
Effectiveness of Bivalent Omicron-Containing Booster Vaccines against SARS-CoV-2 Omicron Variant among Individuals with and without Prior SARS-CoV-2 Infection

Abstract
:1. Introduction
2. Material and Methods
2.1. Study Design
2.2. Participant Recruitment and Sample Collection
2.3. Detection of the SARS-CoV-2-Specific IgM/IgG
2.4. Neutralizing Antibody Assay
2.5. Plaque Reduction Neutralization Test (PRNT)
2.6. Statistical Analysis
3. Results
3.1. Cohort of Study
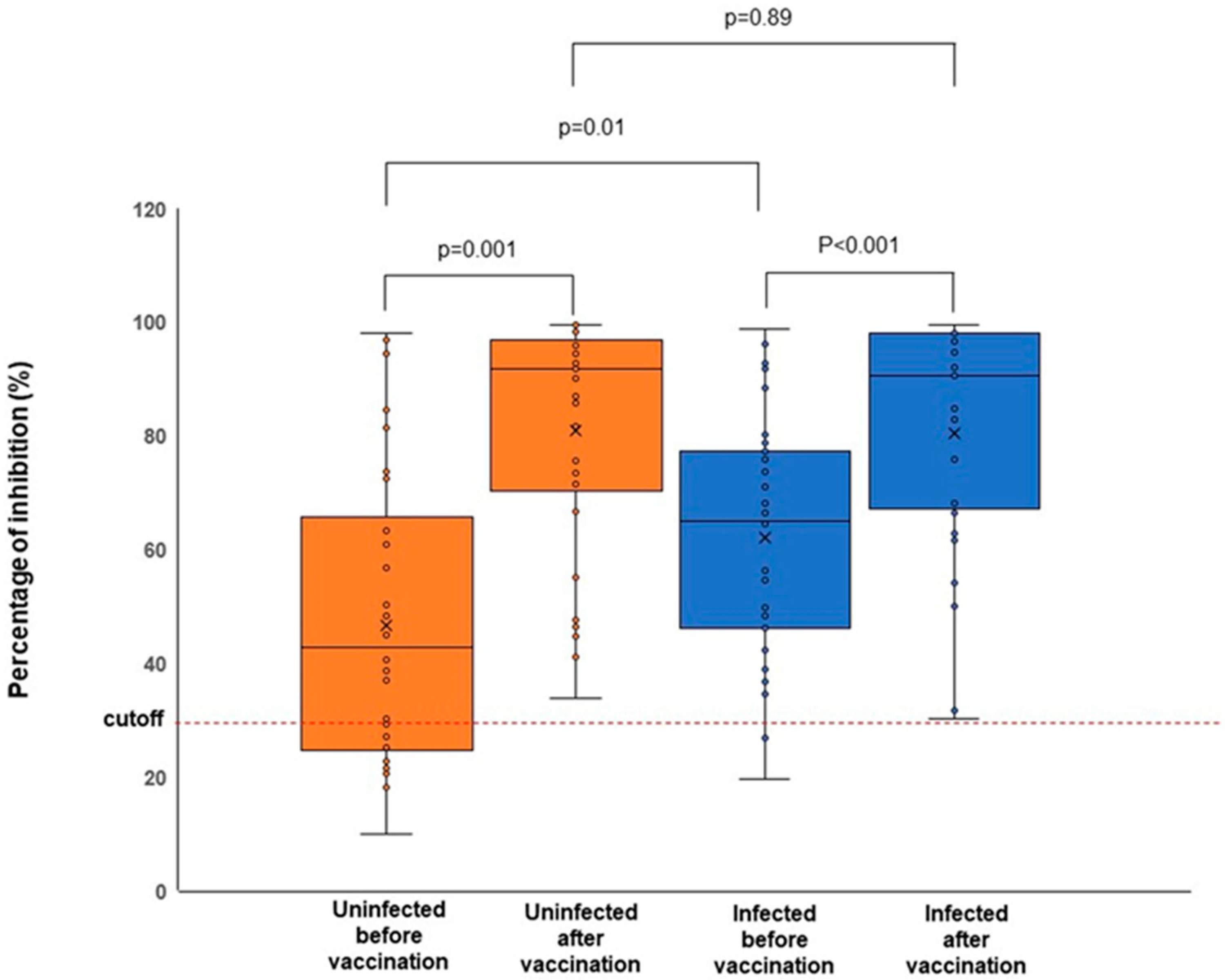
3.2. The SARS-CoV-2 Specific Antibodies Were Detected Regardless of the Infection Status
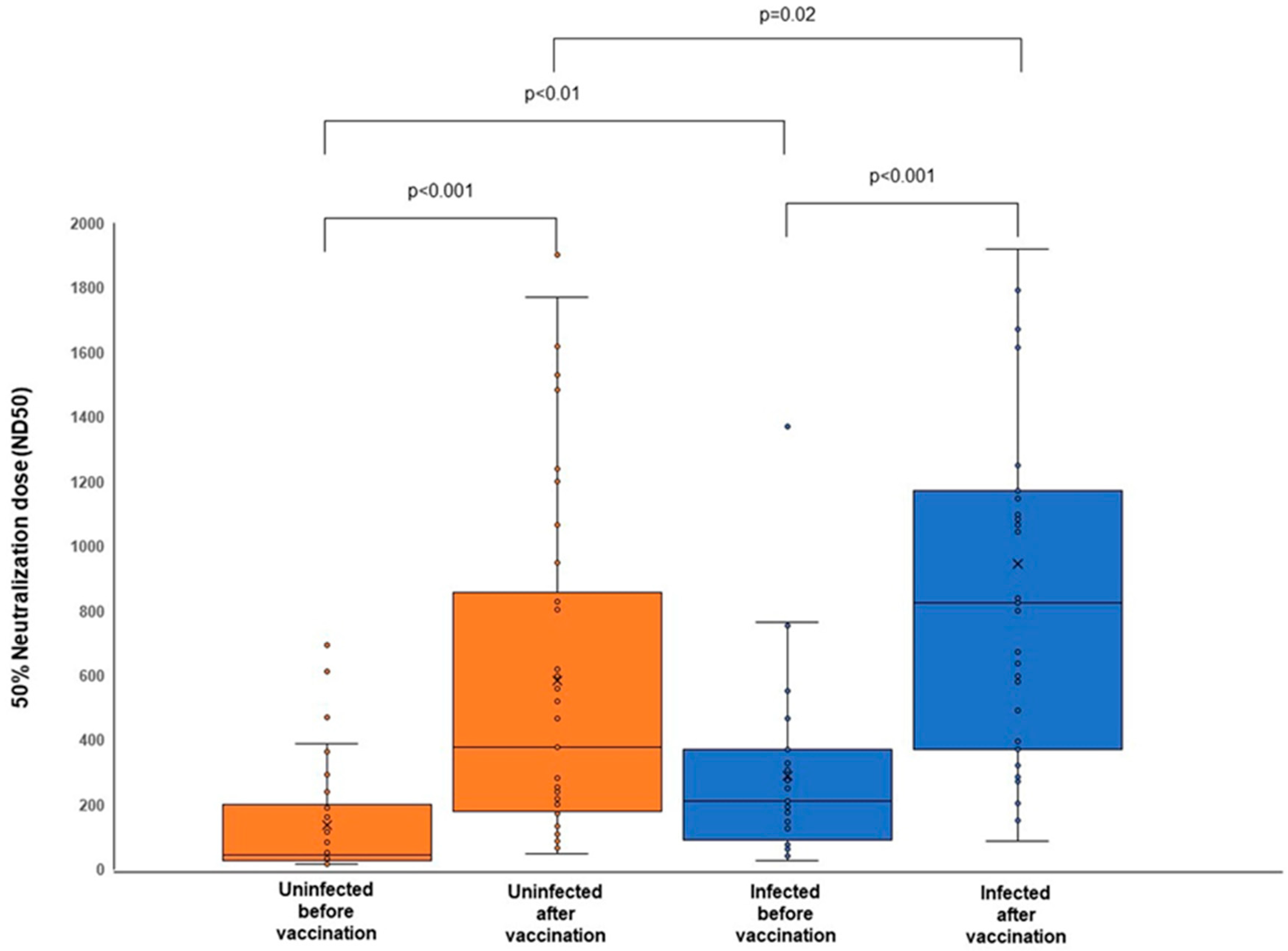
3.3. The Bivalent mRNA COVID-19 Vaccines Boost the Neutralizing Antibody Response
3.4. The Bivalent mRNA COVID-19 Vaccines Induce Neutralizing Capacity against the Omicron Variant
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mohammed, I.; Nauman, A.; Paul, P.; Ganesan, S.; Chen, K.-H.; Jalil, S.M.S.; Jaouni, S.H.; Kawas, H.; Khan, W.A.; Vattoth, A.L. The efficacy and effectiveness of the COVID-19 vaccines in reducing infection, severity, hospitalization, and mortality: A systematic review. Hum. Vaccin. Immunother. 2022, 18, 2027160. [Google Scholar] [CrossRef] [PubMed]
- Krammer, F. SARS-CoV-2 vaccines in development. Nature 2020, 586, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Madhi, S.A.; Baillie, V.; Cutland, C.L.; Voysey, M.; Koen, A.L.; Fairlie, L.; Padayachee, S.D.; Dheda, K.; Barnabas, S.L.; Bhorat, Q.E. Efficacy of the ChAdOx1 nCoV-19 COVID-19 vaccine against the B. 1.351 variant. N. Engl. J. Med. 2021, 384, 1885–1898. [Google Scholar] [CrossRef] [PubMed]
- Gruell, H.; Vanshylla, K.; Tober-Lau, P.; Hillus, D.; Schommers, P.; Lehmann, C.; Kurth, F.; Sander, L.E.; Klein, F. mRNA booster immunization elicits potent neutralizing serum activity against the SARS-CoV-2 Omicron variant. Nat. Med. 2022, 28, 477–480. [Google Scholar] [CrossRef]
- Zeng, B.; Gao, L.; Zhou, Q.; Yu, K.; Sun, F. Effectiveness of COVID-19 vaccines against SARS-CoV-2 variants of concern: A systematic review and meta-analysis. BMC Med. 2022, 20, 200. [Google Scholar] [CrossRef]
- McLean, G.; Kamil, J.; Lee, B.; Moore, P.; Schulz, T.F.; Muik, A.; Sahin, U.; Türeci, Ö.; Pather, S. The impact of evolving SARS-CoV-2 mutations and variants on COVID-19 vaccines. Mbio 2022, 13, e02979-21. [Google Scholar] [CrossRef]
- Dhawan, M.; Sharma, A.; Priyanka, N.; Thakur, N.; Rajkhowa, T.K.; Choudhary, O.P. Delta variant (B. 1.617. 2) of SARS-CoV-2: Mutations, impact, challenges, and possible solutions. Hum. Vaccin. Immunother. 2022, 18, 2068883. [Google Scholar] [CrossRef]
- Tchesnokova, V.; Kulasekara, H.; Larson, L.; Bowers, V.; Rechkina, E.; Kisiela, D.; Sledneva, Y.; Choudhury, D.; Maslova, I.; Deng, K. Acquisition of the L452R mutation in the ACE2-binding interface of Spike protein triggers recent massive expansion of SARS-CoV-2 variants. J. Clin. Microbiol. 2021, 59, e00921. [Google Scholar] [CrossRef]
- Viana, R.; Moyo, S.; Amoako, D.G.; Tegally, H.; Scheepers, C.; Althaus, C.L.; Anyaneji, U.J.; Bester, P.A.; Boni, M.F.; Chand, M. Rapid epidemic expansion of the SARS-CoV-2 Omicron variant in southern Africa. Nature 2022, 603, 679–686. [Google Scholar] [CrossRef]
- The Economic Online. Scientists Say the Omicron Sub-Lineage BA.1 Replacing the Delta Variant. 2022. Available online: https://economictimes.indiatimes.com/news/india/scientists-say-omicron-sub-lineage-ba-1-replacing-delta-variant/articleshow/88806759.cms?from=mdr (accessed on 13 August 2023).
- The Guardian. Why Are There So Many Omicron Subvariants and Why Have BA.4 and BA.5 Taken Hold? 2022. Available online: https://www.theguardian.com/australia-news/2022/jul/21/why-are-there-so-many-omicron-subvariants-and-why-have-ba4-and-ba5-taken-hold (accessed on 13 August 2023).
- Ren, S.-Y.; Wang, W.-B.; Gao, R.-D.; Zhou, A.-M. Omicron variant (B. 1.1. 529) of SARS-CoV-2: Mutation, infectivity, transmission, and vaccine resistance. World J. Clin. Cases. 2022, 10, 1. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, J.; Wang, J. Surface charge changes in spike RBD mutations of SARS-CoV-2 and its variant strains alter the virus evasiveness via HSPGs: A review and mechanistic hypothesis. Front. Public Health 2022, 10, 952916. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.; do Nascimento, G.M.; Nooruzzaman, M.; Yuan, F.; Chen, C.; Caserta, L.C.; Miller, A.D.; Whittaker, G.R.; Fang, Y.; Diel, D.G. The Omicron variant BA. 1.1 presents lower pathogenicity than B. 1 D614G and Delta variants in a feline model of SARS-CoV-2 infection. J. Virol. 2022, 96, e00961-22. [Google Scholar] [CrossRef] [PubMed]
- Arbel, R.; Peretz, A.; Sergienko, R.; Friger, M.; Beckenstein, T.; Yaron, S.; Hammerman, A.; Bilenko, N.; Netzer, D. Effectiveness of the bivalent mRNA vaccine in preventing severe COVID-19 outcomes: An observational cohort study. Lancet Infect. Dis. 2023; ahead of print. [Google Scholar] [CrossRef]
- Offit, P.A. Bivalent COVID-19 Vaccines—A Cautionary Tale. N. Engl. J. Med. 2023, 388, 481–483. [Google Scholar] [CrossRef]
- Choe, Y.J.; Choi, E.H.; Choi, J.W.; Eun, B.W.; Eun, L.Y.; Kim, Y.-J.; Kim, Y.H.; Kim, Y.A.; Kim, Y.-K.; Kwak, J.H. Change in Severity and Clinical Manifestation of MIS-C Over SARS-CoV-2 Variant Outbreaks in Korea. J. Korean Med. Sci. 2023, 38, e225. [Google Scholar] [CrossRef]
- Lee, D.-W.; Kim, J.-M.; Park, A.K.; Kim, D.-W.; Kim, J.-Y.; Lim, N.; Lee, H.; Kim, I.-H.; Kim, J.-A.; Lee, C.Y. Genomic epidemiology of SARS-CoV-2 Omicron variants in the Republic of Korea. Sci. Rep. 2022, 12, 22414. [Google Scholar] [CrossRef] [PubMed]
- Biosensor, S. STANDARD Q COVID-19 IgM/IgG Plus. 2021. Available online: https://www.sdbiosensor.com/product/product_view?product_no=239 (accessed on 29 June 2023).
- Biosensor, S. STANDARDTM Q COVID-19 IgM/IgG Plus Test. 2020. Available online: https://bestbion.com/wp-content/uploads/2021/05/bestbiondx-GA-STANDARD-Q-COVID-19-IgMIgG-Plus.pdf (accessed on 29 June 2023).
- Tan, C.W.; Chia, W.N.; Qin, X.; Liu, P.; Chen, M.I.-C.; Tiu, C.; Hu, Z.; Chen, V.C.-W.; Young, B.E.; Sia, W.R. A SARS-CoV-2 surrogate virus neutralization test based on antibody-mediated blockage of ACE2–spike protein–protein interaction. Nat. Biotechnol. 2020, 38, 1073–1078. [Google Scholar] [CrossRef] [PubMed]
- Nandakumar, V.; Profaizer, T.; Lozier, B.K.; Elgort, M.G.; Larragoite, E.T.; Williams, E.S.; Solis-Leal, A.; Lopez, J.B.; Berges, B.K.; Planelles, V. Evaluation of a Surrogate Enzyme-Linked Immunosorbent Assay–Based Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) cPass Neutralization Antibody Detection Assay and Correlation With Immunoglobulin G Commercial Serology Assays. Arch. Pathol. Lab. Med. 2021, 145, 1212–1220. [Google Scholar] [CrossRef]
- Hamady, A.; Lee, J.; Loboda, Z.A. Waning antibody responses in COVID-19: What can we learn from the analysis of other coronaviruses? Infection 2022, 50, 11–25. [Google Scholar] [CrossRef]
- Choe, P.G.; Kang, C.K.; Suh, H.J.; Jung, J.; Song, K.-H.; Bang, J.H.; Kim, E.S.; Kim, H.B.; Park, S.W.; Kim, N.J. Waning antibody responses in asymptomatic and symptomatic SARS-CoV-2 infection. Emerg. Infect. Dis. 2021, 27, 327. [Google Scholar] [CrossRef]
- Wilhelm, A.; Widera, M.; Grikscheit, K.; Toptan, T.; Schenk, B.; Pallas, C.; Metzler, M.; Kohmer, N.; Hoehl, S.; Marschalek, R. Limited neutralisation of the SARS-CoV-2 Omicron subvariants BA. 1 and BA. 2 by convalescent and vaccine serum and monoclonal antibodies. EBioMedicine 2022, 82, 104158. [Google Scholar] [CrossRef]
- Ikemura, N.; Hoshino, A.; Higuchi, Y.; Taminishi, S.; Inaba, T.; Matoba, S. SARS-CoV-2 Omicron variant escapes neutralization by vaccinated and convalescent sera and therapeutic monoclonal antibodies. MedRxiv, 2021; Preprint. [Google Scholar] [CrossRef]
- Gruell, H.; Vanshylla, K.; Korenkov, M.; Tober-Lau, P.; Zehner, M.; Münn, F.; Janicki, H.; Augustin, M.; Schommers, P.; Sander, L.E. SARS-CoV-2 Omicron sublineages exhibit distinct antibody escape patterns. Cell Host Microbe 2022, 30, 1231–1241. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Wang, J.; Jian, F.; Xiao, T.; Song, W.; Yisimayi, A.; Huang, W.; Li, Q.; Wang, P.; An, R. Omicron escapes the majority of existing SARS-CoV-2 neutralizing antibodies. Nature 2022, 602, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Nie, J.; Wu, J.; Zhang, L.; Ding, R.; Wang, H.; Zhang, Y.; Li, T.; Liu, S.; Zhang, M. SARS-CoV-2 501Y. V2 variants lack higher infectivity but do have an immune escape. Cell 2021, 184, 2362–2371. [Google Scholar] [CrossRef]
- Starr, T.N.; Greaney, A.J.; Hilton, S.K.; Ellis, D.; Crawford, K.H.; Dingens, A.S.; Navarro, M.J.; Bowen, J.E.; Tortorici, M.A.; Walls, A.C. Deep mutational scanning of SARS-CoV-2 receptor binding domain reveals constraints on folding and ACE2 binding. Cell 2020, 182, 1295–1310. [Google Scholar] [CrossRef] [PubMed]
- FDA. Coronavirus (COVID-19) Update: FDA Authorizes Moderna, Pfizer-BioNTech Bivalent COVID-19 Vaccines for Use as a Booster Dose. 2022. Available online: https://www.fda.gov/news-events/press-announcements/coronavirus-covid-19-update-fda-authorizes-moderna-pfizer-biontech-bivalent-covid-19-vaccines-use (accessed on 4 July 2023).
- Han-soo, L. Korea to Start Administering Updated COVID-19 Vaccine from 11 October 2022. Available online: https://www.koreabiomed.com/news/articleView.html?idxno=14644 (accessed on 5 July 2023).
- Korean Ministry of Health and Welfare. Basic Directions for the COVID-19 Vaccination Scheme in the 2022-23 Winter. 2022. Available online: https://www.mohw.go.kr/eng/nw/nw0101vw.jsp?PAR_MENU_ID=1007&MENU_ID=100701&page=1&CONT_SEQ=372816 (accessed on 5 July 2023).
- Yoon-seung, K. (LEAD) S. Korea’s New COVID-19 Cases Rise More Than 11,000 On-Week. 2022. Available online: https://en.yna.co.kr/view/AEN20221113000651320 (accessed on 5 July 2023).
- Collier, A.-R.Y.; Brown, C.M.; McMahan, K.A.; Yu, J.; Liu, J.; Jacob-Dolan, C.; Chandrashekar, A.; Tierney, D.; Ansel, J.L.; Rowe, M. Characterization of immune responses in fully vaccinated individuals after breakthrough infection with the SARS-CoV-2 delta variant. Sci. Transl. Med. 2022, 14, eabn6150. [Google Scholar] [CrossRef]
- The Lancet Infectious Disease Editorial. Why hybrid immunity is so triggering. Lancet Infect. Dis. 2022, 22, 1649. [Google Scholar] [CrossRef]
- Han-soo, L. Korea to No Longer Use Moderna’s Original Monovalent COVID-19 Vaccine. 2022. Available online: https://www.koreabiomed.com/news/articleView.html?idxno=15084 (accessed on 12 July 2023).



{kind=link}
{kind=link}
| Characteristics | Uninfected Population N = 38 | Infected Population N = 31 |
|---|---|---|
| Sex | ||
| Male, N (%) | 12 (31.6) | 9 (29.1) |
| Female, N (%) | 26 (68.4) | 22 (70.9) |
| Age (years), median (range) | 43 (28–63) | 42 (25–58) |
| Vaccine type | ||
| BA.1 MD, N (%) | 3 (7.8%) | 2 (6.4%) |
| BA.1 PF, N (%) | 1 (2.6%) | 0 (0%) |
| BA.4/5 PF, N (%) | 33 (86.8%) | 29 (93.6%) |
| N/D, N (%) | 1 (2.6%) | 0 (0%) |
| STANDARD Q COVID-19 IgM/IgG Plus | Uninfected (N = 38), % | Infected (N = 31), % |
|---|---|---|
| Negative | 1 (2.6%) | 0 (0%) |
| Positive | 37 (97.4%) | 31 (100%) |
| cPass sVNT | Uninfected (N = 38), % | Infected (N = 31), % |
|---|---|---|
| Negative | 0 (0%) | 0 (0%) |
| Positive | 38 (100%) | 31 (100%) |
| Vaccination Status | Uninfected (N = 38) | Infected (N = 31) | ||
|---|---|---|---|---|
| N | Median (IQR) | N | Median (IQR) | |
| Before vaccination | 38 | 42.8% (25.1–63.4%) | 31 | 65.0% (46.1–77.2%) |
| After vaccination | 38 | 91.9% (71.6–96.4%) | 31 | 90.5% (67.2–98.1%) |
| BA.1 MD | 3 | 90.1% (86.5–99.6%) | 2 | 96.5% (94.8–98.1%) |
| BA.1 PF | 1 | 96.0% (96.0–96.0%) | 0 | N/A |
| BA.4/5 PF | 33 | 91.8% (66.7–96.0%) | 29 | 84.9% (67.2–96.8%) |
| N/D | 1 | 98.3% (98.3–98.3%) | 0 | N/A |
| Vaccination Status | Uninfected (N = 38) | Infected (N = 31) | ||
|---|---|---|---|---|
| N | Median (IQR) | N | Median (IQR) | |
| Before vaccination | 38 | 42.3 (26.2–199.3) | 31 | 209.5 (91.5–369.3) |
| After vaccination | 38 | 378.5 (180.7–828.1) | 31 | 823.6 (370.0–1173.2) |
| BA.1 MD | 3 | 215.5 (46.4–1617.6) | 2 | 1088.1 (1083.1–1093.1) |
| BA.1 PF | 1 | 565.7 (565.7–565.7) | 0 | N/A |
| BA.4/5 PF | 33 | 377.7 (180.7–803.5) | 29 | 800.8 (370.0–1173.2) |
| N/D | 1 | 828.1 (828.1–828.1) | 0 | N/A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Widyasari, K.; Jang, J.; Kang, T.; Kim, S. Effectiveness of Bivalent Omicron-Containing Booster Vaccines against SARS-CoV-2 Omicron Variant among Individuals with and without Prior SARS-CoV-2 Infection. Viruses 2023, 15, 1756. https://doi.org/10.3390/v15081756
Widyasari K, Jang J, Kang T, Kim S. Effectiveness of Bivalent Omicron-Containing Booster Vaccines against SARS-CoV-2 Omicron Variant among Individuals with and without Prior SARS-CoV-2 Infection. Viruses. 2023; 15(8):1756. https://doi.org/10.3390/v15081756
Chicago/Turabian StyleWidyasari, Kristin, Jieun Jang, Taejoon Kang, and Sunjoo Kim. 2023. "Effectiveness of Bivalent Omicron-Containing Booster Vaccines against SARS-CoV-2 Omicron Variant among Individuals with and without Prior SARS-CoV-2 Infection" Viruses 15, no. 8: 1756. https://doi.org/10.3390/v15081756