
Identification and Characterisation of a Myxoma Virus Detected in the Italian Hare (Lepus corsicanus)

 , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
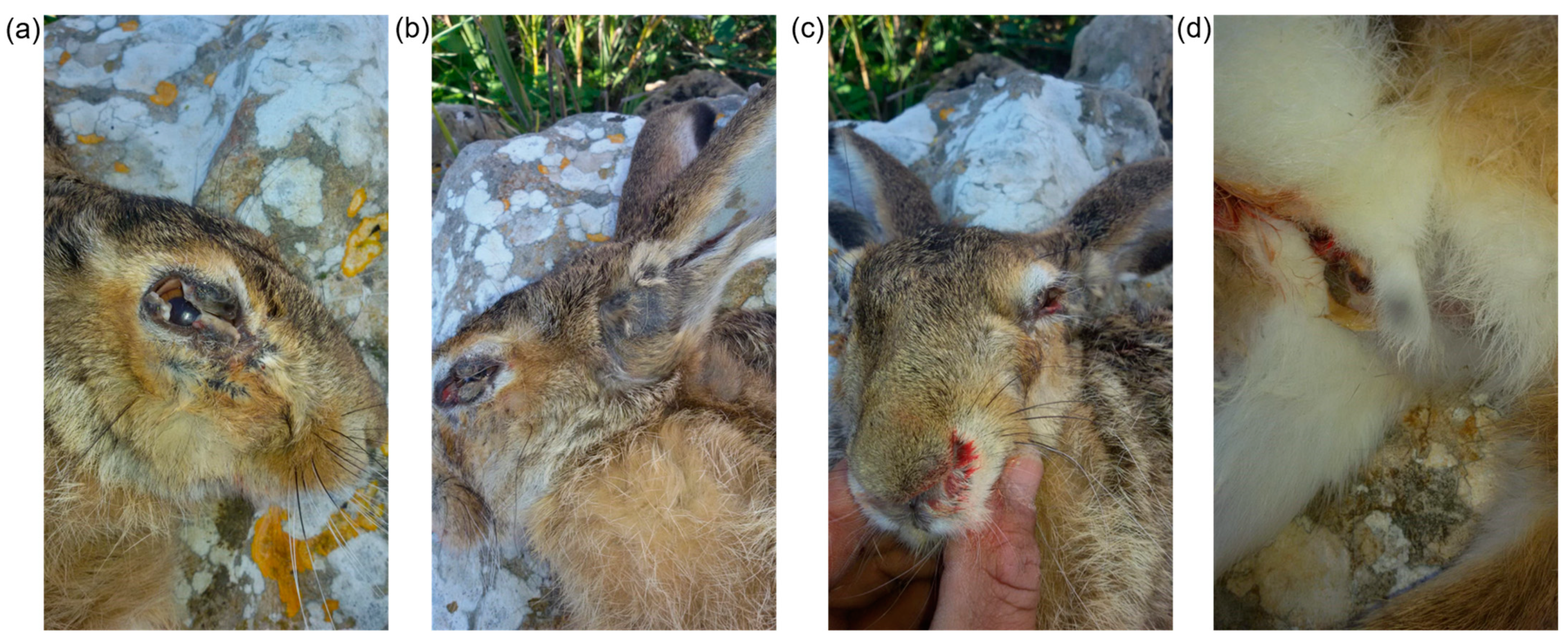
2.1. Sampling and Pathology
2.2. Viral Isolation
2.3. Electron Microscopy
2.4. DNA Extraction and PCR Amplification
2.5. Whole Genome Sequencing
3. Results
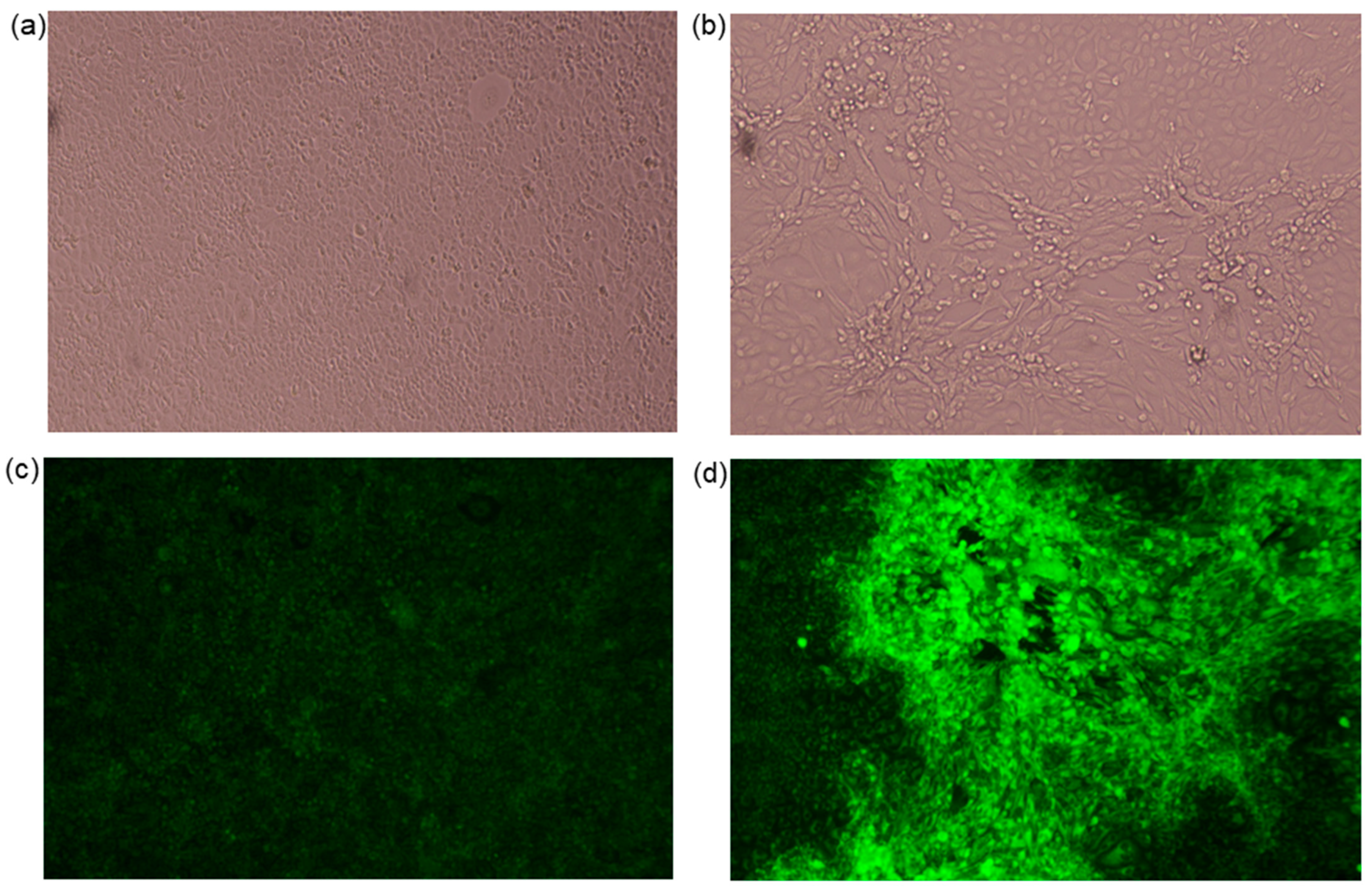
3.1. Viral Isolation
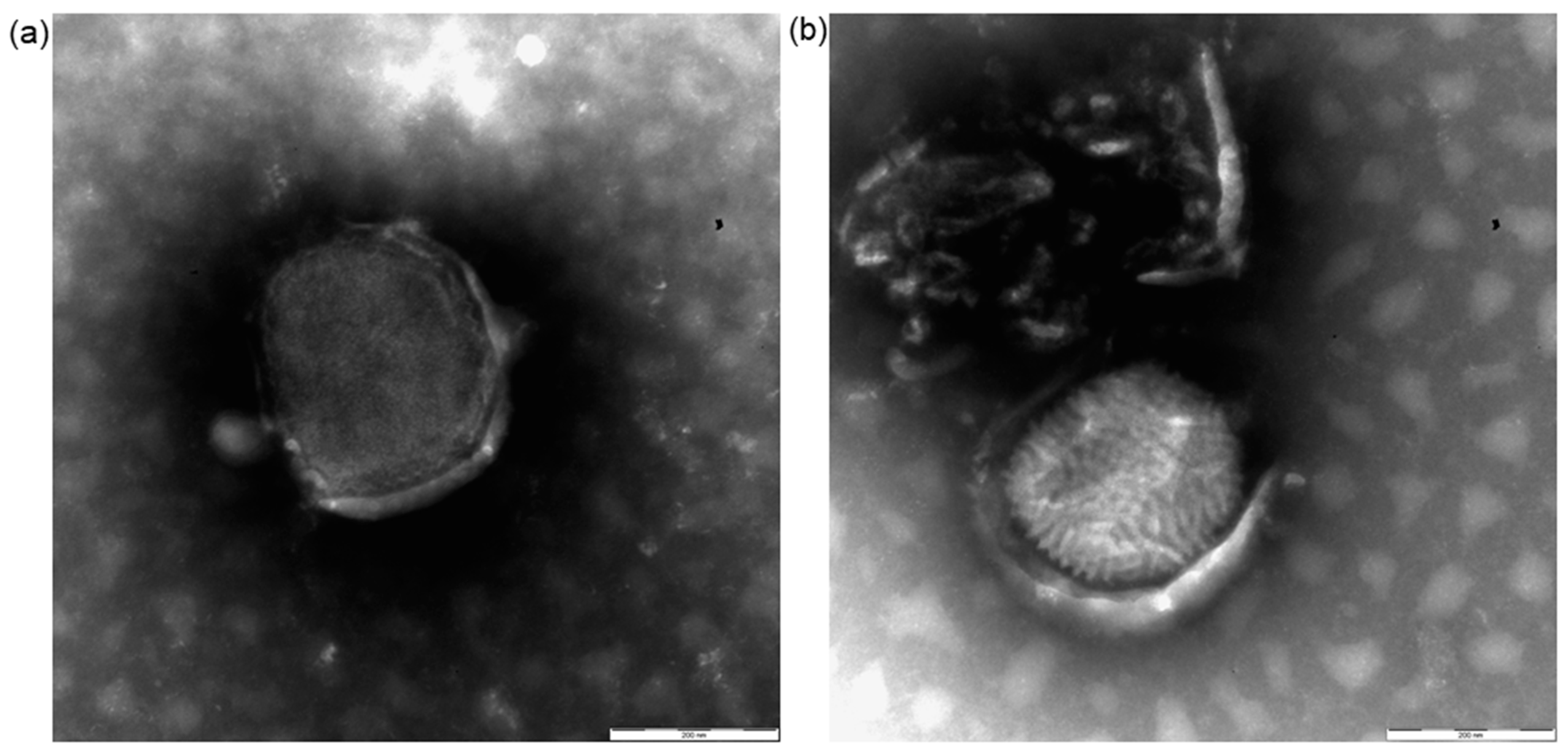
3.2. Electron Microscopy
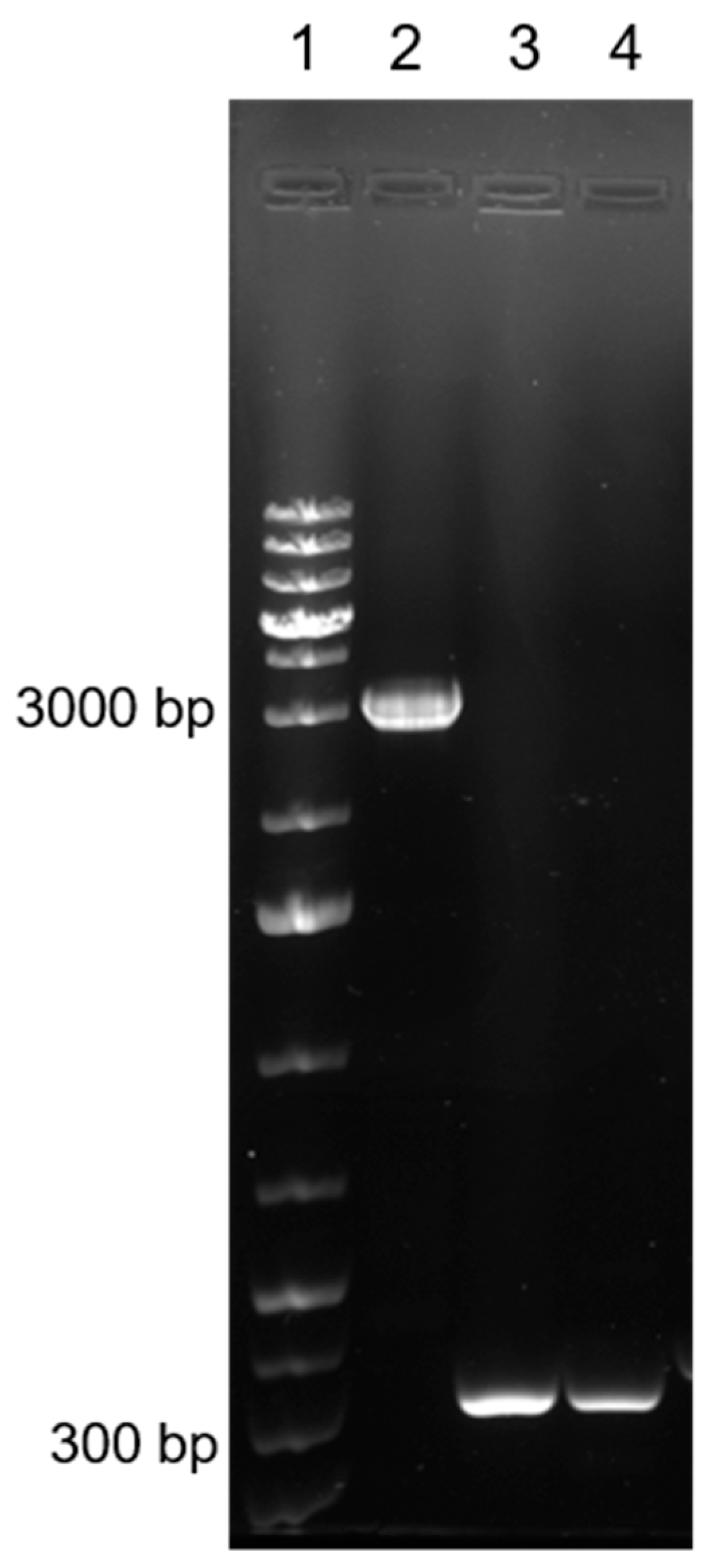
3.3. Molecular Characterisation
3.4. Whole Genome Sequencing
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fenner, F. Biological control, as exemplified by smallpox eradication and myxomatosis. Proc. R. Soc. 1983, 218, 259–285. [Google Scholar] [CrossRef]
- Cameron, C.; Hota-Mitchell, S.; Chen, L.; Barrett, J.; Cao, J.X.; Macaulay, C.; Willer, D.; Evans, D.; McFadden, G. The complete DNA sequence of myxoma virus. Virology 1999, 264, 298–318. [Google Scholar] [CrossRef]
- Duteyrat, J.L.; Gelfi, J.; Bertagnoli, S. Ultrastructural study of myxoma virus morphogenesis. Arch. Virol. 2006, 151, 2161–2180. [Google Scholar] [CrossRef]
- Farsang, A.; Makranszki, L.; Dobos-Kovacs, M.; Virag, G.; Fabian, K.; Barna, T.; Kuclsar, G.; Kucsera, L.; Vetesi, F. Occurrence of atypical myxomatosis in central Europe: Clinical and virological examinations. Acta Vet. Hung. 2003, 51, 493–501. [Google Scholar] [CrossRef]
- Fenner, F. Myxomatosis. Br. Med. Bull. 1959, 15, 240–245. [Google Scholar] [CrossRef]
- Kerr, P.; McFadden, G. Immune responses to myxoma virus. Viral Immunol. 2002, 15, 229–246. [Google Scholar] [CrossRef]
- Spiesschaert, B.; McFadden, G.; Hermans, K.; Nauwynck, H.; Van de Walle, G.R. The current status and future directions of myxoma virus, a master in immune evasion. Vet. Res. 2011, 42, 76. [Google Scholar] [CrossRef]
- Kerr, P.J.; Cattadori, I.M.; Rogers, M.B.; Fitch, A.; Geber, A.; Liu, J.; Sim, D.G.; Boag, B.; Eden, J.S.; Ghedin, E.; et al. Genomic and phenotypic characterization of myxoma virus from Great Britain reveals multiple evolutionary pathways distinct from those in Australia. PLoS Pathog. 2017, 13, e1006252. [Google Scholar] [CrossRef] [PubMed]
- Kerr, P.J.; Liu, J.; Cattadori, I.; Ghedin, E.; Read, A.F.; Holmes, E.C. Myxoma virus and the Leporipoxviruses: An evolutionary paradigm. Viruses 2015, 7, 1020–1061. [Google Scholar] [CrossRef]
- Barlow, A.; Lawrence, K.; Everest, D.; Dastjerdi, A.; Finnegan, C.; Steinbach, F. Confirmation of myxomatosis in a European brown hare in Great Britain. Vet. Rec. 2014, 175, 75–76. [Google Scholar] [CrossRef]
- García-Bocanegra, I.; Camacho-Sillero, L.; Risalde, M.A.; Dalton, K.P.; Caballero-Gómez, J.; Agüero, M.; Zorrilla, I.; Gómez-Guillamón, F. First outbreak of myxomatosis in Iberian hares (Lepus granatensis). Transbound. Emerg. Dis. 2019, 66, 2204–2208. [Google Scholar] [CrossRef]
- Águeda-Pinto, A.; Lemos de Matos, A.; Abrantes, M.; Kraberger, S.; Risalde, M.A.; Gortázar, C.; McFadden, G.; Varsani, A.; Esteves, P.J. Genetic Characterization of a Recombinant Myxoma Virus in the Iberian Hare (Lepus granatensis). Viruses 2019, 11, 530. [Google Scholar] [CrossRef]
- Abade Dos Santos, F.A.; Carvalho, C.L.; Monteiro, M.; Carvalho, P.; Mendonça, P.; Peleteiro, M.D.C.; Duarte, M.D. Recombinant myxoma virus infection associated with high mortality in rabbit farming (Oryctolagus cuniculus). Transbound. Emerg. Dis. 2021, 68, 2616–2621. [Google Scholar] [CrossRef]
- Lumeij, J.T.; de Koning, J.; Bosma, R.B.; van der Sluis, J.J.; Schellekens, J.F. Treponemal infections in hares in The Netherlands. J. Clin. Microbiol. 1994, 32, 543–546. [Google Scholar] [CrossRef]
- Cavadini, P.; Trogu, T.; Velarde, R.; Lavazza, A.; Capucci, L. Recombination between non-structural and structural genes as a mechanism of selection in lagoviruses: The evolutionary dead-end of an RHDV2 isolated from European hare. Virus Res. 2024, 339, 199257. [Google Scholar] [CrossRef] [PubMed]
- Hisgen, L.; Abel, L.; Hallmaier-Wacker, L.K.; Lueert, S.; Siebert, U.; Faehndrich, M.; Strauss, E.; Voigt, U.; Nováková, M.; Šmajs, D.; et al. High syphilis seropositivity in European brown hares (Lepus europaeus), Lower Saxony, Germany. Transbound. Emerg. Dis. 2020, 67, 2240–2244. [Google Scholar] [CrossRef]
- Cavadini, P.; Botti, G.; Barbieri, I.; Lavazza, A.; Capucci, L. Molecular characterization of SG33 and Borghi vaccines used against myxomatosis. Vaccine 2010, 28, 5414–5420. [Google Scholar] [CrossRef] [PubMed]
- World Organization for Animal Health, WOAH. Chapter 3.7.1. Myxomatosis. B. Diagnostic Techniques—1.6 Molecular Methods—detection of nucleic acid. In Terrestrial Manual; WOAH: Paris, France, 2021. [Google Scholar]
- Galaxy Community. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2022 update. Nucleic Acids Res. 2022, 50, W345–W351. [Google Scholar] [CrossRef]
- Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 7 March 2024).
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Chevreux, B.; Pfisterer, T.; Drescher, B.; Driesel, A.J.; Müller, W.E.; Wetter, T.; Suhai, S. Using the miraEST assembler for reliable and automated mRNA transcript assembly and SNP detection in sequenced ESTs. Genome Res. 2004, 14, 1147–1159. [Google Scholar] [CrossRef]
- Zerbino, D.R.; Birney, E. Velvet: Algorithms for de novo short read assembly using de Bruijn graphs. Genome Res. 2008, 18, 821–829. [Google Scholar] [CrossRef]
- World Organization for Animal Health, WOAH. Chapter 3.7.1. Myxomatosis. B. Diagnostic Techniques—1.3. In-vitro cell culture. In Terrestrial Manual; WOAH: Paris, France, 2021. [Google Scholar]
- Wilson, T.M.; Sweeney, P.R. Morphological studies of seal poxvirus. J. Wildl. Dis. 1970, 6, 94–97. [Google Scholar] [CrossRef] [PubMed]
- Dalton, K.P.; Nicieza, I.; Baragaño, A.; Alonso, J.M.; Parra, F. Molecular characterisation of virulence graded field isolates of myxoma virus. Virol. J. 2010, 7, 49. [Google Scholar] [CrossRef] [PubMed]
- Kerr, P.J.; Ghedin, E.; DePasse, J.V.; Fitch, A.; Cattadori, I.M.; Hudson, P.J.; Tscharke, D.C.; Read, A.F.; Holmes, E.C. Evolutionary history and attenuation of myxoma virus on two continents. PLoS Pathog. 2012, 8, e1002950. [Google Scholar] [CrossRef] [PubMed]
- Kerr, P.J.; Rogers, M.B.; Fitch, A.; Depasse, J.V.; Cattadori, I.M.; Hudson, P.J.; Tscharke, D.C.; Holmes, E.C.; Ghedin, E. Comparative analysis of the complete genome sequence of the California MSW strain of myxoma virus reveals potential host adaptations. J. Virol. 2013, 87, 12080–12089. [Google Scholar] [CrossRef]
- Haller, S.L.; Peng, C.; McFadden, G.; Rothenburg, S. Poxviruses and the evolution of host range and virulence. Infect. Genet. Evol. 2014, 21, 15–40. [Google Scholar] [CrossRef]
- Peng, C.; Haller, S.L.; Rahman, M.M.; McFadden, G.; Rothenburg, S. Myxoma virus M156 is a specific inhibitor of rabbit PKR but contains a loss-of-function mutation in Australian virus isolates. Proc. Natl. Acad. Sci. USA 2016, 113, 3855–3860. [Google Scholar] [CrossRef]
- McNab, F.; Mayer-Barber, K.; Sher, A.; Wack, A.; O’Garra, A. Type I interferons in infectious disease. Nat. Rev. Immunol. 2015, 15, 87–103. [Google Scholar] [CrossRef]
- Gasteiger, G.; D’Osualdo, A.; Schubert, D.A.; Weber, A.; Bruscia, E.M.; Hartl, D. Cellular Innate Immunity: An Old Game with New Players. J. Innate Immun. 2017, 9, 111–125. [Google Scholar] [CrossRef]
- Yu, H.; Bruneau, R.C.; Brennan, G.; Rothenburg, S. Battle Royale: Innate Recognition of Poxviruses and Viral Immune Evasion. Biomedicines 2021, 9, 765. [Google Scholar] [CrossRef]
- Yu, H.; Peng, C.; Zhang, C.; Stoian, A.M.M.; Tazi, L.; Brennan, G.; Rothenburg, S. Maladaptation after a virus host switch leads to increased activation of the pro-inflammatory NF-κB pathway. Proc. Natl. Acad. Sci. USA 2022, 119, e2115354119. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequences of Primers for MYXV Infection Diagnosis | |||
|---|---|---|---|
| Primer ID | Sequence (5′-3′) | Position nt a | Expected Amplicon (bp) |
| M071-F | ACCCGCCAAGAACCACAGTAGT | 67,229–67,250 | 272 |
| M071-R | TAACGCGAGGAATATCCTGTACCA | 67,700–67,677 | |
| M009-F | ATACACGCCGACGCATTACG | 12,181–12,200 | 316 |
| M009-R | ACGAGAGATACGCTGAAGAAC | 12,496–12,476 | |
| Sequences of primers for MYXV TIR amplification | |||
| M0005-F | ACGCGGAAGTCTGCCTATTT | 230–249 | 12,266 |
| M009-R | ACGAGAGATACGCTGAAGAAC | 12,496–12,476 | |
| M153-F | CATTTATGGTATCCGCATTAAC | 148,944–148,965 | 12,600 |
| M0005-F | ACGCGGAAGTCTGCCTATTT | 161,544–161,525 | |
| Gene Name | Function of Gene Product | Start | Stop | +/− | Gene Size | Protein Size | MYXV-Lu Protein Size | % aa Similarity | aa Differences |
|---|---|---|---|---|---|---|---|---|---|
| M002L | Tumor necrosis factor receptor (TNF-R) homologue | 1664 | 2644 | − | 981 | 326 | 326 | 99.7 | A131V, E224K |
| M005L | E3 Ub ligase | 4864 | 6315 | − | 1452 | 483 | 483 | 99.8 | V320A |
| M012L | dUTP nucleotidylhydrolase | 14,070 | 14,516 | − | 447 | 148 | 148 | 99.3 | K47T |
| M031R | Virosome protein | 30,071 | 31,252 | + | 1182 | 393 | 393 | 99.7 | R75H |
| M032R | VACV E6R | 31,262 | 32,959 | + | 1698 | 565 | 565 | 99.8 | G190V |
| M034L | DNA Pol | 33,777 | 36,797 | − | 3021 | 1 006 | 1 006 | 100.0 | V620I |
| M036L | VACV O1L/Erk1/2 signaling? | 37,142 | 39,184 | − | 2043 | 680 | 680 | 99.7 | R95C |
| M044R | RNA helicase NPH-II | 44,090 | 46,126 | + | 2037 | 678 | 678 | 99.9 | P82S |
| M064R | Host range protein | 59,564 | 60,178 | + | 615 | 204 | 203 | 99.0 | ▼164E, D173G |
| M079R | Uracil-DNA glycosylase | 76,264 | 76,920 | + | 657 | 218 | 218 | 99.5 | A183T |
| M092L | Core protein | 89,899 | 91,860 | − | 1962 | 653 | 653 | 99.8 | S85L |
| M094R | RNA Pol subunit | 92,414 | 92,908 | + | 495 | 164 | 164 | 99.4 | D15G |
| M096L | Early transcription factor subunit | 94,054 | 96,189 | − | 2136 | 711 | 711 | 100.0 | N472S |
| M099L | Core protein precursor | 97,331 | 100,036 | − | 2706 | 901 | 901 | 99.9 | R249H |
| M109L | VACV A19L | 105,676 | 105,897 | − | 222 | 73 | 73 | 98.6 | G6E |
| M116L | IMV membrane protein | 113,227 | 113,649 | − | 423 | 140 | 140 | 99.3 | A139T |
| M127L | Photolyase | 119,653 | 120,990 | − | 1338 | 445 | 445 | 99.6 | R207H, A257P |
| M138L | Alpha-2,3 sialyltransferase | 133,811 | 134,683 | − | 873 | 290 | 290 | 99.7 | A15T |
| M147R | Ser/Thr protein kinase | 140,637 | 141,503 | + | 867 | 288 | 288 | 100.0 | H162Y |
| M148R | Putative E3 Ub ligase | 141,564 | 143,591 | + | 2 028 | 675 | 675 | 99.9 | P643S |
| M150R | E3 Ub ligase; NF-κB inhibition | 145,129 | 146,613 | + | 1485 | 494 | 494 | 99.8 | P173S |
| M152R | SERP-3 | 147,626 | 148,426 | + | 801 | 266 | 266 | 99.6 | R218C |
| M153R | E3 Ub ligase/MHC-1 downregulation | 148,464 | 149,075 | + | 612 | 203 | 206 | 98.5 | Δ176–178 |
| M156R | Interferon resistance; elF2α homologue | 150,007 | 150,234 | + | 228 | 75 | 102 | 73.5 | short form |
| M005R | E3 Ub ligase | 155,320 | 156,771 | + | 1 452 | 483 | 483 | 99.8 | V320A |
| M002R | Tumor necrosis factor receptor (TNF-R) homologue | 158,991 | 159,971 | + | 981 | 326 | 326 | 99.7 | A131V, E224K |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossini, E.; Bazzucchi, M.; Trocchi, V.; Merzoni, F.; Bertasio, C.; Knauf, S.; Lavazza, A.; Cavadini, P. Identification and Characterisation of a Myxoma Virus Detected in the Italian Hare (Lepus corsicanus). Viruses 2024, 16, 437. https://doi.org/10.3390/v16030437
Rossini E, Bazzucchi M, Trocchi V, Merzoni F, Bertasio C, Knauf S, Lavazza A, Cavadini P. Identification and Characterisation of a Myxoma Virus Detected in the Italian Hare (Lepus corsicanus). Viruses. 2024; 16(3):437. https://doi.org/10.3390/v16030437
Chicago/Turabian StyleRossini, Elisa, Moira Bazzucchi, Valter Trocchi, Francesca Merzoni, Cristina Bertasio, Sascha Knauf, Antonio Lavazza, and Patrizia Cavadini. 2024. "Identification and Characterisation of a Myxoma Virus Detected in the Italian Hare (Lepus corsicanus)" Viruses 16, no. 3: 437. https://doi.org/10.3390/v16030437