
Efficacy, Pharmacokinetics, and Toxicity Profiles of a Broad Anti-SARS-CoV-2 Neutralizing Antibody
 ,
,  , and
, and 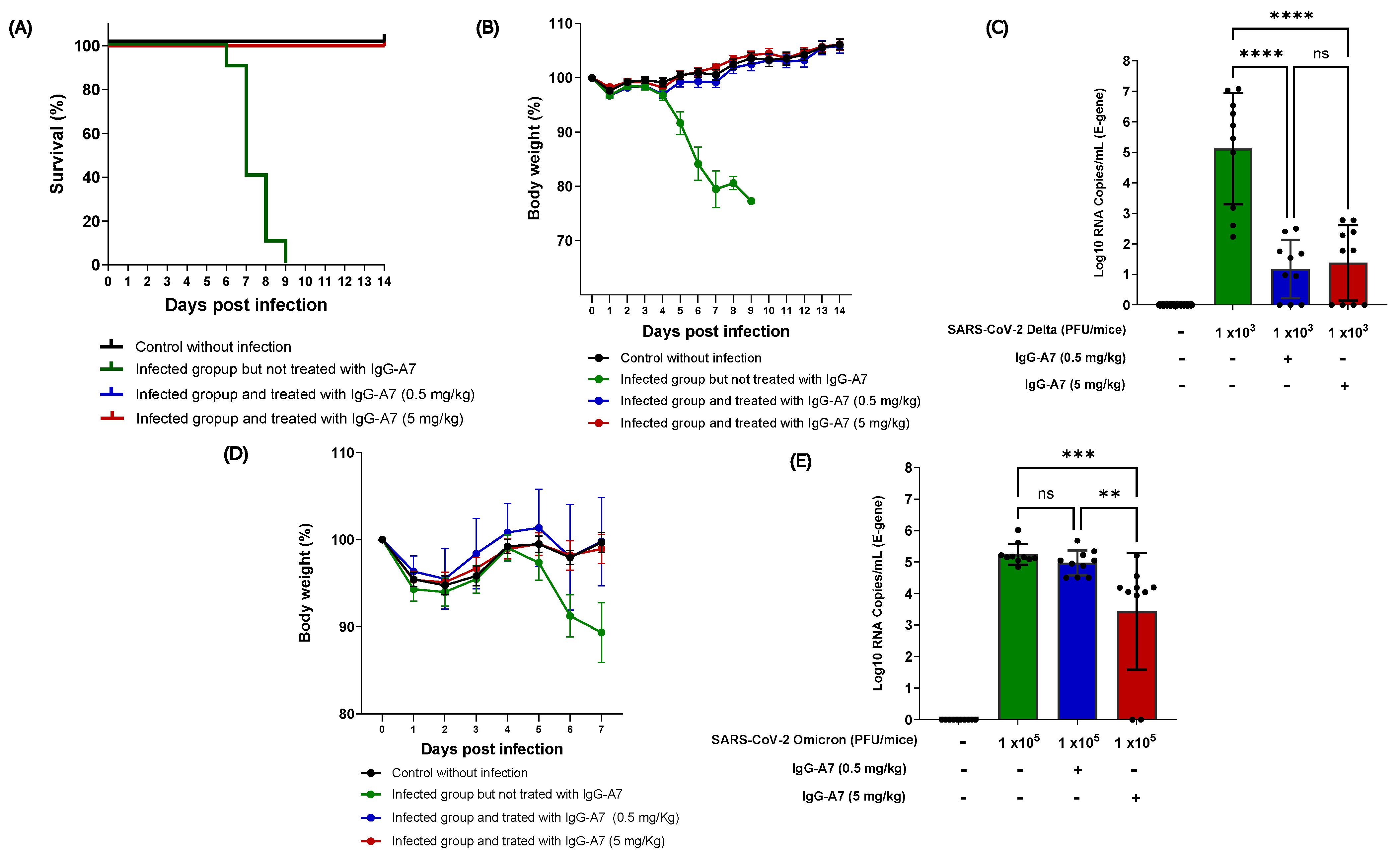{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Ethics
2.2. Efficacy of IgG-A7 against SARS-CoV-2 Delta and Omicron
2.3. PK Study at High Single Doses
2.4. Tox Studies
2.5. TCR in Mouse and Humans
3. Results
3.1. IgG-A7 Efficacy against Delta and Omicron SARS-CoV-2 Variants
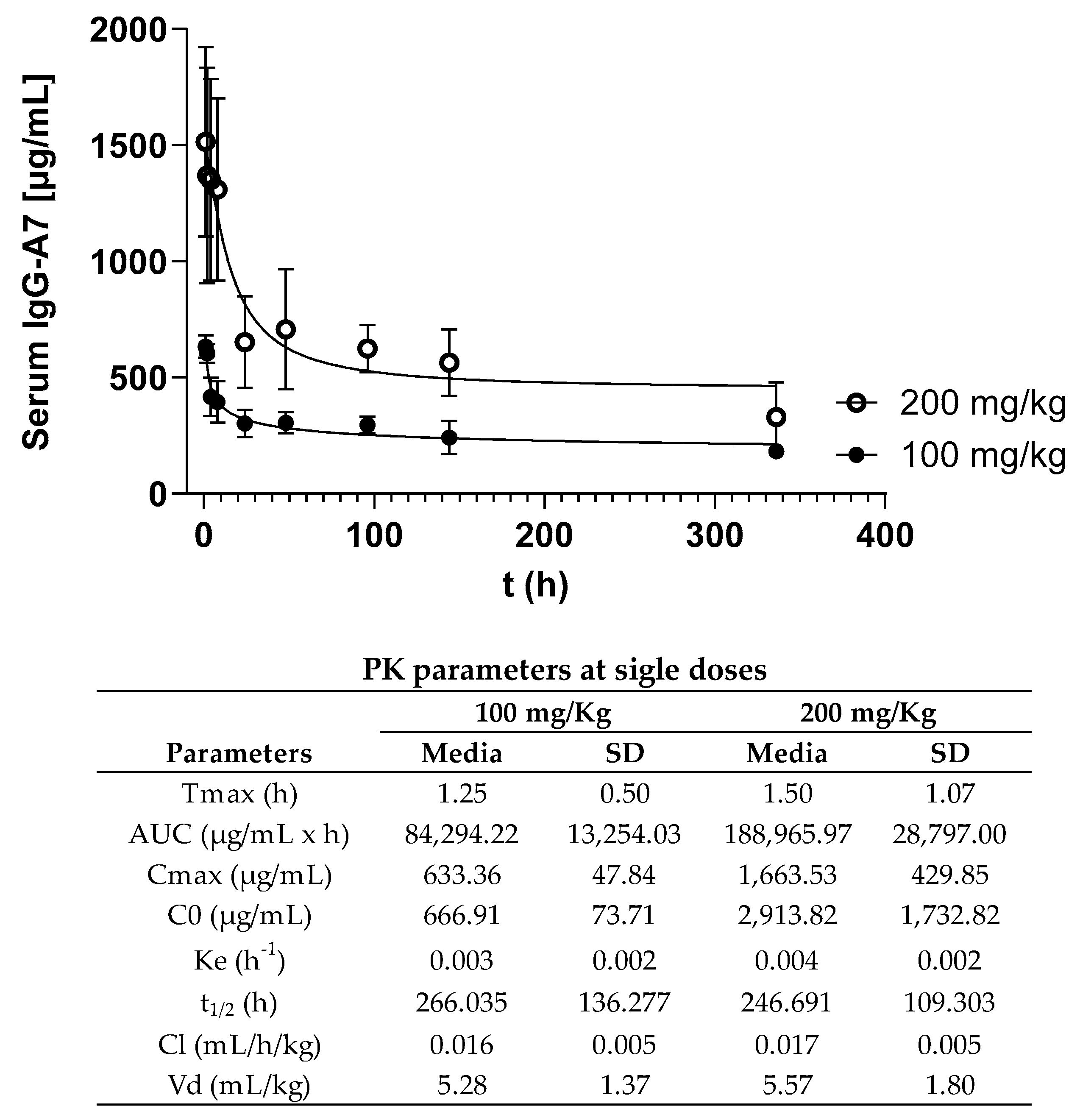
3.2. PK Profiling
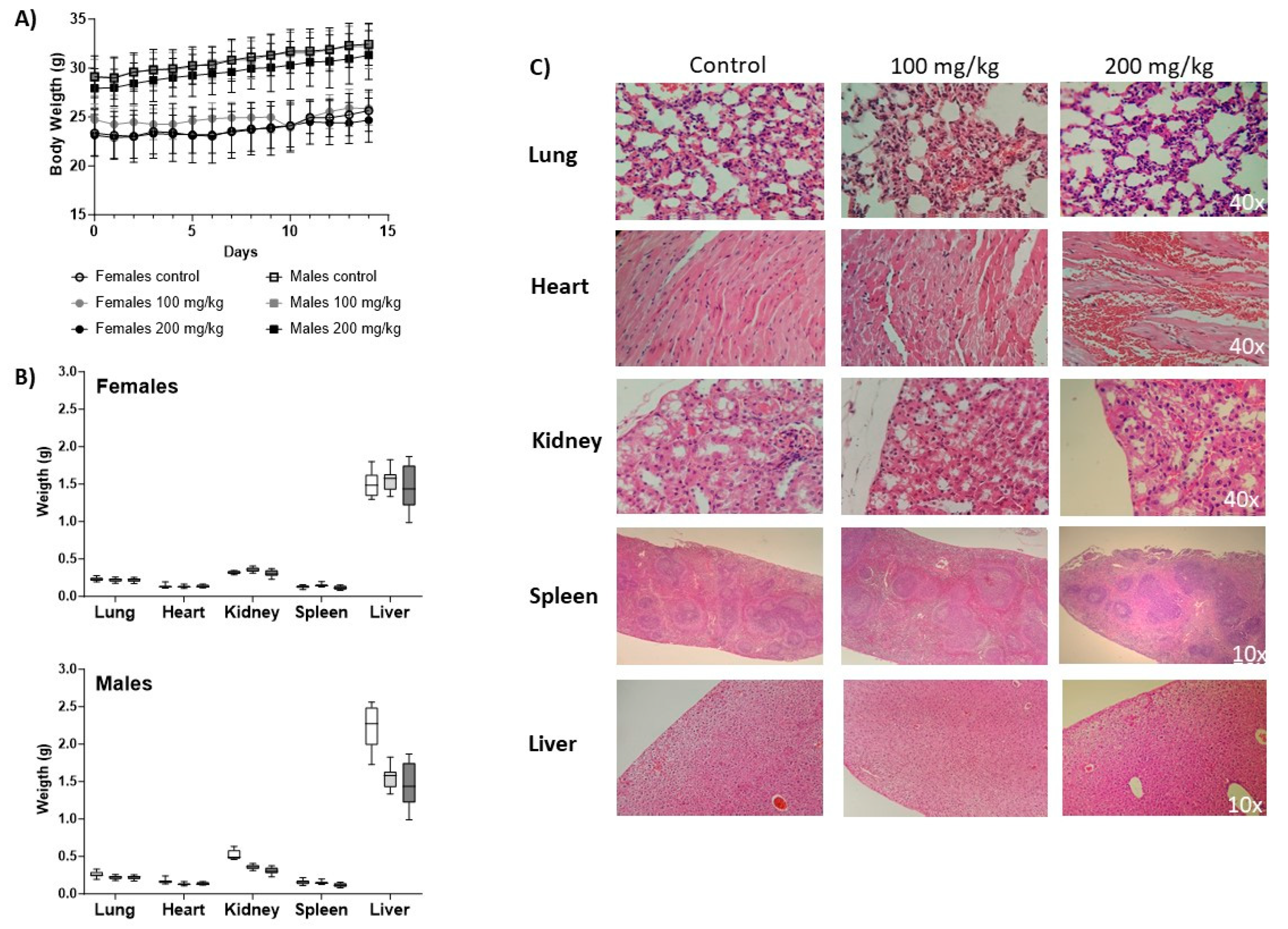
3.3. Tox Assessment
3.3.1. Body Weight
3.3.2. Urinalysis
3.3.3. Hematology
3.3.4. Blood Chemistry
3.3.5. Histopathology
3.4. Mouse and Human TCR
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Almagro, J.C.; Mellado-Sanchez, G.; Pedraza-Escalona, M.; Perez-Tapia, S.M. Evolution of Anti-SARS-CoV-2 Therapeutic Antibodies. Int. J. Mol. Sci. 2022, 23, 9763. [Google Scholar] [CrossRef] [PubMed]
- Takashita, E.; Kinoshita, N.; Yamayoshi, S.; Sakai-Tagawa, Y.; Fujisaki, S.; Ito, M.; Iwatsuki-Horimoto, K.; Halfmann, P.; Watanabe, S.; Maeda, K.; et al. Efficacy of Antiviral Agents against the SARS-CoV-2 Omicron Subvariant BA.2. N. Engl. J. Med. 2022, 386, 1475–1477. [Google Scholar] [CrossRef]
- VanBlargan, L.A.; Errico, J.M.; Halfmann, P.J.; Zost, S.J.; Crowe, J.E.; Purcell, L.A.; Kawaoka, Y.; Corti, D.; Fremont, D.H.; Diamond, M.S. An infectious SARS-CoV-2 B.1.1.529 Omicron virus escapes neutralization by therapeutic monoclonal antibodies. Nat. Med. 2022, 28, 490–495. [Google Scholar] [CrossRef]
- Singh, D.D.; Sharma, A.; Lee, H.J.; Yadav, D.K. SARS-CoV-2: Recent Variants and Clinical Efficacy of Antibody-Based Therapy. Front. Cell Infect. Microbiol. 2022, 12, 839170. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-Salazar, I.; Gómez-Castellano, K.M.; González-González, E.; Gamboa-Suasnavart, R.; Rodríguez-Luna, S.D.; Santiago-Casas, G.; Cortés-Paniagua, M.I.; Pérez-Tapia, S.M.; Almagro, J.C. Anti-SARS-CoV-2 Omicron Antibodies Isolated from a SARS-CoV-2 Delta Semi-Immune Phage Display Library. Antibodies 2022, 11, 13. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Gonzalez, E.; Carballo-Uicab, G.; Salinas-Trujano, J.; Cortes-Paniagua, M.I.; Vazquez-Leyva, S.; Vallejo-Castillo, L.; Mendoza-Salazar, I.; Gomez-Castellano, K.; Perez-Tapia, S.M.; Almagro, J.C. In Vitro and In Vivo Characterization of a Broadly Neutralizing Anti-SARS-CoV-2 Antibody Isolated from a Semi-Immune Phage Display Library. Antibodies 2022, 11, 57. [Google Scholar] [CrossRef] [PubMed]
- NOM-062-ZOO-1999; Norma Oficial Mexicana De La Federación-NOM-062-ZOO-1999, Especificaciones Técnicas Para la Producción, Cuidado y Uso de los Animales de Laboratorio. Comité Consultivo Nacional de Normalización de Protección Zoosanitaria: Mexico, 2001; p. 477.
- Bewley, K.R.; Coombes, N.S.; Gagnon, L.; McInroy, L.; Baker, N.; Shaik, I.; St-Jean, J.R.; St-Amant, N.; Buttigieg, K.R.; Humphries, H.E.; et al. Quantification of SARS-CoV-2 neutralizing antibody by wild-type plaque reduction neutralization, microneutralization and pseudotyped virus neutralization assays. Nat. Protoc. 2021, 16, 3114–3140. [Google Scholar] [CrossRef]
- Biosafety in Microbiological and Biomedical Laboratories—6th Edition: 604. Available online: https://www.cdc.gov/labs/pdf/SF__19_308133-A_BMBL6_00-BOOK-WEB-final-3.pdf (accessed on 11 June 2022).
- Laboratory Biosafety Manual, 4th Edition. Available online: https://www.who.int/publications-detail-redirect/9789240011311 (accessed on 11 June 2022).
- CDC. Labs. Centers for Disease Control and Prevention. Published 11 February 2020. Available online: https://www.cdc.gov/coronavirus/2019-ncov/lab/lab-safety-practices.html (accessed on 21 June 2022).
- Guidance-for-BSL2+ COVID-Laboratories-Anschutz.pdf. Available online: https://research.cuanschutz.edu/docs/librariesprovider148/ehs_documents/default-library/guidance-for-bsl2-covid-laboratories-anschutz.pdf?sfvrsn=79c7e4b9_0 (accessed on 21 June 2022).
- Camacho-Sandoval, R.; Nieto-Patlán, A.; Carballo-Uicab, G.; Montes-Luna, A.; Jiménez-Martínez, M.C.; Vallejo-Castillo, L.; González-González, E.; Arrieta-Oliva, H.I.; Gómez-Castellano, K.; Guzmán-Bringas, O.U.; et al. Development and Evaluation of a Set of Spike and Receptor Binding Domain-Based Enzyme-Linked Immunosorbent Assays for SARS-CoV-2 Serological Testing. Diagnostics 2021, 11, 1506. [Google Scholar] [CrossRef]
- ICH Harmonised Guideline. M10 Bioanalytical Method Validation and Study Sample Analysis to be implemented by PQT/MED. In WHO-Prequalification of Medical Products (IVDs, Medicines, Vaccines and Immunization Devices, Vector Control); WHO: Geneva, Switzerland, 2023. [Google Scholar]
- Halfmann, P.J.; Iida, S.; Iwatsuki-Horimoto, K.; Maemura, T.; Kiso, M.; Scheaffer, S.M.; Darling, T.L.; Joshi, A.; Loeber, S.; Singh, G.; et al. SARS-CoV-2 Omicron virus causes attenuated disease in mice and hamsters. Nature 2022, 603, 687–692. [Google Scholar] [CrossRef]
- Leach, M.W.; Halpern, W.G.; Johnson, C.W.; Rojko, J.L.; MacLachlan, T.K.; Chan, C.M.; Galbreath, E.J.; Ndifor, A.M.; Blanset, D.L.; Polack, E.; et al. Use of tissue cross-reactivity studies in the development of antibody-based biopharmaceuticals: History, experience, methodology, and future directions. Toxicol. Pathol. 2010, 38, 1138–1166. [Google Scholar] [CrossRef]
- Luo, F.R.; Yang, Z.; Dong, H.; Camuso, A.; McGlinchey, K.; Fager, K.; Flefleh, C.; Kan, D.; Inigo, I.; Castaneda, S.; et al. Correlation of pharmacokinetics with the antitumor activity of Cetuximab in nude mice bearing the GEO human colon carcinoma xenograft. Cancer Chemother. Pharmacol. 2005, 56, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kim, D.; Son, E.; Yoo, S.J.; Sa, J.K.; Shin, Y.J.; Yoon, Y.; Nam, D.H. Pharmacokinetics, Biodistribution, and Toxicity Evaluation of Anti-SEMA3A (F11) in In Vivo Models. Anticancer. Res. 2018, 38, 2803–2810. [Google Scholar] [CrossRef]
- Sandker, G.G.W.; Adema, G.; Molkenboer-Kuenen, J.; Wierstra, P.; Bussink, J.; Heskamp, S.; Aarntzen, E. PD-L1 Antibody Pharmacokinetics and Tumor Targeting in Mouse Models for Infectious Diseases. Front. Immunol. 2022, 13, 837370. [Google Scholar] [CrossRef]
- Deng, R.; Bumbaca, D.; Pastuskovas, C.V.; Boswell, C.A.; West, D.; Cowan, K.J.; Chiu, H.; Mcbride, J.; Johnson, C.; Xin, Y.; et al. Preclinical pharmacokinetics, pharmacodynamics, tissue distribution, and tumor penetration of anti-PD-L1 monoclonal antibody, an immune checkpoint inhibitor. mAbs 2016, 8, 593–603. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, D.P.; Cooke, R.J.; McLachlan, A.J.; Daley-Yates, P.T.; Rowland, M. Pharmacokinetics and tissue distribution of cisplatin and conjugates of cisplatin with carboxymethyldextran and A5B7 monoclonal antibody in CD1 mice. J. Pharm. Sci. 1997, 86, 1478–1483. [Google Scholar] [CrossRef]
- Xie, H.; Audette, C.; Hoffee, M.; Lambert, J.M.; Blättler, W.A. Pharmacokinetics and biodistribution of the antitumor immunoconjugate, cantuzumab mertansine (huC242-DM1), and its two components in mice. J. Pharmacol. Exp. Ther. 2004, 308, 1073–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jha, A.; Doyle-Eisele, M.; Revelli, D.; Carnelley, T.; Barker, D.; Kodihalli, S. Pharmacokinetic and Pharmacodynamic Effects of Polyclonal Antibodies against SARS-CoV2 in Mice. Viruses 2022, 15, 123. [Google Scholar] [CrossRef]
- Unverdorben, F.; Richter, F.; Hutt, M.; Seifert, O.; Malinge, P.; Fischer, N.; Kontermann, R.E. Pharmacokinetic properties of IgG and various Fc fusion proteins in mice. mAbs 2016, 8, 120–128. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Ulrich, M.L.; Shih, V.F.; Cochran, J.H.; Hunter, J.H.; Westendorf, L.; Neale, J.; Benjamin, D.R. Mouse Strains Influence Clearance and Efficacy of Antibody and Antibody-Drug Conjugate Via Fc-FcγR Interaction. Mol. Cancer Ther. 2019, 18, 780–787. [Google Scholar] [CrossRef] [Green Version]
- Kurtz, D.M.; Travlos, G.S. (Eds.) The Clinical Chemistry of Laboratory Animals, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2017; p. 1162. [Google Scholar]
- CD-1® IGS Mouse. Available online: https://www.criver.com/products-services/find-model/cd-1r-igs-mouse?region=3616 (accessed on 26 June 2023).
- DiBartola, S.P.; Willard, M.D. Chapter 7-Disorders of Phosphorus: Hypophosphatemia and Hyperphosphatemia. In Fluid, Electrolyte, and Acid-Base Disorders in Small Animal Practice, 3rd ed.; Dibartola, S.P., Ed.; W.B. Saunders: Saint Louis, MO, USA, 2006; pp. 195–209. [Google Scholar]
- Moe, S.M. Disorders Involving Calcium, Phosphorus, and Magnesium. Prim. Care Clin. Off. Pract. 2008, 35, 215–237. [Google Scholar] [CrossRef] [Green Version]
- Center for Drug Evaluation and Research. Development of Monoclonal Antibody Products Targeting SARS-CoV-2, Including Addressing the Impact of Emerging Variants, during the COVID 19 Public Health Emergency; U.S. Food and Drug Administration: Rockville, MD, USA, 2023. [Google Scholar]
- European Medicines Agency. Development, Production, Characterisation and Specifications for Monoclonal Antibodies Related Products-Scientific Guideline; European Medicines Agency: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Center for Biologics Evaluation and Research. Points to Consider in the Manufacture and Testing of Monoclonal Antibody Products for Human Use; U.S. Food and Drug Administration: Rockville, MD, USA, 2020. [Google Scholar]
- Carvalho, C.; Gaspar, A.; Knight, A.; Vicente, L. Ethical and Scientific Pitfalls Concerning Laboratory Research with Non-Human Primates, and Possible Solutions. Animals 2018, 9, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Godínez-Palma, S.; González-González, E.; Ramírez-Villedas, F.; Garzón-Guzmán, C.; Vallejo-Castillo, L.; Carballo-Uicab, G.; Marcelín-Jiménez, G.; Batista, D.; Pérez-Tapia, S.M.; Almagro, J.C. Efficacy, Pharmacokinetics, and Toxicity Profiles of a Broad Anti-SARS-CoV-2 Neutralizing Antibody. Viruses 2023, 15, 1733. https://doi.org/10.3390/v15081733
Godínez-Palma S, González-González E, Ramírez-Villedas F, Garzón-Guzmán C, Vallejo-Castillo L, Carballo-Uicab G, Marcelín-Jiménez G, Batista D, Pérez-Tapia SM, Almagro JC. Efficacy, Pharmacokinetics, and Toxicity Profiles of a Broad Anti-SARS-CoV-2 Neutralizing Antibody. Viruses. 2023; 15(8):1733. https://doi.org/10.3390/v15081733
Chicago/Turabian StyleGodínez-Palma, Silvia, Edith González-González, Frida Ramírez-Villedas, Circe Garzón-Guzmán, Luis Vallejo-Castillo, Gregorio Carballo-Uicab, Gabriel Marcelín-Jiménez, Dany Batista, Sonia M. Pérez-Tapia, and Juan C. Almagro. 2023. "Efficacy, Pharmacokinetics, and Toxicity Profiles of a Broad Anti-SARS-CoV-2 Neutralizing Antibody" Viruses 15, no. 8: 1733. https://doi.org/10.3390/v15081733