

Correlation between PPARG Pro12Ala Polymorphism and Therapeutic Responses to Thiazolidinediones in Patients with Type 2 Diabetes: A Meta-Analysis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Protocol and Registration
2.2. Search Strategy and Study Selection
2.3. Data Extraction and Quality Assessment
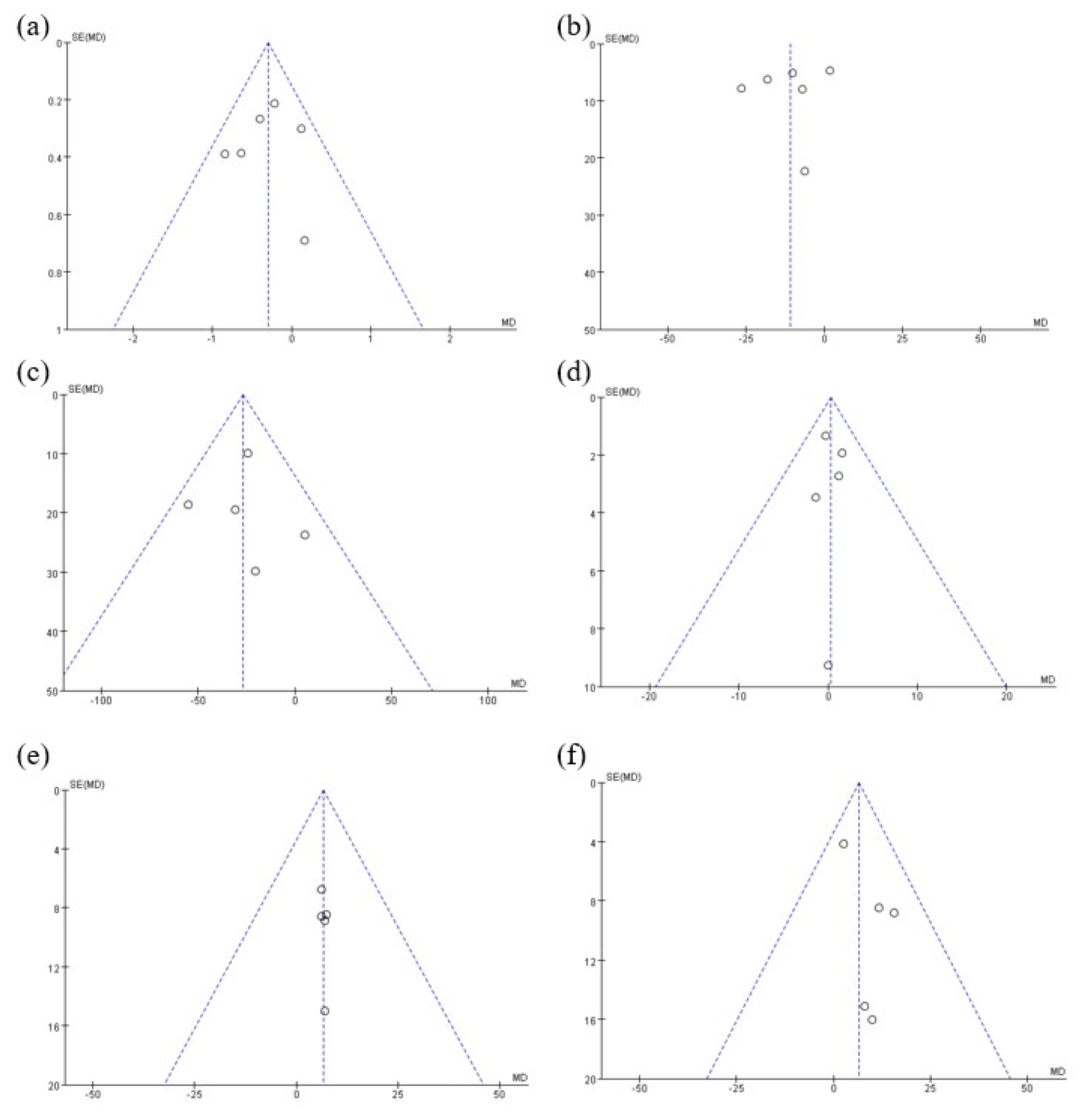
2.4. Statistical Analysis
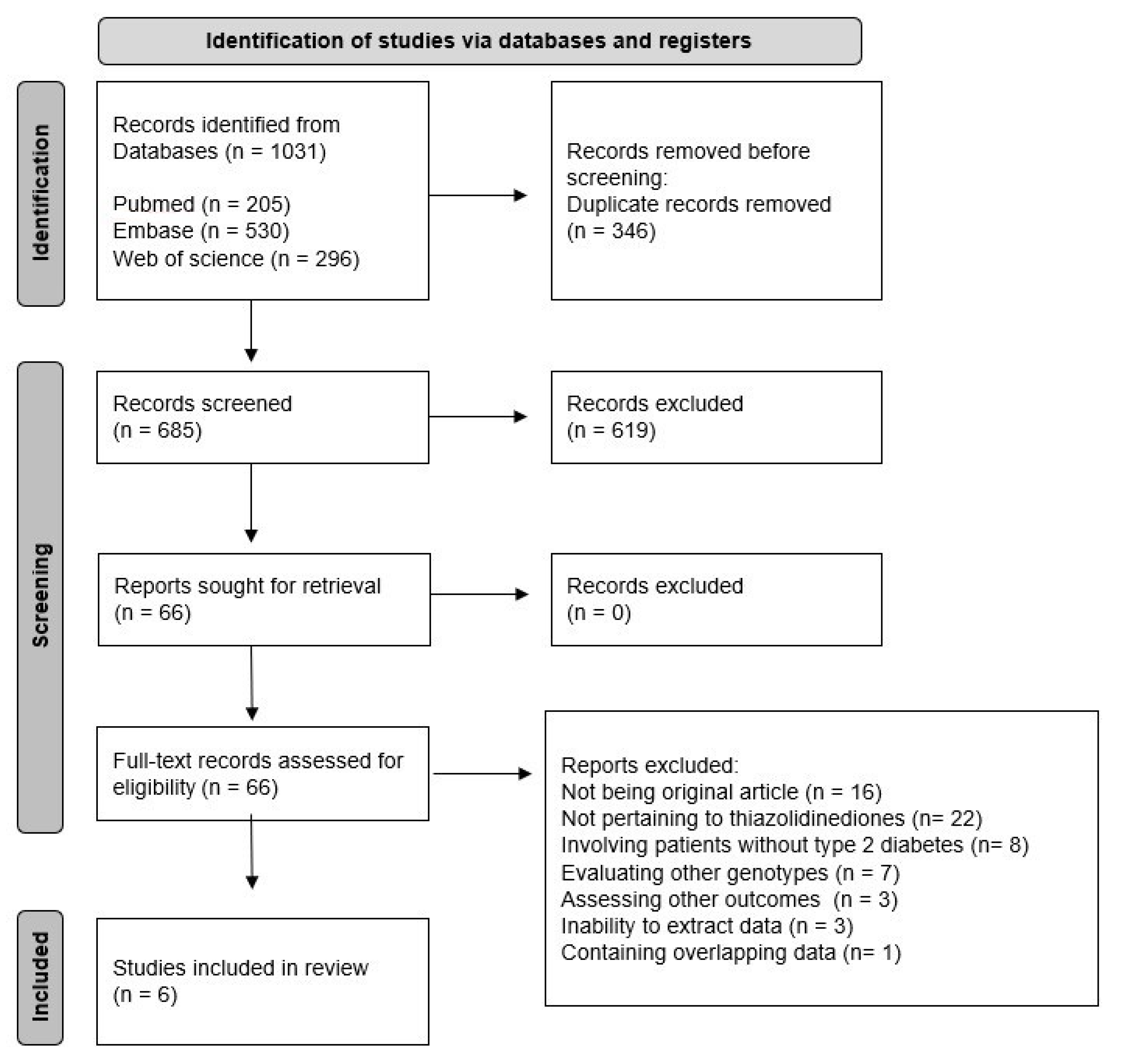
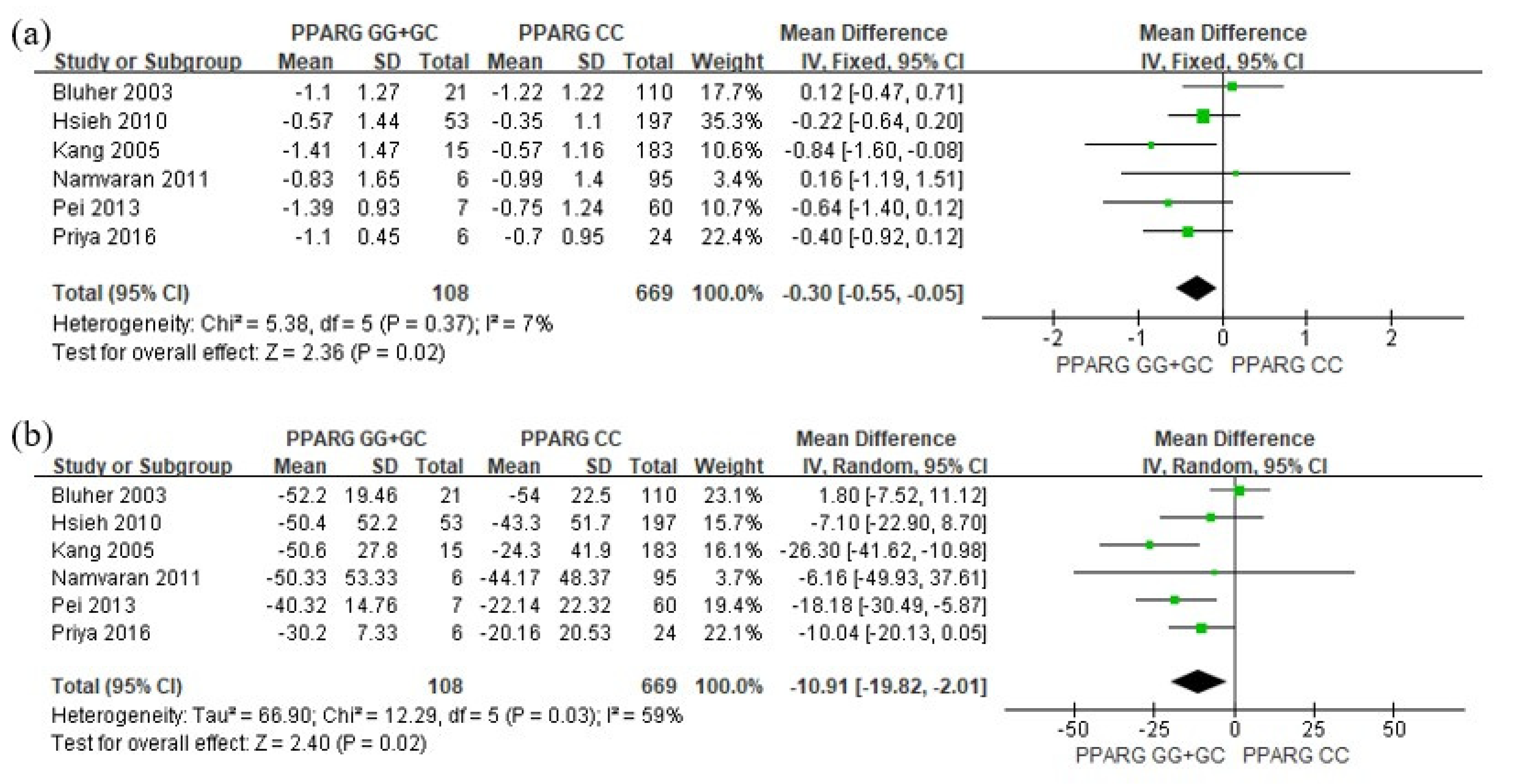
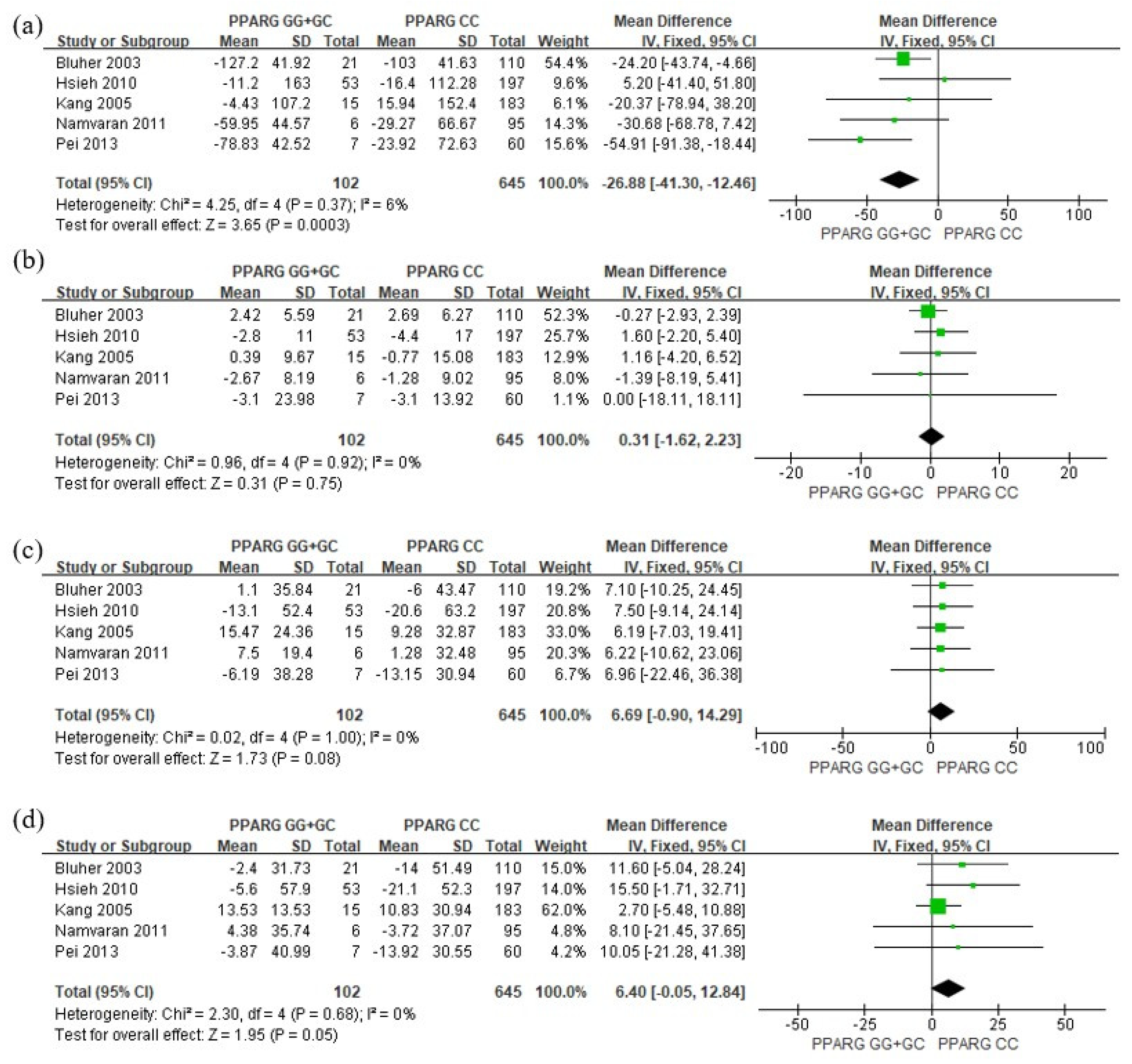
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Whiting, D.R.; Guariguata, L.; Weil, C.; Shaw, J. IDF Diabetes Atlas: Global estimates of the prevalence of diabetes for 2011 and 2030. Diabetes Res. Clin. Pract. 2011, 94, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Yki-Järvinen, H. Thiazolidinediones. New Engl. J. Med. 2004, 351, 1106–1118. [Google Scholar] [CrossRef] [PubMed]
- Grey, A. Skeletal consequences of thiazolidinedione therapy. Osteoporos. Int. 2008, 19, 129–137. [Google Scholar] [CrossRef]
- Hauner, H. The mode of action of thiazolidinediones. Diabetes Metab. Res. Rev. 2002, 18, S10–S15. [Google Scholar] [CrossRef] [PubMed]
- Fujita, T.; Sugiyama, Y.; Taketomi, S.; Sohda, T.; Kawamatsu, Y.; Iwatsuka, H.; Suzuoki, Z. Reduction of insulin resistance in obese and/or diabetic animals by 5-[4-(1-methylcyclohexylmethoxy) benzyl]-thiazolidine-2, 4-dione (ADD-3878, U-63,287, ciglitazone), a new antidiabetic agent. Diabetes 1983, 32, 804–810. [Google Scholar] [CrossRef]
- Fujiwara, T.; Yoshioka, S.; Yoshioka, T.; Ushiyama, I.; Horikoshi, H. Characterization of new oral antidiabetic agent CS-045: Studies in KK and ob/ob mice and Zucker fatty rats. Diabetes 1988, 37, 1549–1558. [Google Scholar] [CrossRef]
- Bays, H.; Mandarino, L.; DeFronzo, R.A. Role of the Adipocyte, Free Fatty Acids, and Ectopic Fat in Pathogenesis of Type 2 Diabetes Mellitus: Peroxisomal Proliferator-Activated Receptor Agonists Provide a Rational Therapeutic Approach. J. Clin. Endocrinol. Metab. 2004, 89, 463–478. [Google Scholar] [CrossRef] [Green Version]
- Jia, Z.; Sun, Y.; Yang, G.; Zhang, A.; Huang, S.; Heiney, K.M.; Zhang, Y. New insights into the PPARγ agonists for the treatment of diabetic nephropathy. PPAR Res. 2014, 2014, 818530. [Google Scholar] [CrossRef] [Green Version]
- Day, C. Thiazolidinediones: A new class of antidiabetic drugs. Diabet. Med. 1999, 16, 179–192. [Google Scholar] [CrossRef]
- Reginato, M.J.; Lazar, M.A. Mechanisms by which Thiazolidinediones Enhance Insulin Action. Trends Endocrinol. Metab. 1999, 10, 9–13. [Google Scholar] [CrossRef]
- Debril, M.-B.; Renaud, J.-P.; Fajas, L.; Auwerx, J. The pleiotropic functions of peroxisome proliferator-activated receptor γ. J. Mol. Med. 2001, 79, 30–47. [Google Scholar] [CrossRef]
- Mannino, G.C.; Andreozzi, F.; Sesti, G. Pharmacogenetics of type 2 diabetes mellitus, the route toward tailored medicine. Diabetes Metab. Res. Rev. 2019, 35, e3109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, C.X.; Ma, X.N.; Guan, C.H.; Li, Y.D.; Mauricio, D.; Fu, S.B. Cardiovascular disease in type 2 diabetes mellitus: Progress toward personalized management. Cardiovasc. Diabetol. 2022, 21, 74. [Google Scholar] [CrossRef]
- Aquilante, C.L. Pharmacogenetics of thiazolidinedione therapy. Pharmacogenomics 2007, 8, 917–931. [Google Scholar] [CrossRef] [PubMed]
- Kintscher, U.; Law, R.E. PPARγ-mediated insulin sensitization: The importance of fat versus muscle. Am. J. Physiol. Metab. 2005, 288, E287–E291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masugi, J.; Tamori, Y.; Mori, H.; Koike, T.; Kasuga, M. Inhibitory Effect of a Proline-to-Alanine Substitution at Codon 12 of Peroxisome Proliferator-Activated Receptor-γ 2 on Thiazolidinedione-Induced Adipogenesis. Biochem. Biophys. Res. Commun. 2000, 268, 178–182. [Google Scholar] [CrossRef]
- Gouda, H.N.; Sagoo, G.S.; Harding, A.H.; Yates, J.; Sandhu, M.S.; Higgins, J.P. The association between the peroxisome proliferator-activated receptor-γ2 (PPARG2) Pro12Ala gene variant and type 2 diabetes mellitus: A HuGE review and meta-analysis. Am. J. Epidemiol. 2010, 171, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. Int. J. Surg. 2021, 88, 105906. [Google Scholar] [CrossRef]
- Wells, G.A.; Shea, B.; O’Connell, D.; Peterson, J.; Welch, V.; Losos, M.; Tugwell, P. The Newcastle-Ottawa Scale (NOS) for Assessing the Quality of Nonrandomised Studies in Meta-Analyses 2002. Available online: http://www.ohri.ca/programs/clinical_epidemiology/oxford.asp (accessed on 12 June 2023).
- Higgins, J.P.; Thomas, J.; Chandler, J.; Cumpston, M.; Li, T.; Page, M.J.; Welch, V.A. Cochrane Handbook for Systematic Reviews of Interventions; John Wiley & Sons: Hoboken, NJ, USA, 2019. [Google Scholar]
- Higgins, J.P.T.; White, I.R.; Anzures-Cabrera, J. Meta-analysis of skewed data: Combining results reported on log-transformed or raw scales. Stat. Med. 2008, 27, 6072–6092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higgins, J.P.T.; Thompson, S.G. Quantifying heterogeneity in a meta-analysis. Stat. Med. 2002, 21, 1539–1558. [Google Scholar] [CrossRef]
- Begg, C.B.; Mazumdar, M. Operating Characteristics of a Rank Correlation Test for Publication Bias. Biometrics 1994, 50, 1088–1101. [Google Scholar] [CrossRef] [PubMed]
- Egger, M.; Smith, G.D.; Schneider, M.; Minder, C. Bias in meta-analysis detected by a simple, graphical test. BMJ 1997, 315, 629–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blüher, M.; Lübben, G.; Paschke, R. Analysis of the Relationship Between the Pro12Ala Variant in the PPAR-γ2 Gene and the Response Rate to Therapy with Pioglitazone in Patients with Type 2 Diabetes. Diabetes Care 2003, 26, 825–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, M.-C.; Lin, K.-D.; Tien, K.-J.; Tu, S.-T.; Hsiao, J.-Y.; Chang, S.-J.; Lin, S.-R.; Shing, S.-J.; Chen, H.-C. Common polymorphisms of the peroxisome proliferator-activated receptor–γ (Pro12Ala) and peroxisome proliferator-activated receptor–γ coactivator–1 (Gly482Ser) and the response to pioglitazone in Chinese patients with type 2 diabetes mellitus. Metabolism 2010, 59, 1139–1144. [Google Scholar] [CrossRef] [PubMed]
- Kang, E.S.; Park, S.Y.; Kim, H.J.; Kim, C.S.; Ahn, C.W.; Cha, B.S.; Lim, S.K.; Nam, C.M.; Lee, H.C. Effects of Pro12Ala polymorphism of peroxisome proliferator-activated receptor γ2 gene on rosiglitazone response in type 2 diabetes. Clin. Pharmacol. Ther. 2005, 78, 202–208. [Google Scholar] [CrossRef]
- Namvaran, F.; Azarpira, N.; Rahimi-Moghaddam, P.; Dabbaghmanesh, M.H. Polymorphism of peroxisome proliferator-activated receptor γ (PPARγ) Pro12Ala in the Iranian population: Relation with insulin resistance and response to treatment with pioglitazone in type 2 diabetes. Eur. J. Pharmacol. 2011, 671, 1–6. [Google Scholar] [CrossRef]
- Pei, Q.; Huang, Q.; Yang, G.P.; Zhao, J.C.; Jin, J.Y.; Song, M.; Zheng, Y.; Mo, Z.H.; Zhou, H.H.; Liu, Z.Q. PPAR-γ2 and PTPRD gene polymorphisms influence type 2 diabetes patients’ response to pioglitazone in China. Acta Pharmacol. Sin. 2013, 34, 255–261. [Google Scholar] [CrossRef] [Green Version]
- Priya, S.S.; Sankaran, R.; Ramalingam, S.; Sairam, T.; Somasundaram, L. Genotype phenotype correlation of genetic polymorphism of PPAR gamma gene and Therapeutic response to pioglitazone in type 2 diabetes mellitus-a pilot study. J. Clin. Diagn. Res. JCDR 2016, 10, FC11–FC14. [Google Scholar] [CrossRef]
- Berger, J.P.; Akiyama, T.E.; Meinke, P.T. PPARs: Therapeutic targets for metabolic disease. Trends Pharmacol. Sci. 2005, 26, 244–251. [Google Scholar] [CrossRef]
- Okuno, A.; Tamemoto, H.; Tobe, K.; Ueki, K.; Mori, Y.; Iwamoto, K.; Umesono, K.; Akanuma, Y.; Fujiwara, T.; Horikoshi, H.; et al. Troglitazone increases the number of small adipocytes without the change of white adipose tissue mass in obese Zucker rats. J. Clin. Investig. 1998, 101, 1354–1361. [Google Scholar] [CrossRef]
- Teboul, L.; Febbraio, M.; Gaillard, D.; Amri, E.-Z.; Silverstein, R.; Grimaldi, P.A. Structural and functional characterization of the mouse fatty acid translocase promoter: Activation during adipose differentiation. Biochem. J. 2001, 360, 305–312. [Google Scholar] [CrossRef]
- Filipova, E.; Uzunova, K.; Kalinov, K.; Vekov, T. Effects of pioglitazone therapy on blood parameters, weight and BMI: A meta-analysis. Diabetol. Metab. Syndr. 2017, 9, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebovitz, H.E.; Dole, J.F.; Patwardhan, R.; Rappaport, E.B.; Freed, M.I.; Rosiglitazone Clinical Trials Study Group. Rosiglitazone monotherapy is effective in patients with type 2 diabetes. J. Clin. Endocrinol. Metab. 2001, 86, 280–288. [Google Scholar] [CrossRef]
- Kadowaki, T.; Yamauchi, T.; Kubota, N.; Hara, K.; Ueki, K.; Tobe, K. Adiponectin and adiponectin receptors in insulin resistance, diabetes, and the metabolic syndrome. J. Clin. Investig. 2006, 116, 1784–1792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geloneze, B.; Pereira, J.A.; Pareja, J.C.; Lima, M.M.D.O.; Lazarin, M.A.C.T.; De Souza, I.C.P.; Tambascia, M.A.; Chaim, E.; Muscelli, E. Overcoming metabolic syndrome in severe obesity: Adiponectin as a marker of insulin sensitivity and HDL-cholesterol improvements after gastric bypass. Arq. Bras. Endocrinol. Metabol. 2009, 53, 293–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsubara, M.; Maruoka, S.; Katayose, S. Decreased plasma adiponectin concentrations in women with dyslipidemia. J. Clin. Endocrinol. Metab. 2002, 87, 2764–2769. [Google Scholar] [CrossRef]
- Chan, D.C.; Barrett, P.H.R.; Ooi, E.M.; Ji, J.; Chan, D.T.; Watts, G.F. Very low density lipoprotein metabolism and plasma adiponectin as predictors of high-density lipoprotein apolipoprotein AI kinetics in obese and nonobese men. J. Clin. Endocrinol. Metab. 2009, 94, 989–997. [Google Scholar] [CrossRef] [Green Version]
- Kimm, H.; Lee, S.W.; Lee, H.S.; Shim, K.W.; Cho, C.Y.; Yun, J.E.; Jee, S.H. Associations Between Lipid Measures and Metabolic Syndrome, Insulin Resistance and Adiponectin–Usefulness of Lipid Ratios in Korean Men and Women. Circ. J. 2010, 74, 931–937. [Google Scholar] [CrossRef] [Green Version]
- Christou, G.A.; Kiortsis, D.N. Adiponectin and lipoprotein metabolism. Obes. Rev. 2013, 14, 939–949. [Google Scholar] [CrossRef]
- Oz, O.; Tuncel, E.; Eryilmaz, S.; Fazlioglu, M.; Gul, C.B.; Ersoy, C.; Ocak, N.; Dirican, M.; Cangur, S.; Baran, I.; et al. Arterial elasticity and plasma levels of adiponectin and leptin in type 2 diabetic patients treated with thiazolidinediones. Endocrine 2008, 33, 101–105. [Google Scholar] [CrossRef]
- Tai, E.S.; Corella, D.; Deurenberg-Yap, M.; Adiconis, X.; Chew, S.K.; Tan, C.E.; Ordovas, J.M. Differential effects of the C1431T and Pro12Ala PPARγ gene variants on plasma lipids and diabetes risk in an Asian population. J. Lipid Res. 2004, 45, 674–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, L.; Wang, L.; Yun, H.; Su, L.; Su, X. Association of the PPAR gamma 2 gene Pro 12 Ala variant with primary hypertension and metabolic lipid disorders in Han Chinese of Inner Mongolia. Genet. Mol. Res. 2010, 9, 1312–1320. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Chen, R.; Bie, L.; Zhao, D.; Huang, C.; Hong, J. Association of the variants in the PPARG gene and serum lipid levels: A meta-analysis of 74 studies. J. Cell. Mol. Med. 2015, 19, 198–209. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Kamon, J.; Waki, H.; Murakami, K.; Motojima, K.; Komeda, K.; Ide, T.; Kubota, N.; Terauchi, Y.; Tobe, K.; et al. The mechanisms by which both heterozygous peroxisome prolifera-tor-activated receptor γ (PPARγ) deficiency and PPARγ agonist improve insulin resistance. J. Biol. Chem. 2001, 276, 41245–41254. [Google Scholar] [CrossRef] [Green Version]
- Vallo, J.; Arbas, R.; Basilio, J.E.; Cayabyab, I.; Miranda, C.N.; Santos, M.P.I.; Legaspi, L.F. Association of the Pro12Ala gene polymorphism with treatment response to thia-zolidinediones in patients with type 2 diabetes: A meta-analysis. Int. J. Diabetes Dev. Ctries. 2022, 42, 412–419. [Google Scholar] [CrossRef]
- Criqui, M.H.; Golomb, B.A. Epidemiologic aspects of lipid abnormalities. Am. J. Med. 1998, 105, 48S–57S. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| First Author, Year | Ethnicity | N (Male %) | Age, Year (SD) | Baseline BMI (SD) | TZD Treatment, mg/Day | Time to Outcome Extraction (Weeks) | Genotyping Methods | Outcomes Measured | Total NOS |
|---|---|---|---|---|---|---|---|---|---|
| Bluher et al., 2003 [25] | European | 131 (54.2) | 60.7 (9.3) | 31.0 (3.3) | Pioglitazone, 45 | 26 | PCR | HbA1C, FPG, TG, HDL, LDL, TC | 9 |
| Hsieh et al., 2010 [26] | Asian | 250 (47.6) | 57.86 (11.5) | 26.52 (4.4) | Pioglitazone, 30 | 24 | PCR-RFLP | HbA1C, FPG, TG, HDL, LDL, TC | 8 |
| Kang et al., 2005 [27] | Asian | 198 (52.5) | 56.5 (9.3) | 26.1 (2.8) | Rosiglitazone, 4 | 12 | PCR | HbA1C, FPG, TG, HDL, LDL, TC | 9 |
| Namvaran et al., 2011 [28] | Asian | 101 (20.8) | 51.44 (7.7) | 27.33 (4.2) | Pioglitazone, 15 | 12 | RT-PCR, TaqMan assay | HbA1C, FPG, TG, HDL, LDL, TC | 8 |
| Pei et al., 2013 [29] | Asian | 67 (58.2) | 56.63 (8.6) | 25.15 (2.8) | Pioglitazone, 30 | 12 | MALDI-TOF | HbA1C, FPG, TG, HDL, LDL, TC | 8 |
| Priya et al., 2016 [30] | Asian | 30 (46.0) | 53.2 (11.0) | 26.25 (3.7) | Pioglitazone, 30 | 12 | PCR | HbA1C, FPG | 8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jang, E.J.; Lee, D.H.; Im, S.-S.; Yee, J.; Gwak, H.S. Correlation between PPARG Pro12Ala Polymorphism and Therapeutic Responses to Thiazolidinediones in Patients with Type 2 Diabetes: A Meta-Analysis. Pharmaceutics 2023, 15, 1778. https://doi.org/10.3390/pharmaceutics15061778
Jang EJ, Lee DH, Im S-S, Yee J, Gwak HS. Correlation between PPARG Pro12Ala Polymorphism and Therapeutic Responses to Thiazolidinediones in Patients with Type 2 Diabetes: A Meta-Analysis. Pharmaceutics. 2023; 15(6):1778. https://doi.org/10.3390/pharmaceutics15061778
Chicago/Turabian StyleJang, Eun Jeong, Da Hoon Lee, Sae-Seul Im, Jeong Yee, and Hye Sun Gwak. 2023. "Correlation between PPARG Pro12Ala Polymorphism and Therapeutic Responses to Thiazolidinediones in Patients with Type 2 Diabetes: A Meta-Analysis" Pharmaceutics 15, no. 6: 1778. https://doi.org/10.3390/pharmaceutics15061778