
Arginine-Rich Cell-Penetrating Peptide-Mediated Transduction of Mouse Nasal Cells with FOXP3 Protein Alleviates Allergic Rhinitis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
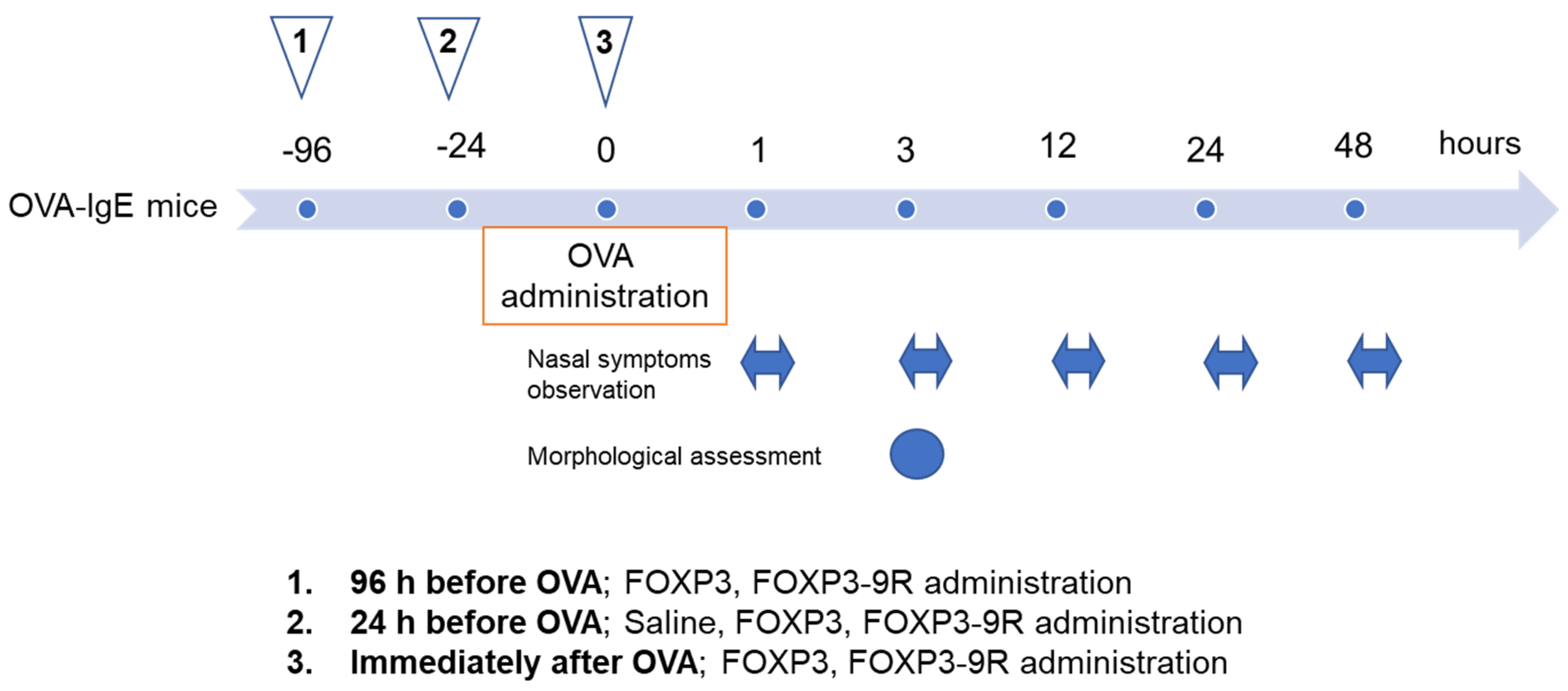
2. Materials and Methods
2.1. Recombinant FOXP3 and 9R Peptide-Conjugated FOXP3
2.2. Immunoglobulin E (IgE) Transgenic Mice
2.3. Protein Administration into the Mouse Nasal Cavity
2.4. Assessment of Nasal Symptoms
2.5. Immunohistochemistry
2.6. Hematoxylin and Eosin (H&E) Staining
2.7. Antibody Staining and Flow Cytometry
2.8. Dot Blotting
2.9. Statistical Analysis
3. Results
3.1. Nasal Symptoms after Administration of OVA in OVA-IgE Mice
3.2. Effect of FOXP3-9R on Nasal Symptoms
3.3. Histopathological Changes
3.4. Flow Cytometric Analysis and Immunohistochemistry
3.5. Cytokine Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Okubo, K.; Kurono, Y.; Ichimura, K.; Enomoto, T.; Okamoto, Y.; Kawauchi, H.; Suzaki, H.; Fujieda, S.; Masuyama, K. Japanese Guidelines for Allergic Rhinitis 2017. Allergol. Int. 2017, 66, 205–219. [Google Scholar] [CrossRef] [PubMed]
- Bousquet, J.; Hellings, P.W.; Agache, I.; Amat, F.; Annesi-Maesano, I.; Ansotegui, I.J.; Anto, J.M.; Bachert, C.; Bateman, E.D.; Bedbrook, A.; et al. Allergic Rhinitis and Its Impact on Asthma (ARIA) Phase 4 (2018): Change Management in Allergic Rhinitis and Asthma Multimorbidity Using Mobile Technology. J. Allergy Clin. Immunol. 2019, 143, 864–879. [Google Scholar] [CrossRef] [Green Version]
- Sousa, J.; Alves, G.; Fortuna, A.; Falcão, A. Intranasal Delivery of Topically-Acting Levofloxacin to Rats: A Proof-of-Concept Pharmacokinetic Study. Pharm. Res. 2017, 34, 2260–2269. [Google Scholar] [CrossRef] [PubMed]
- Pardeshi, C.V.; Rajput, P.V.; Belgamwar, V.S.; Tekade, A.R.; Surana, S.J. Novel Surface Modified Solid Lipid Nanoparticles as Intranasal Carriers for Ropinirole Hydrochloride: Application of Factorial Design Approach. Drug Deliv. 2013, 20, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Warnken, Z.N.; Smyth, H.D.C.; Watts, A.B.; Weitman, S.; Kuhn, J.G.; Williams, R.O. Formulation and Device Design to Increase Nose to Brain Drug Delivery. J. Drug Deliv. Sci. Technol. 2016, 35, 213–222. [Google Scholar] [CrossRef]
- Pardeshi, C.V.; Belgamwar, V.S. Direct Nose to Brain Drug Delivery via Integrated Nerve Pathways Bypassing the Blood–Brain Barrier: An Excellent Platform for Brain Targeting. Expert Opin. Drug Deliv. 2013, 10, 957–972. [Google Scholar] [CrossRef] [PubMed]
- Altuntaş, E.; Yener, G.; Doğan, R.; Aksoy, F.; Şerif Aydın, M.; Karataş, E. Effects of a Thermosensitive in Situ Gel Containing Mometasone Furoate on a Rat Allergic Rhinitis Model. Am. J. Rhinol. Allergy 2018, 32, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Awad, R.; Avital, A.; Sosnik, A. Polymeric Nanocarriers for Nose-to-Brain Drug Delivery in Neurodegenerative Diseases and Neurodevelopmental Disorders. Acta Pharm. Sin. B 2023, 13, 1866–1886. [Google Scholar] [CrossRef]
- Guidotti, G.; Brambilla, L.; Rossi, D. Cell-Penetrating Peptides: From Basic Research to Clinics. Trends Pharmacol. Sci. 2017, 38, 406–424. [Google Scholar] [CrossRef]
- Yousef, M.; Szabó, I.; Murányi, J.; Illien, F.; Soltész, D.; Bató, C.; Tóth, G.; Batta, G.; Nagy, P.; Sagan, S.; et al. Cell-Penetrating Dabcyl-Containing Tetraarginines with Backbone Aromatics as Uptake Enhancers. Pharmaceutics 2022, 15, 141. [Google Scholar] [CrossRef]
- Zorko, M.; Langel, Ü. Cell-Penetrating Peptides. Methods Mol. Biol. 2022, 2383, 3–32. [Google Scholar] [CrossRef]
- Miwa, T.; Tachii, K.; Wei, F.Y.; Kaitsuka, T.; Tomizawa, K. Intranasal Drug Delivery into Mouse Nasal Mucosa and Brain Utilizing Arginine-Rich Cell-Penetrating Peptide-Mediated Protein Transduction. Int. J. Pept. Res. Ther. 2020, 26, 1643–1650. [Google Scholar] [CrossRef]
- Miwa, T.; Minoda, R.; Kaitsuka, T.; Ise, M.; Tomizawa, K.; Yumoto, E. Protein Transduction into the Mouse Otocyst Using Arginine-Rich Cell-Penetrating Peptides. Neuroreport 2011, 22, 994–999. [Google Scholar] [CrossRef]
- Takeda, H.; Kurioka, T.; Kaitsuka, T.; Tomizawa, K.; Matsunobu, T.; Hakim, F.; Mizutari, K.; Miwa, T.; Yamada, T.; Ise, M.; et al. Protein Transduction Therapy into Cochleae via the Round Window Niche in Guinea Pigs. Mol. Ther.-Methods Clin. Dev. 2016, 3, 16055. [Google Scholar] [CrossRef]
- Matsuoka, T.; Shamji, M.H.; Durham, S.R. Allergen Immunotherapy and Tolerance. Allergol. Int. 2013, 62, 403–413. [Google Scholar] [CrossRef] [Green Version]
- Terada, T.; Matsuda, M.; Inaba, M.; Hamaguchi, J.; Takemoto, N.; Kikuoka, Y.; Inaka, Y.; Sakae, H.; Hashimoto, K.; Shimora, H.; et al. Sublingual Immunotherapy for 4 Years Increased the Number of Foxp3+ Treg Cells, Which Correlated with Clinical Effects. Inflamm. Res. 2021, 70, 581–589. [Google Scholar] [CrossRef]
- Hori, S.; Nomura, T.; Sakaguchi, S. Control of Regulatory T Cell Development by the Transcription Factor Foxp3. Science 2003, 299, 1057–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, L.; Barbi, J.; Pan, F. The Regulation of Immune Tolerance by FOXP3. Nat. Rev. Immunol. 2017, 17, 703–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allan, S.E.; Alstad, A.N.; Merindol, N.; Crellin, N.K.; Amendola, M.; Bacchetta, R.; Naldini, L.; Roncarolo, M.G.; Soudeyns, H.; Levings, M.K. Generation of Potent and Stable Human CD4+ T Regulatory Cells by Activation-Independent Expression of FOXP3. Mol. Ther. 2008, 16, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, M.; Tomizawa, K.; Moriwaki, A.; Li, S.T.; Terada, H.; Matsui, H. A High-Efficiency Protein Transduction System Demonstrating the Role of PKA in Long-Lasting Long-Term Potentiation. J. Neurosci. 2001, 21, 6000–6007. [Google Scholar] [CrossRef] [Green Version]
- Sato, E.; Hirahara, K.; Wada, Y.; Yoshitomi, T.; Azuma, T.; Matsuoka, K.; Kubo, S.; Taya, C.; Yonekawa, H.; Karasuyama, H.; et al. Chronic Inflammation of the Skin Can Be Induced in IgE Transgenic Mice by Means of a Single Challenge of Multivalent Antigen. J. Allergy Clin. Immunol. 2003, 111, 143–148. [Google Scholar] [CrossRef]
- Omata, N.; Ohshima, Y.; Yasutomi, M.; Yamada, A.; Karasuyama, H.; Mayumi, M. Ovalbumin-Specific IgE Modulates Ovalbumin-Specific T-Cell Response after Repetitive Oral Antigen Administration. J. Allergy Clin. Immunol. 2005, 115, 822–827. [Google Scholar] [CrossRef]
- Wen, W.-D.; Yuan, F.; Wang, J.-L.; Hou, Y.-P. Botulinum Toxin Therapy in the Ovalbumin-Sensitized Rat. Neuroimmunomodulation 2007, 14, 78–83. [Google Scholar] [CrossRef]
- Fontenot, J.D.; Rasmussen, J.P.; Williams, L.M.; Dooley, J.L.; Farr, A.G.; Rudensky, A.Y. Regulatory T Cell Lineage Specification by the Forkhead Transcription Factor Foxp3. Immunity 2005, 22, 329–341. [Google Scholar] [CrossRef]
- Takahashi, R.; Nishimoto, S.; Muto, G.; Sekiya, T.; Tamiya, T.; Kimura, A.; Morita, R.; Asakawa, M.; Chinen, T.; Yoshimura, A. SOCS1 Is Essential for Regulatory T Cell Functions by Preventing Loss of Foxp3 Expression as Well as IFN-γ and IL-17A Production. J. Exp. Med. 2011, 208, 2055. [Google Scholar] [CrossRef]
- Miwa, T.; Minoda, R.; Ise, M.; Yamada, T.; Yumoto, E. Mouse Otocyst Transuterine Gene Transfer Restores Hearing in Mice with Connexin 30 Deletion-Associated Hearing Loss. Mol. Ther. 2013, 21, 1142–1150. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.Y. Sublingual Immunotherapy: Current Concepts for the U.S. Practitioner. Int. Forum Allergy Rhinol. 2014, 4, e7694. [Google Scholar] [CrossRef] [PubMed]
- Abràmoff, M.D.; Magalhães, P.J.; Ram, S.J. Image Processing with ImageJ. Biophotonics Int. 2004, 11, 36–41. [Google Scholar]
- Eguchi, A.; Meade, B.R.; Chang, Y.-C.; Fredrickson, C.T.; Willert, K.; Puri, N.; Dowdy, S.F. Efficient SiRNA Delivery into Primary Cells by a Peptide Transduction Domain–DsRNA Binding Domain Fusion Protein. Nat. Biotechnol. 2009, 27, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.Y.; Ni, X.C.; Yi, Y.; He, H.W.; Wang, J.X.; Fu, Y.P.; Sun, J.; Zhou, J.; Cheng, Y.F.; Jin, J.J.; et al. Overexpression of CD39 in Hepatocellular Carcinoma Is an Independent Indicator of Poor Outcome after Radical Resection. Medicine 2016, 95, e4989. [Google Scholar] [CrossRef]
- Fujita, H.; Soyka, M.B.; Akdis, M.; Akdis, C.A. Mechanisms of Allergen-Specific Immunotherapy. Clin. Transl. Allergy 2012, 2, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feifei, X.; Shaoqing, Y.; Chuanliang, Z.; Na, C.; Ling, J.; Qiuli, W.; Rongming, G. Effect of Hydrogen-Rich Saline on the CD4 + CD25 + Foxp3 + Treg Cells of Allergic Rhinitis Guinea Pigs Model. Chin. J. Otorhinolaryngol. Head Neck Surg. 2017, 52, 506–511. [Google Scholar] [CrossRef]
- Bellinghausen, I.; Khatri, R.; Saloga, J. Current Strategies to Modulate Regulatory T Cell Activity in Allergic Inflammation. Front. Immunol. 2022, 13, 912529. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Chong, M.M.W.; Littman, D.R. Plasticity of CD4+ T Cell Lineage Differentiation. Immunity 2009, 30, 646–655. [Google Scholar] [CrossRef] [Green Version]
- Ezzeddini, R.; Somi, M.H.; Taghikhani, M.; Moaddab, S.Y.; Masnadi Shirazi, K.; Shirmohammadi, M.; Eftekharsadat, A.T.; Sadighi Moghaddam, B.; Salek Farrokhi, A. Association of Foxp3 Rs3761548 Polymorphism with Cytokines Concentration in Gastric Adenocarcinoma Patients. Cytokine 2021, 138, 155351. [Google Scholar] [CrossRef]
- Takenaka, M.; Seki, N.; Toh, U.; Hattori, S.; Kawahara, A.; Yamaguchi, T.; Koura, K.; Takahashi, R.; Otsuka, H.; Takahashi, H.; et al. FOXP3 Expression in Tumor Cells and Tumor-Infiltrating Lymphocytes Is Associated with Breast Cancer Prognosis. Mol. Clin. Oncol. 2013, 1, 625–632. [Google Scholar] [CrossRef] [Green Version]
- Raffin, C.; Vo, L.T.; Bluestone, J.A. Treg Cell-Based Therapies: Challenges and Perspectives. Nat. Rev. Immunol. 2020, 20, 158. [Google Scholar] [CrossRef]
- Brimnes, J.; Kildsgaard, J.; Jacobi, H.; Lund, K. Sublingual Immunotherapy Reduces Allergic Symptoms in a Mouse Model of Rhinitis. Clin. Exp. Allergy 2007, 37, 488–497. [Google Scholar] [CrossRef]
- Durham, S.R.; Yang, W.H.; Pedersen, M.R.; Johansen, N.; Rak, S. Sublingual Immunotherapy with Once-Daily Grass Allergen Tablets: A Randomized Controlled Trial in Seasonal Allergic Rhinoconjunctivitis. J. Allergy Clin. Immunol. 2006, 117, 802–809. [Google Scholar] [CrossRef]
- Palomares, O.; Yaman, G.; Azkur, A.K.; Akkoc, T.; Akdis, M.; Akdis, C.A. Role of Treg in Immune Regulation of Allergic Diseases. Eur. J. Immunol. 2010, 40, 1232–1240. [Google Scholar] [CrossRef]
- Ghyselinck, N.B.; Duester, G. Retinoic Acid Signaling Pathways. Development 2019, 146, dev167502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goswami, T.K.; Singh, M.; Dhawan, M.; Mitra, S.; Bin Emran, T.; Rabaan, A.A.; Al Mutair, A.; Al Alawi, Z.; Alhumaid, S.; Dhama, K. Regulatory T Cells (Tregs) and Their Therapeutic Potential against Autoimmune Disorders–Advances and Challenges. Hum. Vaccines Immunother. 2022, 18, 2035117. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhao, X.; Wan, Y.Y. Intricacies of TGF-β Signaling in Treg and Th17 Cell Biology. Cell. Mol. Immunol. 2023, 1–21. [Google Scholar] [CrossRef]
- Liu, W.; Fan, M.; Lu, W.; Zhu, W.; Meng, L.; Lu, S. Emerging Roles of T Helper Cells in Non-Infectious Neuroinflammation: Savior or Sinner. Front. Immunol. 2022, 13, 3208. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miwa, T.; Takemiya, Y.; Amesara, K.; Kawai, H.; Teranishi, Y. Arginine-Rich Cell-Penetrating Peptide-Mediated Transduction of Mouse Nasal Cells with FOXP3 Protein Alleviates Allergic Rhinitis. Pharmaceutics 2023, 15, 1770. https://doi.org/10.3390/pharmaceutics15061770
Miwa T, Takemiya Y, Amesara K, Kawai H, Teranishi Y. Arginine-Rich Cell-Penetrating Peptide-Mediated Transduction of Mouse Nasal Cells with FOXP3 Protein Alleviates Allergic Rhinitis. Pharmaceutics. 2023; 15(6):1770. https://doi.org/10.3390/pharmaceutics15061770
Chicago/Turabian StyleMiwa, Toru, Yumi Takemiya, Kazuki Amesara, Hiroko Kawai, and Yuichi Teranishi. 2023. "Arginine-Rich Cell-Penetrating Peptide-Mediated Transduction of Mouse Nasal Cells with FOXP3 Protein Alleviates Allergic Rhinitis" Pharmaceutics 15, no. 6: 1770. https://doi.org/10.3390/pharmaceutics15061770