exposure. J. Fish Biol. 2018, 93, 215–228. [Google Scholar] [CrossRef] [PubMed]
- Herndon, L.A.; Schmeissner, P.J.; Dudaronek, J.M.; Brown, P.A.; Listner, K.M.; Sakano, Y.; Paupard, M.C.; Hall, D.H.; Driscoll, M. Stochastic and genetic factors influence tissue-specific decline in aging C. elegans. Nature 2002, 419, 808–814. [Google Scholar] [CrossRef]
- Sun, N.; Youle, R.J.; Finkel, T. The Mitochondrial Basis of Aging. Mol. Cell 2016, 61, 654–666. [Google Scholar] [CrossRef]
- O’Rourke, E.J.; Kuballa, P.; Xavier, R.; Ruvkun, G. omega-6 Polyunsaturated fatty acids extend life span through the activation of autophagy. Genes Dev. 2013, 27, 429–440. [Google Scholar] [CrossRef]
- Wang, C.; Li, Y.Y.; Zeng, L.J.; Shi, C.L.; Peng, Y.; Li, H.; Chen, H.B.; Yu, J.; Zhang, J.; Cheng, B.; et al. Tris(1,3-dichloro-2-propyl) phosphate reduces longevity through a specific microRNA-mediated DAF-16/FoxO in an unconventional insulin/ insulin-like growth factor-1 signaling pathway. J. Hazard. Mater. 2022, 425, 128034. [Google Scholar] [CrossRef]
- Scerbak, C.; Vayndorf, E.; Hernandez, A.; McGill, C.; Taylor, B. Lowbush cranberry acts through DAF-16/FOXO signaling to promote increased lifespan and axon branching in aging posterior touch receptor neurons. Geroscience 2018, 40, 151–162. [Google Scholar] [CrossRef]
- Chen, Q.; Xu, B.J.; Huang, W.S.; Amrouche, A.T.; Maurizio, B.; Simal-Gandara, J.; Tundis, R.; Xiao, J.B.; Zou, L.; Lu, B.Y. Edible flowers as functional raw materials: A review on anti-aging properties. Trends Food Sci. Technol. 2020, 106, 30–47. [Google Scholar] [CrossRef]
- Luyten, W.; Antal, P.; Braeckman, B.P.; Bundy, J.; Cirulli, F.; Fang-Yen, C.; Fuellen, G.; Leroi, A.; Liu, Q.; Martorell, P. Ageing with elegans: A research proposal to map healthspan pathways. Biogerontology 2016, 17, 771–782. [Google Scholar] [CrossRef] [Green Version]
- Shen, P.Y.; Yue, Y.R.; Zheng, J.; Park, Y. Caenorhabditis elegans: A Convenient In Vivo Model for Assessing the Impact of Food Bioactive Compounds on Obesity, Aging, and Alzheimer’s Disease. Annu. Rev. Food Sci. Technol. 2018, 9, 1–22. [Google Scholar] [CrossRef]
- Zheng, S.Q.; Liao, S.T.; Zou, Y.X.; Qu, Z.; Liu, F. ins-7 Gene Expression Is Partially Regulated by the DAF-16/IIS Signaling Pathway in Caenorhabditis elegans under Celecoxib Intervention. PLoS ONE 2014, 9, e100320. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Liu, J.; Bu, L.L.; Liao, D.F.; Cheng, S.W.; Zheng, X.L. Curcumin Acetylsalicylate Extends the Lifespan of Caenorhabditis elegans. Molecules 2021, 26, 6609. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, T.; Sakamoto, K. Killed Bifidobacterium longum enhanced stress tolerance and prolonged life span of Caenorhabditis elegans via DAF-16. Br. J. Nutr. 2018, 120, 872–880. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Whetstine, J.R.; Ghosh, S.; Hanover, J.A.; Gali, R.R.; Grosu, P.; Shi, Y. The conserved NAD(H)-dependent corepressor CTBP-1 regulates Caenorhabditis elegans life span. Proc. Natl. Acad. Sci. USA 2009, 106, 1496–1501. [Google Scholar] [CrossRef] [PubMed]
- Elle, I.C.; Olsen, L.C.B.; Pultz, D.; Rodkaer, S.V.; Faergeman, N.J. Something worth dying for: Molecular tools for the dissection of lipid metabolism in Caenorhabditis elegans. FEBS Lett. 2010, 584, 2183–2193. [Google Scholar] [CrossRef]
- Hansen, M.; Flatt, T.; Aguilaniu, H. Reproduction, Fat Metabolism, and Life Span: What Is the Connection? Cell Metab. 2013, 17, 10–19. [Google Scholar] [CrossRef]
- McCormick, M.; Chen, K.; Ramaswamy, P.; Kenyon, C. New genes that extend Caenorhabditis elegans’ lifespan in response to reproductive signals. Aging Cell 2012, 11, 192–202. [Google Scholar] [CrossRef]
- Matsuoka, T.; Kawashima, T.; Nakamura, T.; Kanamaru, Y.; Yabe, T. Isolation and characterization of proteases that hydrolyze royal jelly proteins from queen bee larvae of the honeybee, Apis mellifera. Apidologie 2012, 43, 685–697. [Google Scholar] [CrossRef]
- Wang, X.X.; Cao, M.; Dong, Y.Q. Royal jelly promotes DAF-16-mediated proteostasis to tolerate β-amyloid toxicity in C. elegans model of Alzheimer’s disease. Oncotarget 2016, 7, 54183–54193. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.N.; Li, M.J.; Zhao, D.F.; Li, X.; Yang, C.L.; Wang, X.C. Lysosome activity is modulated by multiple longevity pathways and is important for lifespan extension in C. elegans. eLife 2020, 9, e55745. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Xu, X.R.; Niklason, L.E. Bmk-1 regulates lifespan in Caenorhabditis elegans by activating hsp-16. Oncotarget 2015, 6, 18790–18799. [Google Scholar] [CrossRef] [PubMed]
- Hars, E.S.; Qi, H.Y.; Ryazanov, A.G.; Jin, S.K.; Cai, L.; Hu, C.C.; Liu, L.F. Autophagy regulates ageing in C-elegans. Autophagy 2007, 3, 93–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | Number of Animals that Died/Total | Mean Lifespan (Days ± SEM) | Median Lifespan (Days ± SEM) | Change from Control (d) | Increase versus Control (%) |
| Control a | 112/120 | 18.5 ± 0.5 | 20.0 ± 0.6 | / | / |
| 0.02 g/L | 116/120 | 20.3 ± 0.5 ** | 20.0 ± 0.7 | 1.8 | 9.7 |
| 0.2 g/L | 97/120 | 23.7 ± 0.7 *** | 24.0 ± 0.7 | 5.2 | 28.1 |
| 2 g/L | 118/120 | 23.1 ± 0.6 *** | 24.0 ± 0.5 | 4.6 | 24.9 |
| 1 mM NMN | 94/120 | 21.1 ± 0.6 *** | 24.0 ± 0.5 | 2.6 | 14.1 |
| 50 mM Met b | 88/120 | 20.6 ± 0.6 *** | 22.0 ± 0.6 | 2.1 | 11.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, T.; Wu, L.; Fan, F.; Yang, Y.; Xue, X. Supplementation with Queen Bee Larva Powder Extended the Longevity of Caenorhabditis elegans. Nutrients 2022, 14, 3976. https://doi.org/10.3390/nu14193976
Zhao T, Wu L, Fan F, Yang Y, Xue X. Supplementation with Queen Bee Larva Powder Extended the Longevity of Caenorhabditis elegans. Nutrients. 2022; 14(19):3976. https://doi.org/10.3390/nu14193976
Chicago/Turabian StyleZhao, Tong, Liming Wu, Fangfang Fan, Yaning Yang, and Xiaofeng Xue. 2022. "Supplementation with Queen Bee Larva Powder Extended the Longevity of Caenorhabditis elegans" Nutrients 14, no. 19: 3976. https://doi.org/10.3390/nu14193976