
Utilizing Molecular Epidemiology and Citizen Science for the Surveillance of Lagoviruses in Australia
 , , , , , , , , and
, , , , , , , , and 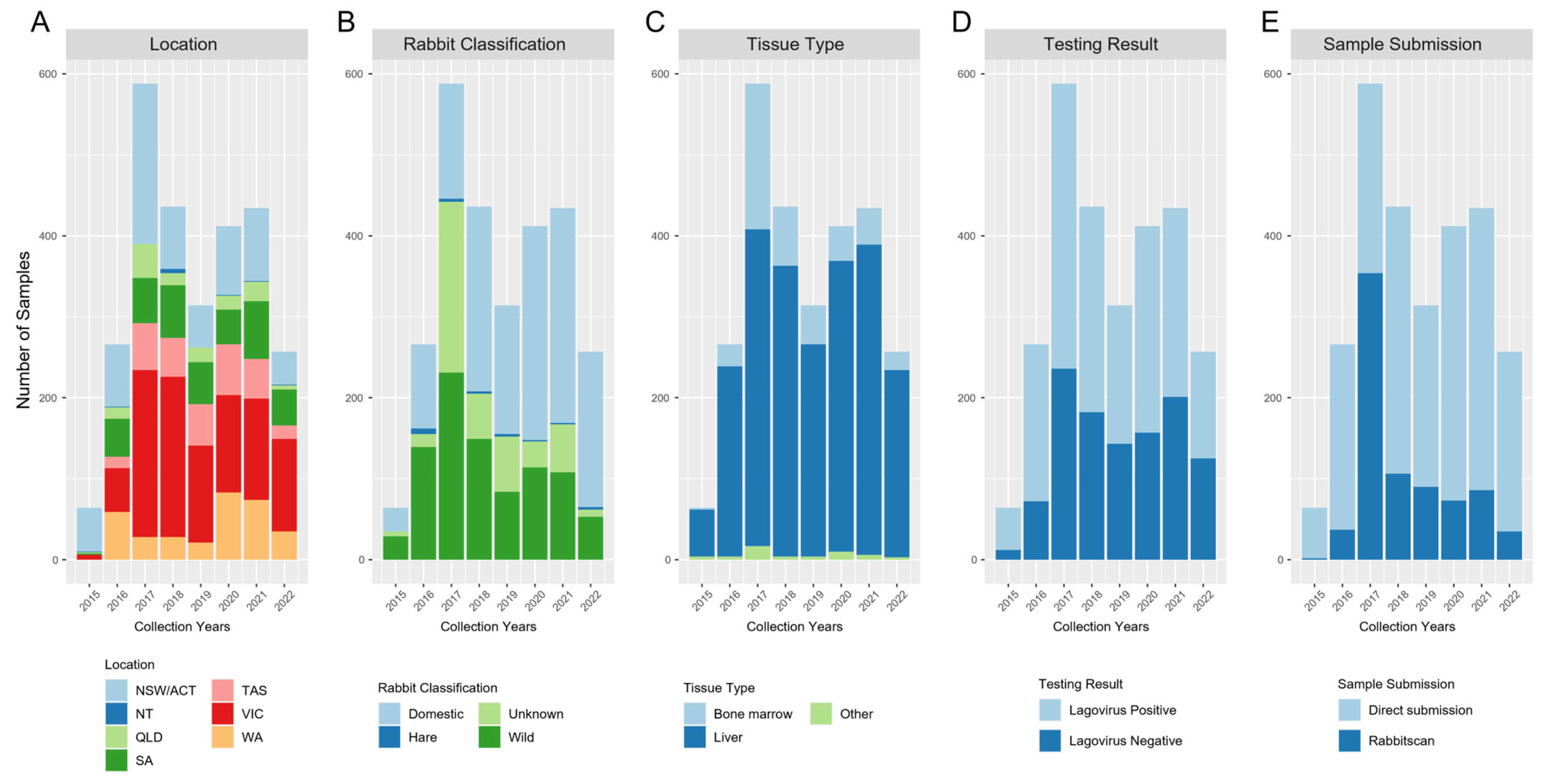{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Sample Collection and RNA Extraction
2.2. RT-qPCR and Variant Identification
2.3. Sequencing
2.4. Sequence Data Analysis
2.5. Bayesian Evolutionary Analyses
2.6. Data Visualization
3. Results
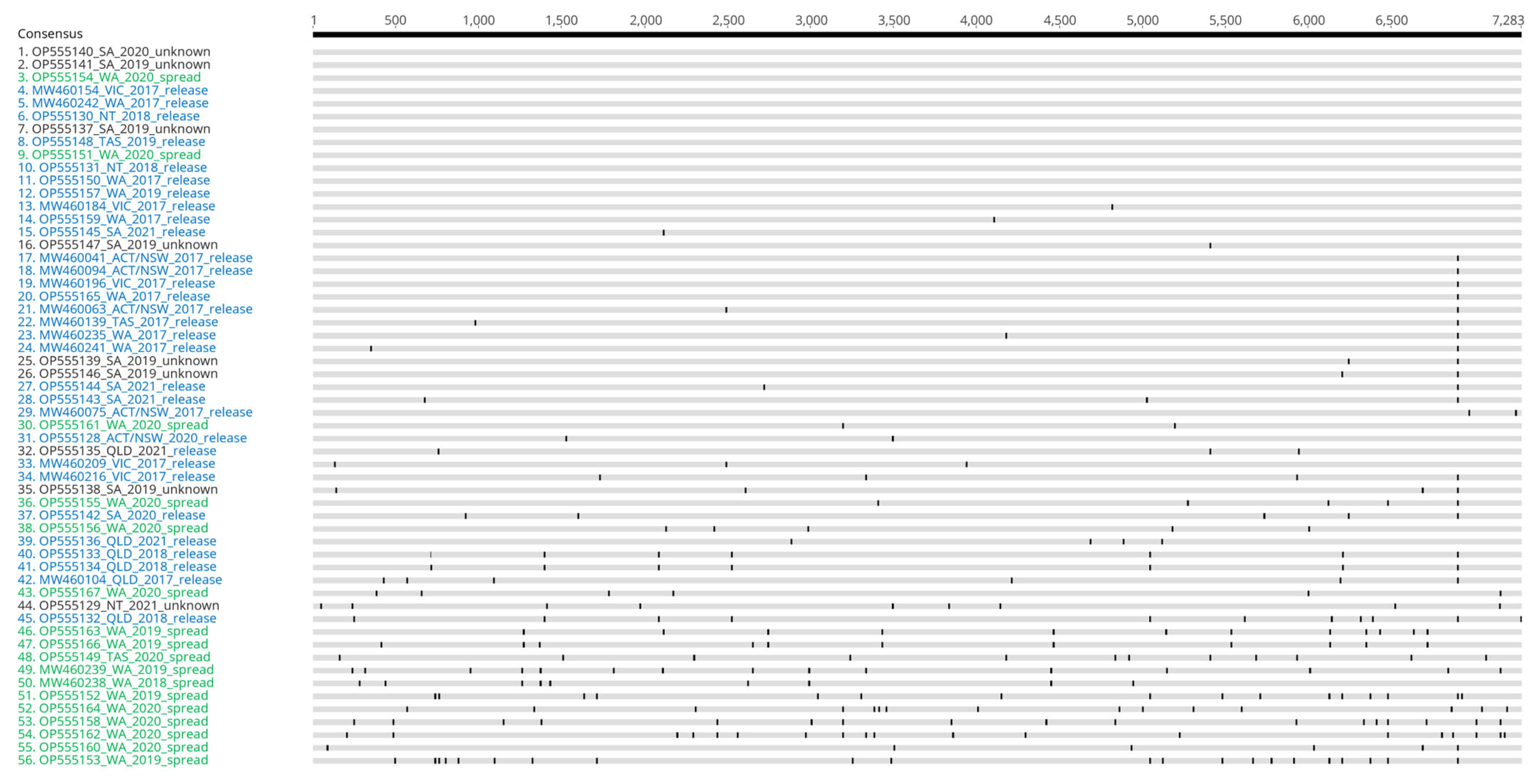
3.1. The Genomic Epidemiology of Lagoviruses Is Improved through Citizen Science
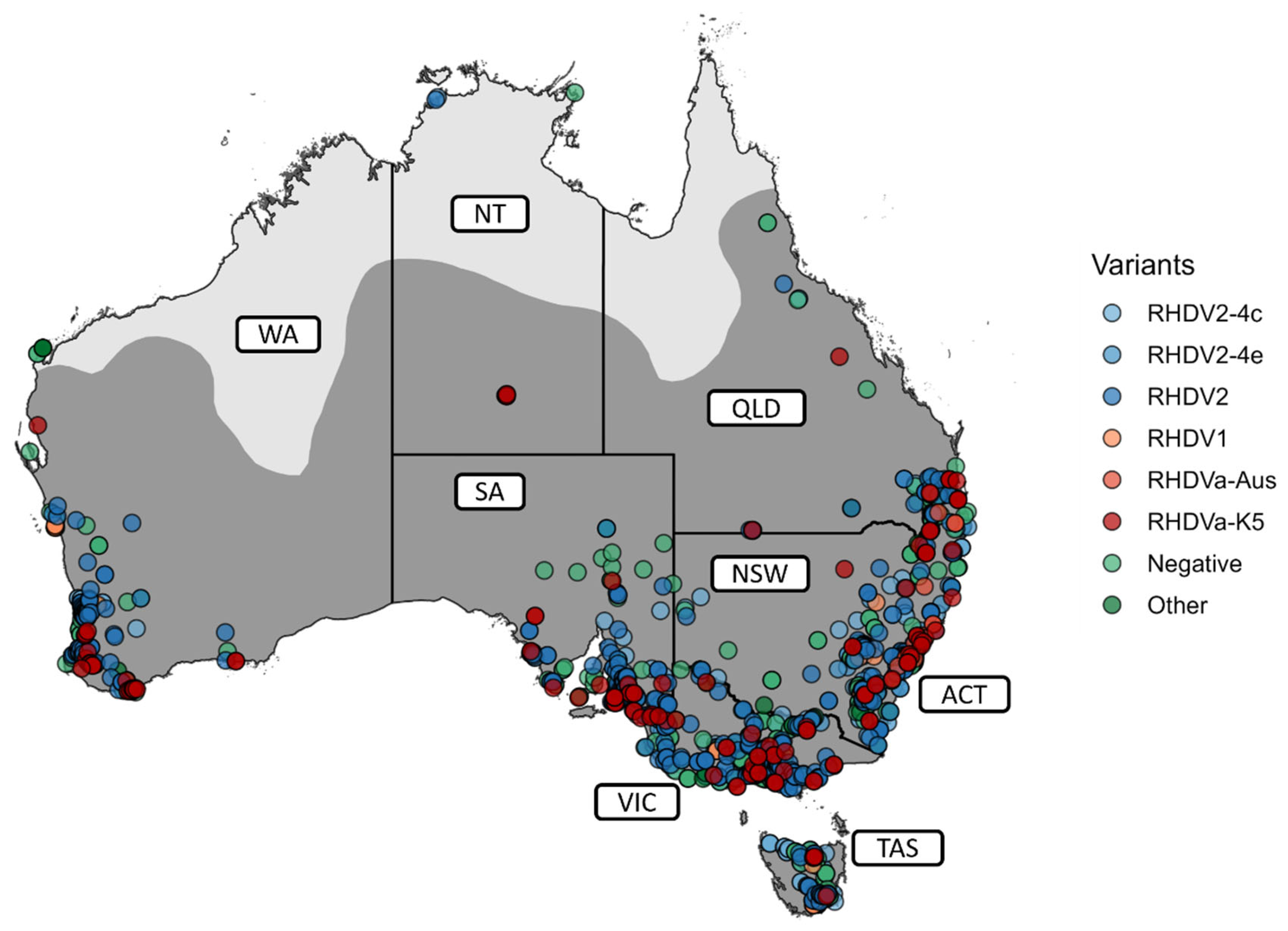
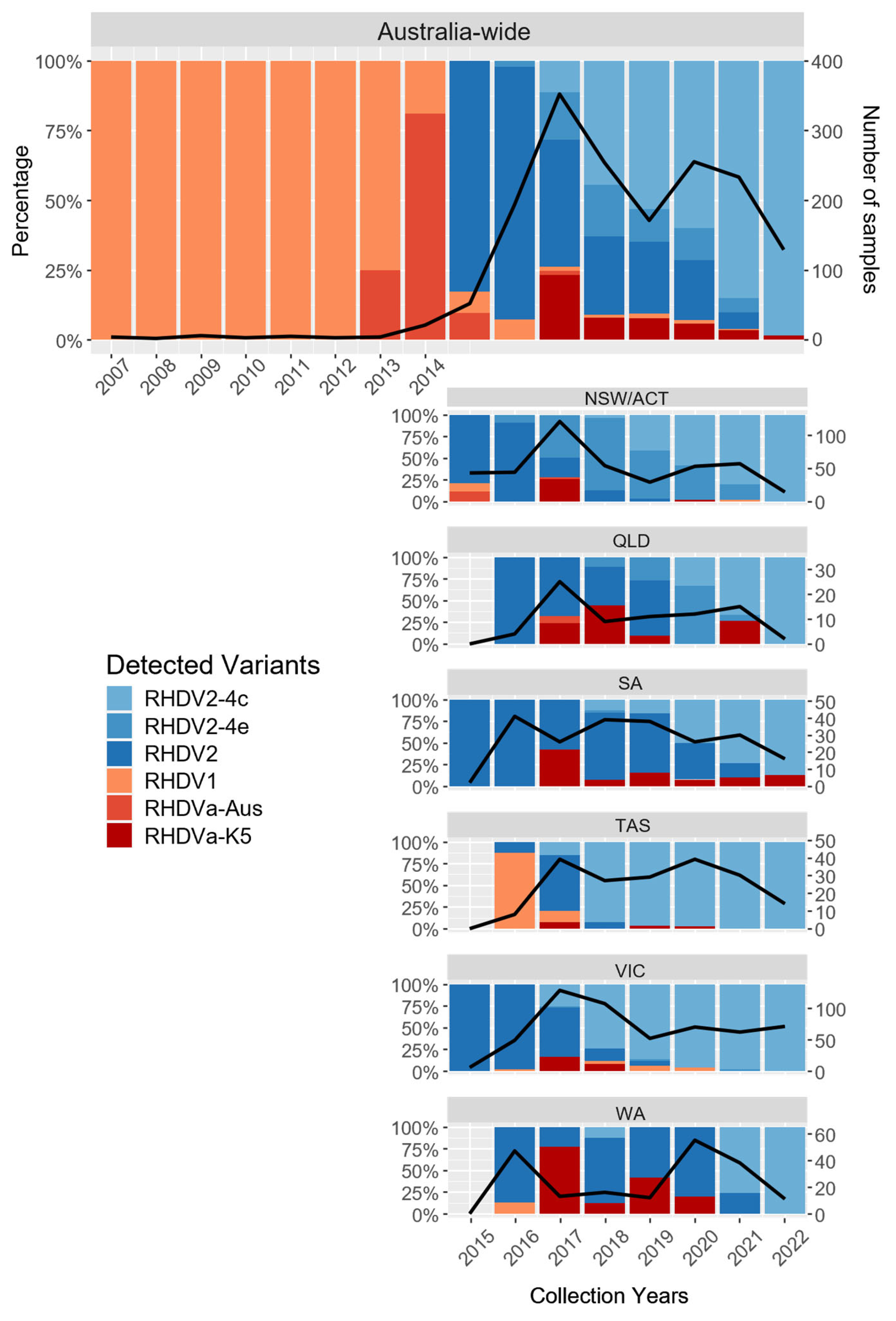
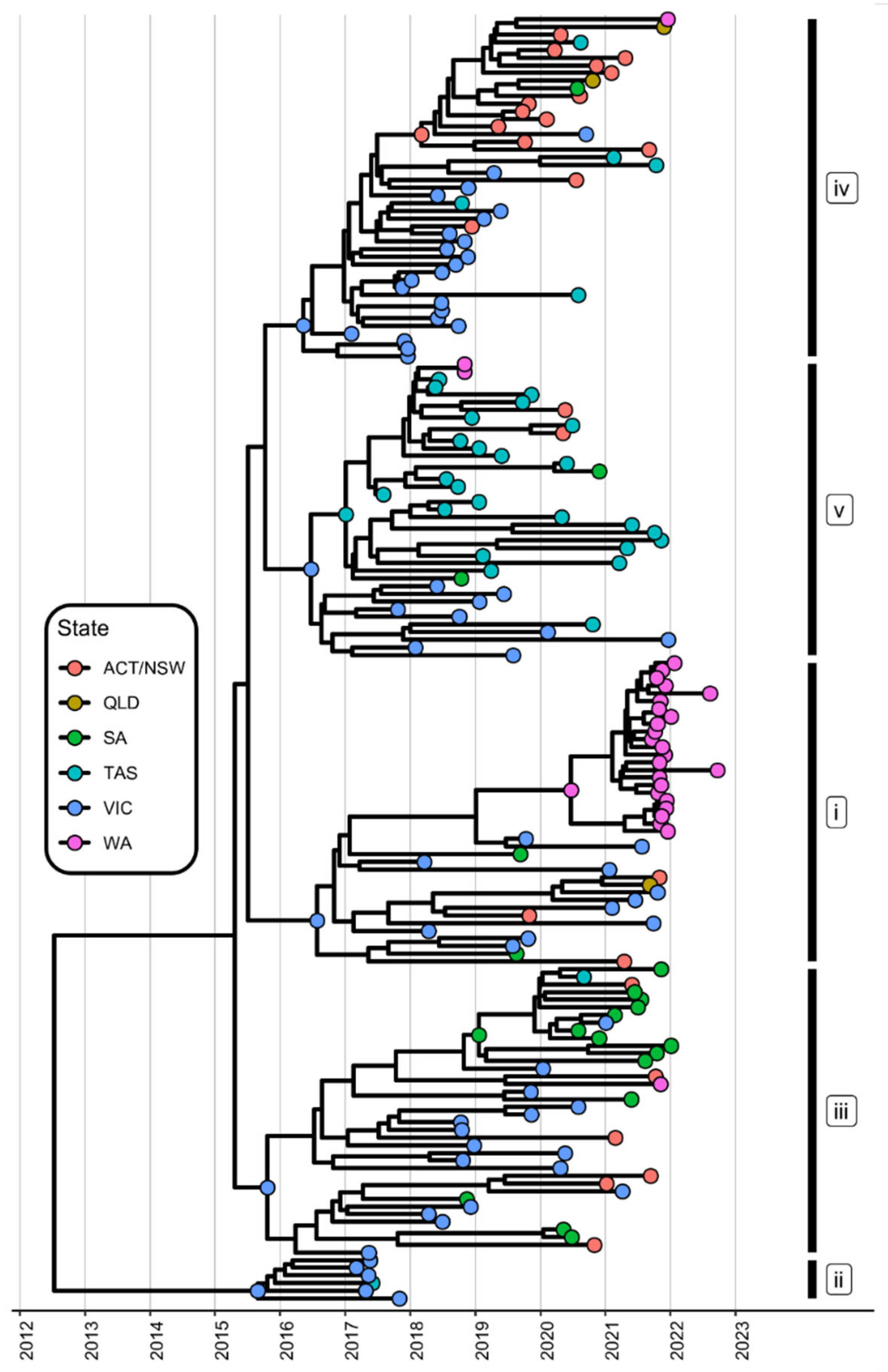
3.2. The Diversity of Lagoviruses Has Been Severely Reduced with the Emergence of RHDV2-4c
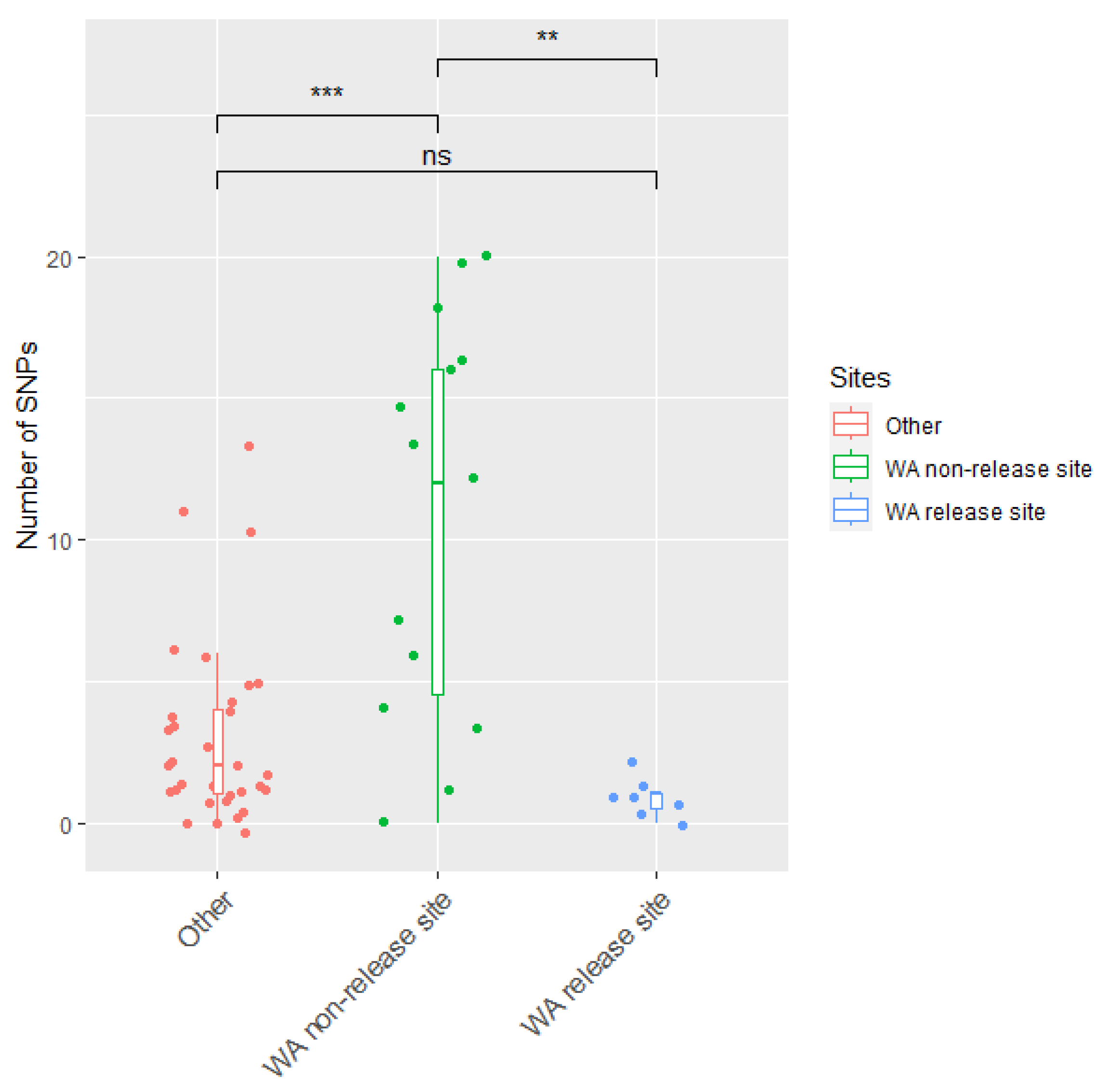
3.3. The Delayed Emergence of RHDV2-4c in Western Australia
3.4. The Circulation of RHDVa-K5 in Western Australia Following Releases
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gong, W.; Sinden, J.; Braysher, M.; Jones, R. The Economic Impacts of Vertebrate Pests in Australia; Invasive Animals Cooperative Research Centre: Canberra, ACT, Australia, 2009. [Google Scholar]
- Cooke, B.; Chudleigh, P.; Simpson, S.; Saunders, G. The Economic Benefits of the Biological Control of Rabbits in Australia, 1950–2011. Aust. Econ. Hist. Rev. 2013, 53, 91–107. [Google Scholar] [CrossRef]
- Cox, T.E.; Ramsey, D.S.L.; Sawyers, E.; Campbell, S.; Matthews, J.; Elsworth, P. The impact of RHDV-K5 on rabbit populations in Australia: An evaluation of citizen science surveys to monitor rabbit abundance. Sci. Rep. 2019, 9, 15229. [Google Scholar] [CrossRef]
- Elsworth, P.; Berman, D.; Brennan, M. Changes in small native animal populations following control of European rabbits (Oryctolagus cuniculus) by warren ripping in the Australian arid zone. Wildl. Res. 2019, 46, 343–354. [Google Scholar] [CrossRef]
- McLeod, R. PESTSMART: Cost of Pest Animals in NSW and Australia, 2013–2014; eSYS Development Pty Ltd.: Sydney, NSW, Australia, 2016. [Google Scholar]
- Bradshaw, C.J.A.; Hoskins, A.J.; Haubrock, P.J.; Cuthbert, R.N.; Diagne, C.; Leroy, B.; Andrews, L.; Page, B.; Cassey, P.; Sheppard, A.W.; et al. Detailed assessment of the reported economic costs of invasive species in Australia. NeoBiota 2021, 67, 511–550. [Google Scholar] [CrossRef]
- CSIRO. CSIRO Future Industries, Rabbit Biocontrol. Available online: https://www.csiro.au/en/about/corporate-governance/ensuring-our-impact/impact-case-studies/future-industries/rabbit-biocontrol (accessed on 23 October 2023).
- Park, J.H.; Lee, Y.S.; Itakura, C. Pathogenesis of acute necrotic hepatitis in rabbit hemorrhagic disease. Lab. Anim. Sci. 1995, 45, 445–449. [Google Scholar]
- Abrantes, J.; van der Loo, W.; Le Pendu, J.; Esteves, P.J. Rabbit haemorrhagic disease (RHD) and rabbit haemorrhagic disease virus (RHDV): A review. Vet. Res. 2012, 43, 12. [Google Scholar] [CrossRef]
- Ohlinger, V.F.; Haas, B.; Meyers, G.; Weiland, F.; Thiel, H.J. Identification and characterization of the virus causing rabbit hemorrhagic disease. J. Virol. 1990, 64, 3331–3336. [Google Scholar] [CrossRef]
- Alonso, J.M.M.; Casais, R.; Boga, J.A.; Parra, F. Processing of rabbit hemorrhagic disease virus polyprotein. J. Virol. 1996, 70, 1261–1265. [Google Scholar] [CrossRef]
- Meyers, G.; Wirblich, C.; Thiel, H.J.; Thumfart, J.O. Rabbit hemorrhagic disease virus: Genome organization and polyprotein processing of a calicivirus studied after transient expression of cDNA constructs. Virology 2000, 276, 349–363. [Google Scholar] [CrossRef]
- Wirblich, C.; Thiel, H.J.; Meyers, G. Genetic map of the calicivirus rabbit hemorrhagic disease virus as deduced from in vitro translation studies. J. Virol. 1996, 70, 7974–7983. [Google Scholar] [CrossRef]
- Sosnovtsev, S.V.; Belliot, G.; Chang, K.-O.; Onwudiwe, O.; Green, K.Y. Feline calicivirus VP2 is essential for the production of infectious virions. J. Virol. 2005, 79, 4012–4024. [Google Scholar] [CrossRef] [PubMed]
- Conley, M.J.; McElwee, M.; Azmi, L.; Gabrielsen, M.; Byron, O.; Goodfellow, I.G.; Bhella, D. Calicivirus VP2 forms a portal-like assembly following receptor engagement. Nature 2019, 565, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Meyers, G.; Wirblich, C.; Thiel, H.J. Rabbit hemorrhagic disease virus--molecular cloning and nucleotide sequencing of a calicivirus genome. Virology 1991, 184, 664–676. [Google Scholar] [CrossRef] [PubMed]
- Meyers, G.; Wirblich, C.; Thiel, H.J. Genomic and subgenomic RNAs of rabbit hemorrhagic disease virus are both protein-linked and packaged into particles. Virology 1991, 184, 677–686. [Google Scholar] [CrossRef]
- Lopes, A.M.; Dalton, K.P.; Magalhaes, M.J.; Parra, F.; Esteves, P.J.; Holmes, E.C.; Abrantes, J. Full genomic analysis of new variant rabbit hemorrhagic disease virus revealed multiple recombination events. J. Gen. Virol. 2015, 96, 1309–1319. [Google Scholar] [CrossRef]
- Mahar, J.E.; Jenckel, M.; Huang, N.; Smertina, E.; Holmes, E.C.; Strive, T.; Hall, R.N. Frequent intergenotypic recombination between the non-structural and structural genes is a major driver of epidemiological fitness in caliciviruses. Virus Evol. 2021, 7, veab080. [Google Scholar] [CrossRef]
- Le Pendu, J.; Abrantes, J.; Bertagnoli, S.; Guitton, J.S.; Le Gall-Recule, G.; Lopes, A.M.; Marchandeau, S.; Alda, F.; Almeida, T.; Celio, A.P.; et al. Proposal for a unified classification system and nomenclature of lagoviruses. J. Gen. Virol. 2017, 98, 1658–1666. [Google Scholar] [CrossRef]
- Hall, R.N.; Mahar, J.E.; Haboury, S.; Stevens, V.; Holmes, E.C.; Strive, T. Emerging Rabbit Hemorrhagic Disease Virus 2 (RHDVb), Australia. Emerg. Infect. Dis. 2015, 21, 2276–2278. [Google Scholar] [CrossRef]
- Hall, R.N.; Mahar, J.E.; Read, A.J.; Mourant, R.; Piper, M.; Huang, N.; Strive, T. A strain-specific multiplex RT-PCR for Australian rabbit haemorrhagic disease viruses uncovers a new recombinant virus variant in rabbits and hares. Transbound. Emerg. Dis. 2018, 65, e444–e456. [Google Scholar] [CrossRef]
- Gould, A.R.; Kattenbelt, J.A.; Lenghaus, C.; Morrissy, C.; Chamberlain, T.; Collins, B.J.; Westbury, H.A. The complete nucleotide sequence of rabbit haemorrhagic disease virus (Czech strain V351): Use of the polymerase chain reaction to detect replication in Australian vertebrates and analysis of viral population sequence variation. Virus Res. 1997, 47, 7–17. [Google Scholar] [CrossRef]
- Mutze, G.; Cooke, B.; Alexander, P. The initial impact of rabbit hemorrhagic disease on European rabbit populations in South Australia. J. Wildl. Dis. 1998, 34, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Kovaliski, J. Monitoring the spread of rabbit hemorrhagic disease virus as a new biological agent for control of wild European rabbits in Australia. J. Wildl. Dis. 1998, 34, 421–428. [Google Scholar] [CrossRef]
- Cooke, B.D.; Fenner, F. Rabbit haemorrhagic disease and the biological control of wild rabbits, Oryctolagus cuniculus, in Australia and New Zealand. Wildl. Res. 2002, 29, 689–706. [Google Scholar] [CrossRef]
- Mutze, G.; Kovaliski, J.; Butler, K.; Capucci, L.; McPhee, S. The effect of rabbit population control programmes on the impact of rabbit haemorrhagic disease in south-eastern Australia. J. Appl. Ecol. 2010, 47, 1137–1146. [Google Scholar] [CrossRef]
- Kovaliski, J.; Sinclair, R.; Mutze, G.; Peacock, D.; Strive, T.; Abrantes, J.; Esteves, P.J.; Holmes, E.C. Molecular epidemiology of Rabbit Haemorrhagic Disease Virus in Australia: When one became many. Mol. Ecol. 2014, 23, 408–420. [Google Scholar] [CrossRef]
- Mahar, J.E.; Read, A.J.; Gu, X.; Urakova, N.; Mourant, R.; Piper, M.; Haboury, S.; Holmes, E.C.; Strive, T.; Hall, R.N. Detection and Circulation of a Novel Rabbit Hemorrhagic Disease Virus in Australia. Emerg. Infect. Dis. 2018, 24, 22–31. [Google Scholar] [CrossRef]
- Taggart, P.L.; O’Connor, T.W.; Cooke, B.; Read, A.J.; Kirkland, P.D.; Sawyers, E.; West, P.; Patel, K. Good intentions with adverse outcomes when conservation and pest management guidelines are ignored: A case study in rabbit biocontrol. Conserv. Sci. Pr. Pract. 2022, 4, e12639. [Google Scholar] [CrossRef]
- Perry, T.; Stenhouse, A.; Wilson, I.; Perfetto, I.; McKelvey, M.W.; Coulson, M.; Ankeny, R.A.; Rismiller, P.D.; Grutzner, F. EchidnaCSI: Engaging the public in research and conservation of the short-beaked echidna. Proc. Natl. Acad. Sci. USA 2022, 119, e2108826119. [Google Scholar] [CrossRef]
- Moulin, N. When Citizen Science highlights alien invasive species in France: The case of Indochina mantis, Hierodula patellifera (Insecta, Mantodea, Mantidae). Biodivers. Data J. 2020, 8, e46989. [Google Scholar] [CrossRef]
- Lehtiniemi, M.; Outinen, O.; Puntila-Dodd, R. Citizen science provides added value in the monitoring for coastal non-indigenous species. J. Env. Environ. Manag. 2020, 267, 110608. [Google Scholar] [CrossRef]
- Eritja, R.; Delacour-Estrella, S.; Ruiz-Arrondo, I.; Gonzalez, M.A.; Barcelo, C.; Garcia-Perez, A.L.; Lucientes, J.; Miranda, M.A.; Bartumeus, F. At the tip of an iceberg: Citizen science and active surveillance collaborating to broaden the known distribution of Aedes japonicus in Spain. Parasites Vectors 2021, 14, 375. [Google Scholar] [CrossRef] [PubMed]
- Hubner, S.; Sisou, D.; Mandel, T.; Todesco, M.; Matzrafi, M.; Eizenberg, H. Wild sunflower goes viral: Citizen science and comparative genomics allow tracking the origin and establishment of invasive sunflower in the Levant. Mol. Ecol. 2022, 31, 2061–2072. [Google Scholar] [CrossRef]
- Carney, R.M.; Mapes, C.; Low, R.D.; Long, A.; Bowser, A.; Durieux, D.; Rivera, K.; Dekramanjian, B.; Bartumeus, F.; Guerrero, D.; et al. Integrating Global Citizen Science Platforms to Enable Next-Generation Surveillance of Invasive and Vector Mosquitoes. Insects 2022, 13, 675. [Google Scholar] [CrossRef] [PubMed]
- Van Haeften, S.; Milic, A.; Addison-Smith, B.; Butcher, C.; Davies, J.M. Grass Gazers: Using citizen science as a tool to facilitate practical and online science learning for secondary school students during the COVID-19 lockdown. Ecol. Evol. 2021, 11, 3488–3500. [Google Scholar] [CrossRef] [PubMed]
- Eden, J.S.; Kovaliski, J.; Duckworth, J.A.; Swain, G.; Mahar, J.E.; Strive, T.; Holmes, E.C. Comparative Phylodynamics of Rabbit Hemorrhagic Disease Virus in Australia and New Zealand. J. Virol. 2015, 89, 9548–9558. [Google Scholar] [CrossRef] [PubMed]
- Mahar, J.E.; Hall, R.N.; Peacock, D.; Kovaliski, J.; Piper, M.; Mourant, R.; Huang, N.; Campbell, S.; Gu, X.; Read, A.; et al. Rabbit Hemorrhagic Disease Virus 2 (RHDV2; GI.2) Is Replacing Endemic Strains of RHDV in the Australian Landscape within 18 Months of Its Arrival. J. Virol. 2018, 92, e01374-17. [Google Scholar] [CrossRef] [PubMed]
- Elsworth, P.; Cooke, B.D.; Kovaliski, J.; Sinclair, R.; Holmes, E.C.; Strive, T. Increased virulence of rabbit haemorrhagic disease virus associated with genetic resistance in wild Australian rabbits (Oryctolagus cuniculus). Virology 2014, 464–465, 415–423. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Rambaut, A.; Lam, T.T.; Max Carvalho, L.; Pybus, O.G. Exploring the temporal structure of heterochronous sequences using TempEst (formerly Path-O-Gen). Virus Evol. 2016, 2, vew007. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Wickham, H.; François, R.; Henry, L.; Müller, K.; Vaughan, D. dplyr: A Grammar of Data Manipulation, R Package Version 1.1.2; The R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Wickham, H.; Bryan, J. readxl: Read Excel Files, R Package Version 1.4.2; The R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Wickham, H.; Henry, L.; Pedersen, T.L.; Luciani, T.J.; Decorde, M.; Lise, V. svglite: An ‘SVG’ Graphics Device, R Package Version 2.1.1; The R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Kassambara, A. ggpubr: ‘ggplot2’ Based Publication Ready Plots, R Package Version 0.6.0; The R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Neuwirth, E. RColorBrewer: ColorBrewer Palettes; RStudio, Inc.: Vienna, Austria, 2022. [Google Scholar]
- Bivand, R.; Keitt, T.; Rowlingson, B. rgdal: Bindings for the ‘Geospatial’ Data Abstraction Library, R Package Version 1.6-7; The R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Pebesma, E.; Bivand, R. Spatial Data Science: With Applications in R; Chapman and Hall/CRC: Boca Raton, FL, USA, 2023. [Google Scholar] [CrossRef]
- Stabler, B. shapefiles: Read and Write ESRI Shapefiles, R Package Version 0.7.2; The R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Hijmans, R.J. terra: Spatial Data Analysis, R Package Version 1.7-39; The R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Hijmans, R.J. raster: Geographic Data Analysis and Modeling, R Package Version 3.6-20; The R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Arnold, J.B. ggthemes: Extra Themes, Scales and Geoms for ‘ggplot2’, R Package Version 4.2.4; The R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Yu, G.; Smith, D.K.; Zhu, H.; Guan, Y.; Lam, T.T.-Y. ggtree: An R package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods Ecol. Evol. 2017, 8, 28–36. [Google Scholar] [CrossRef]
- Wang, L.G.; Lam, T.T.; Xu, S.; Dai, Z.; Zhou, L.; Feng, T.; Guo, P.; Dunn, C.W.; Jones, B.R.; Bradley, T.; et al. Treeio: An R Package for Phylogenetic Tree Input and Output with Richly Annotated and Associated Data. Mol. Biol. Evol. 2020, 37, 599–603. [Google Scholar] [CrossRef]
- Roy-Dufresne, E.; Saltre, F.; Cooke, B.D.; Mellin, C.; Mutze, G.; Cox, T.; Fordham, D.A. Modeling the distribution of a wide-ranging invasive species using the sampling efforts of expert and citizen scientists. Ecol. Evol. 2019, 9, 11053–11063. [Google Scholar] [CrossRef]
- Ramsey, D.S.L.; Cox, T.; Strive, T.; Forsyth, D.M.; Stuart, I.; Hall, R.; Elsworth, P.; Campbell, S. Emerging RHDV2 suppresses the impact of endemic and novel strains of RHDV on wild rabbit populations. J. Appl. Ecol. 2020, 57, 630–641. [Google Scholar] [CrossRef]
- Henning, J.; Meers, J.; Davies, P.R.; Morris, R.S. Survival of rabbit haemorrhagic disease virus (RHDV) in the environment. Epidemiol. Infect. 2005, 133, 719–730. [Google Scholar] [CrossRef]
- Dorji, T.; Jayasingha Ellakkala Appuhamilage, R.M.J.; Bird, P.L.; Huang, N.; O’Connor, T.W.; Patel, K.K.; Strive, T.; Taggart, P.L. Optimising the Delivery of RHDV to Rabbits for Biocontrol: An Experimental Evaluation of Two Novel Methods of Virus Delivery. Viruses 2023, 15, 1814. [Google Scholar] [CrossRef]
- Australian Bureau of Statistics. National, State and Territory Population. Available online: https://www.abs.gov.au/statistics/people/population/national-state-and-territory-population/sep-2022 (accessed on 30 September 2023).
- Abrantes, J.; Droillard, C.; Lopes, A.M.; Lemaitre, E.; Lucas, P.; Blanchard, Y.; Marchandeau, S.; Esteves, P.J.; Le Gall-Recule, G. Recombination at the emergence of the pathogenic rabbit haemorrhagic disease virus Lagovirus europaeus/GI.2. Sci. Rep. 2020, 10, 14502. [Google Scholar] [CrossRef]
- Szillat, K.P.; Höper, D.; Beer, M.; Konig, P. Full-genome sequencing of German rabbit haemorrhagic disease virus uncovers recombination between RHDV (GI.2) and EBHSV (GII.1). Virus Evol. 2020, 6, veaa080. [Google Scholar] [CrossRef] [PubMed]
- Richardson, B.J.; Phillips, S.; Hayes, R.A.; Sindhe, S.; Cooke, B.D. Aspects of the biology of the European rabbit (Oryctolagus cuniculus) and rabbit haemorrhagic disease virus (RHDV) in coastal eastern Australia. Wildl. Res. 2007, 34, 398–407. [Google Scholar] [CrossRef]
- Ramsey, D.S.; Patel, K.K.; Campbell, S.; Hall, R.N.; Taggart, P.L.; Strive, T. Sustained Impact of RHDV2 on Wild Rabbit Populations across Australia Eight Years after Its Initial Detection. Viruses 2023, 15, 1159. [Google Scholar] [CrossRef] [PubMed]
- Hall, R.N.; Capucci, L.; Matthaei, M.; Esposito, S.; Kerr, P.J.; Frese, M.; Strive, T. An in vivo system for directed experimental evolution of rabbit haemorrhagic disease virus. PLoS ONE 2017, 12, e0173727. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, N.Y.G.; Hall, R.N.; Huang, N.; West, P.; Cox, T.E.; Mahar, J.E.; Mason, H.; Campbell, S.; O’Connor, T.; Read, A.J.; et al. Utilizing Molecular Epidemiology and Citizen Science for the Surveillance of Lagoviruses in Australia. Viruses 2023, 15, 2348. https://doi.org/10.3390/v15122348
Peng NYG, Hall RN, Huang N, West P, Cox TE, Mahar JE, Mason H, Campbell S, O’Connor T, Read AJ, et al. Utilizing Molecular Epidemiology and Citizen Science for the Surveillance of Lagoviruses in Australia. Viruses. 2023; 15(12):2348. https://doi.org/10.3390/v15122348
Chicago/Turabian StylePeng, Nias Y. G., Robyn N. Hall, Nina Huang, Peter West, Tarnya E. Cox, Jackie E. Mahar, Hugh Mason, Susan Campbell, Tiffany O’Connor, Andrew J. Read, and et al. 2023. "Utilizing Molecular Epidemiology and Citizen Science for the Surveillance of Lagoviruses in Australia" Viruses 15, no. 12: 2348. https://doi.org/10.3390/v15122348