
Cholesterol Metabolism in Antigen-Presenting Cells and HIV-1 Trans-Infection of CD4+ T Cells

{kind=link}
{kind=link}
{kind=link}
Abstract
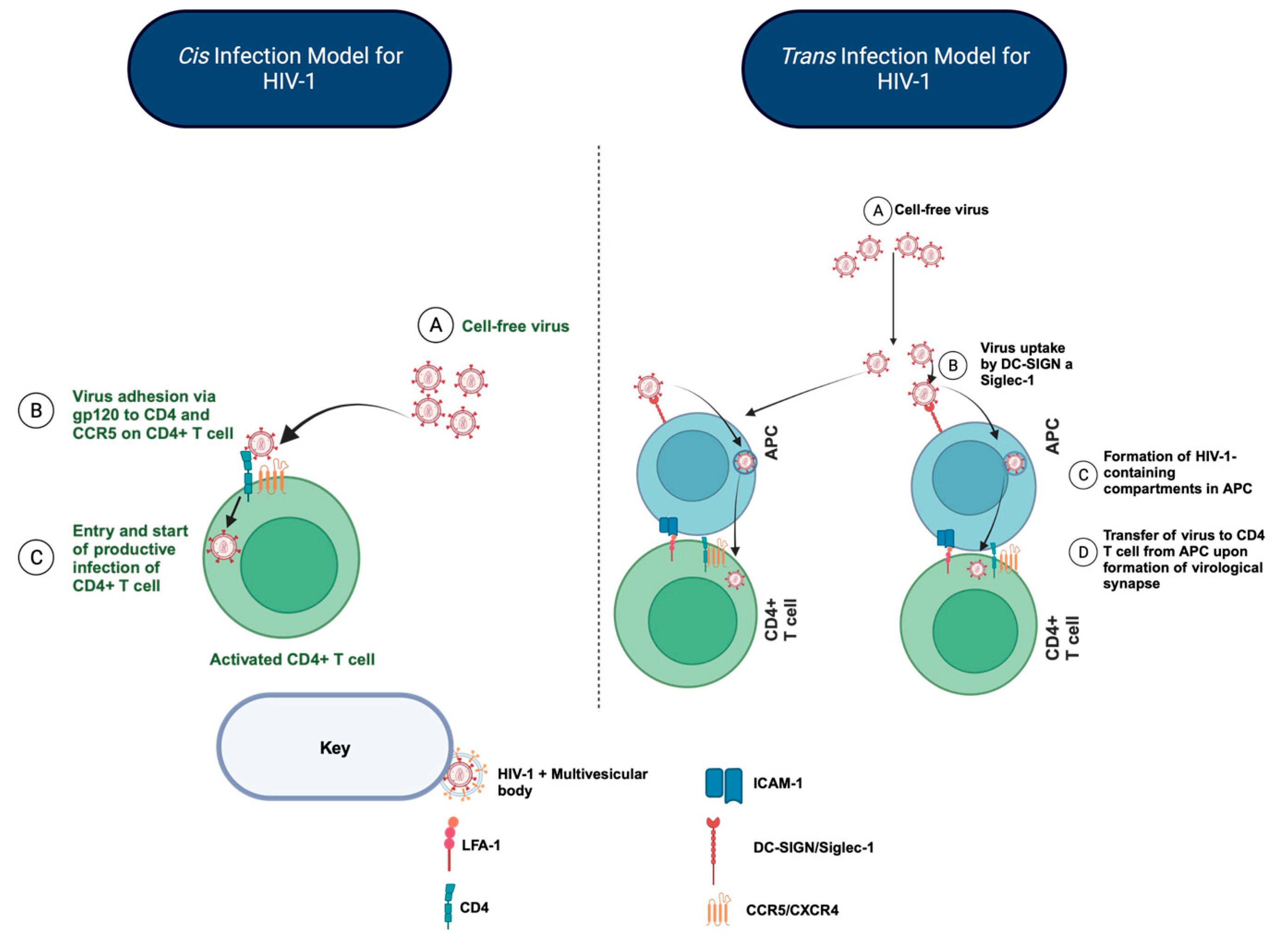
:1. The Importance of HIV-1 Trans-Infection of CD4+ T Cells in Maintaining the HIV-1 Reservoir during Antiretroviral Therapy (ART)
2. Insights Provided by HIV-1 Non-Progressors (NPs) on the Importance of APC-Mediated HIV-1 Trans-Infection of CD4+ T Cell Subsets and the Impact on the Establishment and Maintenance of the HIV-1 Reservoir
3. Cellular Cholesterol Homeostasis: An Intricate, Balanced, and Tightly Controlled Process That, If Perturbed, Can Dysregulate Key Cellular Functions
4. Cholesterol Efflux and HIV-1 Trans-Infection in NPs: Cholesterol Efflux Is One-Half of the Intricate Process of Cholesterol Metabolism for NP APCs Mediating HIV-1 Trans-Infection
5. Conclusions
Funding
Conflicts of Interest
References
- Grant, R.M.; Lama, J.R.; Anderson, P.L.; McMahan, V.; Liu, A.Y.; Vargas, L.; Goicochea, P.; Casapía, M.; Guanira-Carranza, J.V.; Ramirez-Cardich, M.E.; et al. Preexposure chemoprophylaxis for HIV prevention in men who have sex with men. N. Engl. J. Med. 2010, 363, 2587–2599. [Google Scholar] [CrossRef] [PubMed]
- Landovitz, R.J.; Donnell, D.; Clement, M.E.; Hanscom, B.; Cottle, L.; Coelho, L.; Cabello, R.; Chariyalertsak, S.; Dunne, E.F.; Frank, I.; et al. Cabotegravir for HIV Prevention in Cisgender Men and Transgender Women. N. Engl. J. Med. 2021, 385, 595–608. [Google Scholar] [CrossRef]
- Zhong, P.; Agosto, L.M.; Ilinskaya, A.; Dorjbal, B.; Truong, R.; Derse, D.; Uchil, P.D.; Heidecker, G.; Mothes, W. Cell-to-cell transmission can overcome multiple donor and target cell barriers imposed on cell-free HIV. PLoS ONE 2013, 8, e53138. [Google Scholar] [CrossRef] [PubMed]
- Pedro, K.D.; Henderson, A.J.; Agosto, L.M. Mechanisms of HIV-1 cell-to-cell transmission and the establishment of the latent reservoir. Virus Res. 2019, 265, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Bracq, L.; Xie, M.; Benichou, S.; Bouchet, J. Mechanisms for Cell-to-Cell Transmission of HIV-1. Front. Immunol. 2018, 9, 260. [Google Scholar] [CrossRef]
- Dufloo, J.; Bruel, T.; Schwartz, O. HIV-1 cell-to-cell transmission and broadly neutralizing antibodies. Retrovirology 2018, 15, 51. [Google Scholar] [CrossRef]
- Gerberick, A.; Rinaldo, C.R.; Sluis-Cremer, N. Antigen Presenting Cell-Mediated HIV-1 Trans Infection in the Establishment and Maintenance of the Viral Reservoir. Med. Res. Arch. 2023, 11. [Google Scholar] [CrossRef]
- Do, T.; Murphy, G.; Earl, L.A. Three-Dimensional Imaging of HIV-1 Virological Synapses Reveals Membrane Architectures Involved in Virus Transmission. J. Virol. 2017, 91, e01605–e01616. [Google Scholar] [CrossRef]
- Chen, P.; Hübner, W.; Spinelli, M.A.; Chen, B.K. Predominant mode of human immunodeficiency virus transfer between T cells is mediated by sustained Env-dependent neutralization-resistant virological synapses. J. Virol. 2007, 81, 12582–12595. [Google Scholar] [CrossRef]
- Lehman, D.A.; Chung, M.H.; John-Stewart, G.C. HIV-1 persists in breast milk cells despite antiretroviral treatment to prevent mother-to-child transmission. AIDS 2008, 22, 1475–1485. [Google Scholar] [CrossRef]
- Casartelli, N.; Sourisseau, M.; Feldmann, J.; Guivel-Benhassine, F.; Mallet, A. Tetherin Restricts Productive HIV-1 Cell-to-Cell Transmission. PLoS Pathog. 2010, 6, 1000955. [Google Scholar] [CrossRef] [PubMed]
- Jolly, C.; Booth, N.J.; Neil, S.J.D. Cell-cell spread of human immunodeficiency virus type 1 overcomes tetherin/BST-2-mediated restriction in T cells. J. Virol. 2010, 84, 12185–12199. [Google Scholar] [CrossRef]
- Rinaldo, C.R. HIV-1 Trans Infection of CD4+ T Cells by Professional Antigen Presenting Cells. Scientifica 2013, 2013, 164203. [Google Scholar] [CrossRef] [PubMed]
- Rappocciolo, G.; Sluis-Cremer, N.; Rinaldo, C.R. Efficient HIV-1 Trans Infection of CD4(+) T Cells Occurs in the Presence of Antiretroviral Therapy. Open Forum Infect. Dis. 2019, 6, ofz253. [Google Scholar] [CrossRef] [PubMed]
- Rappocciolo, G.; Piazza, P.; Fuller, C.L.; Reinhart, T.A.; Watkins, S.C.; Rowe, T.D.; Jais, M.; Gupta, P.; Rinaldo, C.R. DC-SIGN on B Lymphocytes Is Required for Transmission of HIV-1 to T Lymphocytes. PLoS Pathog. 2006, 2, e70. [Google Scholar] [CrossRef]
- Crocker, P.R.; Paulson, J.C.; Varki, A. Siglecs and their roles in the immune system. Nat. Rev. Immunol. 2007, 7, 255–266. [Google Scholar] [CrossRef]
- Bermejo-Jambrina, M.; Eder, J.; Helgers, L.C.; Hertoghs, N.; Nijmeijer, B.M.; Stunnenberg, M.; Geijtenbeek, T.B.H. C-Type Lectin Receptors in Antiviral Immunity and Viral Escape. Front. Immunol. 2018, 9, 590. [Google Scholar] [CrossRef]
- Pino, M.; Erkizia, I.; Benet, S.; Erikson, E.; Fernández-Figueras, M.T.; Guerrero, D.; Dalmau, J.; Ouchi, D.; Rausell, A.; Ciuffi, A.; et al. HIV-1 immune activation induces Siglec-1 expression and enhances viral trans-infection in blood and tissue myeloid cells. Retrovirology 2015, 12, 37. [Google Scholar] [CrossRef]
- Bromley, S.K.; Burack, W.R.; Johnson, K.G.; Somersalo, K.; Sims, T.N.; Sumen, C.; Davis, M.M.; Shaw, A.S.; Allen, P.M.; Dustin, M.L. The immunological synapse. Annu. Rev. Immunol. 2001, 19, 375–396. [Google Scholar] [CrossRef]
- McDonald, D.; Wu, L.; Bohks, S.M.; KewalRamani, V.N.; Unutmaz, D.; Hope, T.J. Recruitment of HIV and its receptors to dendritic cell-T cell junctions. Science 2003, 300, 1295–1297. [Google Scholar] [CrossRef]
- Gutiérrez-Martínez, E.; Garrabé, S.B.; Mateos, N.; Erkizia, I.; Ander, J.; Nieto-Garai, J.A.; Lorizate, M.; Borgman, K.J.E.; Manzo, C.; Campelo, F.; et al. Actin-regulated Siglec-1 nanoclustering influences HIV-1 capture and virus-containing compartment formation in dendritic cells. eLife 2023, 12, e78836. [Google Scholar] [CrossRef] [PubMed]
- Gerberick, A.; DeLucia, D.C.; Piazza, P.; Alaoui-El-Azher, M.; Rinaldo, C.R.; Sluis-Cremer, N.; Rappocciolo, G. B Lymphocytes, but Not Dendritic Cells, Efficiently HIV-1 Trans Infect Naive CD4+ T Cells: Implications for the Viral Reservoir. Mbio 2021, 12, e02998-20. [Google Scholar] [CrossRef] [PubMed]
- Rappocciolo, G.; Jais, M.; Piazza, P.; Reinhart, T.A.; Berendam, S.J.; Garcia-Exposito, L.; Gupta, P.; Rinaldo, C.R. Alterations in cholesterol metabolism restrict HIV-1 trans infection in nonprogressors. Mbio 2014, 5, e01031-13. [Google Scholar] [CrossRef] [PubMed]
- Walker, B.D.; Yu, X.G. Unravelling the mechanisms of durable control of HIV-1. Nat. Rev. Immunol. 2013, 13, 487–498. [Google Scholar] [CrossRef]
- Jiang, C.; Lian, X.; Gao, C.; Sun, X.; Einkauf, K.B.; Chevalier, J.M.; Chen, S.M.Y.; Hua, S.; Rhee, B.; Chang, K.; et al. Distinct viral reservoirs in individuals with spontaneous control of HIV-1. Nature 2020, 585, 261–267. [Google Scholar] [CrossRef]
- Delucia, D.C.; Rinaldo, C.R.; Rappocciolo, G. Inefficient HIV-1 trans-Infection of CD4 T Cells by Macrophages from HIV-1 Nonprogressors Is Associated with Altered Membrane Cholesterol and DC-SIGN. J. Virol. 2018, 92, e00092-18. [Google Scholar] [CrossRef]
- Casado, C.; Marrero-Hernández, S.; Márquez-Arce, D.; Pernas, M.; Marfil, S.; Borràs-Grañana, F.; Olivares, I.; Cabrera-Rodríguez, R.; Valera, M.S.; de Armas-Rillo, L.; et al. Viral Characteristics Associated with the Clinical Nonprogressor Phenotype Are Inherited by Viruses from a Cluster of HIV-1 Elite Controllers. Mbio 2018, 9, e02338-17. [Google Scholar] [CrossRef]
- Venanzi, R.E.; Pinzone, M.R.; Cannon, L.; Weissman, S.; Ceccarelli, M.; Zurakowski, R.; Nunnari, G.; O’Doherty, U. Persistence of an intact HIV reservoir in phenotypically naive T cells. JCI Insight 2020, 5, e133157. [Google Scholar] [CrossRef]
- Pinzone, M.R.; Weissman, S.; Pasternak, A.O.; Zurakowski, R.; Migueles, S.; O’Doherty, U. Naive infection predicts reservoir diversity and is a formidable hurdle to HIV eradication. JCI Insight 2021, 6, e150794. [Google Scholar] [CrossRef]
- Zerbato, J.M.; McMahon, D.K.; Sobolewski, M.D.; Mellors, J.W.; Sluis-Cremer, N. Naive CD4+ T Cells Harbor a Large Inducible Reservoir of Latent, Replication-competent Human Immunodeficiency Virus Type 1. Clin. Infect. Dis. 2019, 69, 1919–1925. [Google Scholar] [CrossRef]
- Zerbato, J.M.; Serrao, E.; Lenzi, G.; Kim, B.; Ambrose, Z.; Watkins, S.C.; Engelman, A.N.; Sluis-Cremer, N. Establishment and Reversal of HIV-1 Latency in Naive and Central Memory CD4+ T Cells In Vitro. J. Virol. 2016, 90, 8059–8073. [Google Scholar] [CrossRef] [PubMed]
- Buck, M.D.; O’Sullivan, D.; Pearce, E.L. T cell metabolism drives immunity. J. Exp. Med. 2015, 212, 1345–1360. [Google Scholar] [CrossRef] [PubMed]
- Kotas, M.E.; Medzhitov, R. Homeostasis, inflammation, and disease susceptibility. Cell 2015, 160, 816–827. [Google Scholar] [CrossRef] [PubMed]
- Kidani, Y.; Elsaesser, H.; Hock, M.B.; Vergnes, L.; Williams, K.J.; Argus, J.P.; Marbois, B.N.; Komisopoulou, E.; Wilson, E.B.; Osborne, T.F.; et al. Sterol regulatory element-binding proteins are essential for the metabolic programming of effector T cells and adaptive immunity. Nat. Immunol. 2013, 14, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, D.; Perucha, E. Cholesterol metabolism: A new molecular switch to control inflammation. Clin. Sci. 2021, 135, 1389–1408. [Google Scholar] [CrossRef]
- Ruysschaert, J.M.; Lonez, C. Role of lipid microdomains in TLR-mediated signalling. Biochim. Biophys. Acta (BBA)-Biomembr. 2015, 1848, 1860–1867. [Google Scholar] [CrossRef]
- Köberlin, M.S.; Heinz, L.X.; Superti-Furga, G. Functional crosstalk between membrane lipids and TLR biology. Curr. Opin. Cell Biol. 2016, 39, 28–36. [Google Scholar] [CrossRef]
- Brown, M.S.; Goldstein, J.L. A Receptor-Mediated Pathway for Cholesterol Homeostasis. Science 1986, 232, 34–47. [Google Scholar] [CrossRef]
- Pfeffer, S.R. NPC intracellular cholesterol transporter 1 (NPC1)-mediated cholesterol export from lysosomes. J. Biol. Chem. 2019, 294, 1706–1709. [Google Scholar] [CrossRef]
- Charles, K.N.; Shackelford, J.E.; Faust, P.L.; Fliesler, S.J.; Stangl, H.; Kovacs, W.J. Functional Peroxisomes Are Essential for Efficient Cholesterol Sensing and Synthesis. Front. Cell Dev. Biol. 2020, 8, 560266. [Google Scholar] [CrossRef]
- Yu, X.H.; Tang, C.K. ABCA1, ABCG1, and Cholesterol Homeostasis. Adv. Exp. Med. Biol. 2022, 1377, 95–107. [Google Scholar] [PubMed]
- Sakai, J.; Duncan, E.A.; Rawson, R.B.; Hua, X.; Brown, M.S.; Goldstein, J.L. Sterol-Regulated Release of SREBP-2 from Cell Membranes Requires Two Sequential Cleavages, One Within a Transmembrane Segment. Cell 1996, 85, 1037–1046. [Google Scholar] [CrossRef] [PubMed]
- Bauer, R.; Brüne, B.; Schmid, T. Cholesterol metabolism in the regulation of inflammatory responses. Front. Pharmacol. 2023, 14, 1121819. [Google Scholar] [CrossRef]
- Luo, J.; Yang, H.; Song, B.L. Mechanisms and regulation of cholesterol homeostasis. Nat. Rev. Mol. Cell Biol. 2020, 21, 225–245. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; McDonald, J.G.; Patel, A.; Zhang, Y.; Umetani, M.; Xu, F.; Westover, E.J.; Covey, D.F.; Mangelsdorf, D.J.; Cohen, J.C.; et al. Sterol intermediates from cholesterol biosynthetic pathway as liver X receptor ligands. J. Biol. Chem. 2006, 281, 27816–27826. [Google Scholar] [CrossRef] [PubMed]
- Teresa, B.M.; Petillo, S.; Santoni, A.; Cippitelli, M. Liver X Receptors: Regulators of Cholesterol Metabolism, Inflammation, Autoimmunity, and Cancer. Front. Immunol. 2020, 11, 584303. [Google Scholar]
- Tontonoz, P. Transcriptional and Posttranscriptional Control of Cholesterol Homeostasis by Liver X Receptors. Cold Spring Harb. Symp. Quant. Biol. 2011, 76, 129–137. [Google Scholar] [CrossRef]
- Lamerton, R.E.; Lightfoot, A.; Nieves, D.J.; Owen, D.M. The Role of Protein and Lipid Clustering in Lymphocyte Activation. Front. Immunol. 2021, 12, 600961. [Google Scholar] [CrossRef]
- Bunnell, S.C.; Hong, D.I.; Kardon, J.R.; Yamazaki, T.; McGlade, C.J.; Barr, V.A.; Samelson, L.E. T cell receptor ligation induces the formation of dynamically regulated signaling assemblies. J. Cell Biol. 2002, 158, 1263–1275. [Google Scholar] [CrossRef]
- Wang, F.; Beck-García, K.; Zorzin, C.; Schamel, W.W.A.; Davis, M.M. Inhibition of T cell receptor signaling by cholesterol sulfate, a naturally occurring derivative of membrane cholesterol. Nat. Immunol. 2016, 17, 844–850. [Google Scholar] [CrossRef]
- Ashdown, G.W.; Williamson, D.J.; Soh, G.H.M.; Day, N.; Burn, G.L.; Owen, D.M. Membrane lipid order of sub-synaptic T cell vesicles correlates with their dynamics and function. Traffic 2018, 19, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Balagopalan, L.; Yi, J.; Nguyen, T.; McIntire, K.M.; Harned, A.S.; Narayan, K.; Samelson, L.E. Plasma membrane LAT activation precedes vesicular recruitment defining two phases of early T-cell activation. Nat. Commun. 2018, 9, 2013. [Google Scholar] [CrossRef] [PubMed]
- Williamson, D.J.; Owen, D.M.; Rossy, J.; Magenau, A.; Wehrmann, M.; Gooding, J.J.; Gaus, K. Pre-existing clusters of the adaptor Lat do not participate in early T cell signaling events. Nat. Immunol. 2011, 12, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Soares, H.; Henriques, R.; Sachse, M.; Ventimiglia, L.; Alonso, M.A.; Zimmer, C.; Thoulouze, M.I.; Alcover, A. Regulated vesicle fusion generates signaling nanoterritories that control T cell activation at the immunological synapse. J. Exp. Med. 2013, 210, 2415–2433. [Google Scholar] [CrossRef]
- Hanley, T.M.; Viglianti, G.A. Nuclear Receptor Signaling Inhibits HIV-1 Replication in Macrophages through Multiple trans-Repression Mechanisms. J. Virol. 2011, 85, 10834–10850. [Google Scholar] [CrossRef]
- Morrow, M.P.; Grant, A.; Mujawar, Z.; Dubrovsky, L.; Pushkarsky, T.; Kiselyeva, Y.; Jennelle, L.; Mukhamedova, N.; Remaley, A.T.; Kashanchi, F.; et al. Stimulation of the liver X receptor pathway inhibits HIV-1 replication via induction of ATP-binding cassette transporter A1. Mol. Pharmacol. 2010, 78, 215–225. [Google Scholar] [CrossRef]
- Mujawar, Z.; Rose, H.; Morrow, M.P.; Pushkarsky, T.; Dubrovsky, L.; Mukhamedova, N.; Fu, Y.; Dart, A.; Orenstein, J.M.; Bobryshev, Y.V.; et al. Human immunodeficiency virus impairs reverse cholesterol transport from macrophages. PLoS Biol. 2006, 4, e365. [Google Scholar] [CrossRef]
- Sviridov, D.; Mukhamedova, N.; Makarov, A.A.; Adzhubei, A.; Bukrinsky, M. Comorbidities of HIV infection: Role of Nef-induced impairment of cholesterol metabolism and lipid raft functionality. AIDS 2020, 34, 1–13. [Google Scholar] [CrossRef]
- Hanley, T.M.; Blay Puryear, W.; Gummuluru, S.; Viglianti, G.A. PPARγ and LXR Signaling Inhibit Dendritic Cell-Mediated HIV-1 Capture and trans-Infection. PLoS Pathog. 2010, 6, e1000981. [Google Scholar] [CrossRef]
- Saulle, I.B.; Ibba, S.V.; Vittori, C.; Fenizia, C.; Mercurio, V.; Vichi, F.; Caputo, S.L.; Trabattoni, D.; Clerici, M.; Biasin, M. Sterol metabolism modulates susceptibility to HIV-1 Infection. AIDS 2020, 34, 1593–1602. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okpaise, D.; Sluis-Cremer, N.; Rappocciolo, G.; Rinaldo, C.R. Cholesterol Metabolism in Antigen-Presenting Cells and HIV-1 Trans-Infection of CD4+ T Cells. Viruses 2023, 15, 2347. https://doi.org/10.3390/v15122347
Okpaise D, Sluis-Cremer N, Rappocciolo G, Rinaldo CR. Cholesterol Metabolism in Antigen-Presenting Cells and HIV-1 Trans-Infection of CD4+ T Cells. Viruses. 2023; 15(12):2347. https://doi.org/10.3390/v15122347
Chicago/Turabian StyleOkpaise, Daniel, Nicolas Sluis-Cremer, Giovanna Rappocciolo, and Charles R. Rinaldo. 2023. "Cholesterol Metabolism in Antigen-Presenting Cells and HIV-1 Trans-Infection of CD4+ T Cells" Viruses 15, no. 12: 2347. https://doi.org/10.3390/v15122347