
Rotavirus Particle Disassembly and Assembly In Vivo and In Vitro

{kind=link}
{kind=link}
{kind=link}
{kind=link}
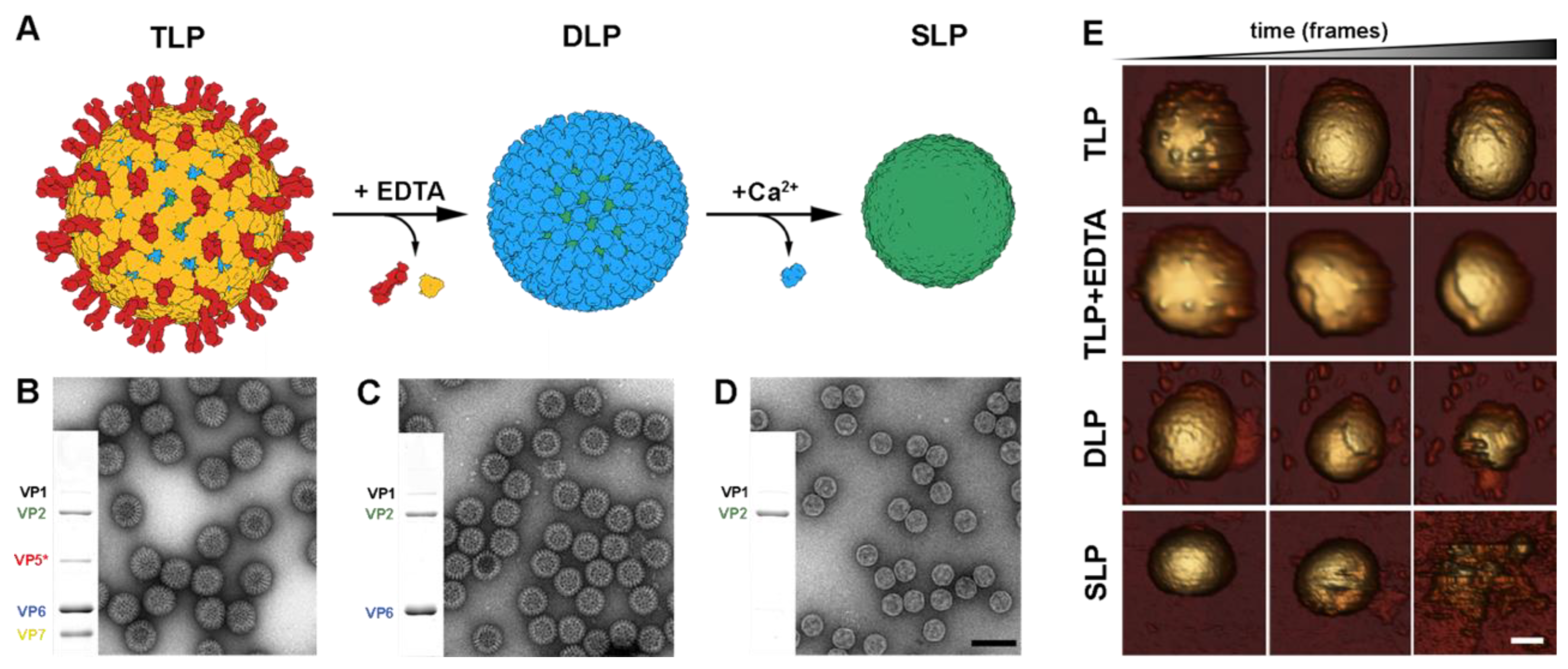{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Rotavirus Virion Structure

3. Particle Disassembly during the Viral Cycle

4. Particle Assembly during the Viral Cycle
5. In Vitro Disassembly and Assembly of RV Particles
6. In Vivo Assembly of RV Virus-like Particles

7. Future Outlook
- The current molecular model for membrane disruption and entry is being developed in an outlier system (RRV strain/BSC-1 cells) that does not depend on the cellular endocytic machinery and instead relies solely on the viral machinery for entry from the plasma membrane. How does this model apply to other rotavirus strains that do rely on the cellular endocytic machinery and enter the cell from late or recycling endosomes?
- What are the mechanisms underlying the entry and non-lytic release of rotavirus as vesicle-enveloped clusters, and how does vesicle-mediated en bloc transmission affect viral pathogenesis, spread, and evolution?
- Identifying the membrane in eDLP as of COPII origin is a paradigm-shifting result that awaits further characterization of the mechanisms involved. Additionally, it is crucial to investigate the extent to which rotavirus disrupts the autophagy system and the specific mechanisms underlying VP7 transport within these membranes.
- The description of viroplasms as biomolecular condensates is crucial in understanding the dynamics of these viral organelles. To develop a comprehensive understanding of the formation and maturation of viroplasms, it is important to address key questions related to the maturation of viroplasms, the roles of lipid droplets and other cellular components in the formation of viroplasms, and the roles of NSP2 and VP2 in the formation and evolution of initial genome complexes.
- The description of viroplasms as highly organized structures with a concentric distribution of viral proteins around a center formed by NSP5 suggests that there exists a temporal and spatial coordination of viral processes, which has yet to be characterized in detail.
- Cryo-electron tomography of cellular lamellae and subtomogram averaging have provided valuable insights into the structure of eDLP and have been demonstrated as a promising approach to characterizing transient rotavirus structures in their native cellular environment. With further advancements in these techniques, it is anticipated that the reconstruction of VP7 and NSP4 within the eDLP can be achieved.
- The presence and the structure of the genome-less SLP are difficult to conciliate with current models for rotavirus particle assembly and require further investigation.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mertens, P. The DsRNA Viruses. Virus Res. 2004, 101, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Frias, A.H.; Jones, R.M.; Fifadara, N.H.; Vijay-Kumar, M.; Gewirtz, A.T. Rotavirus-Induced IFN-β Promotes Anti-Viral Signaling and Apoptosis That Modulate Viral Replication in Intestinal Epithelial Cells. Innate Immun. 2012, 18, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Arnold, M.M.; Sen, A.; Greenberg, H.B.; Patton, J.T. The Battle between Rotavirus and Its Host for Control of the Interferon Signaling Pathway. PLoS Pathog. 2013, 9, e1003064. [Google Scholar] [CrossRef] [PubMed]
- Patton, J.T. Segmented Double-Stranded RNA Viruses. Structure and Molecular Biology; Caister Academic Press: Norfolk, VA, USA, 2008. [Google Scholar]
- Harrison, S.C. Principles of Virus Structure. In Fields Virology; Knipe, D.M., Howley, P.M., Griffin, D.E., Martin, M.A., Lamb, R.A., Roizman, B., Strauss, S.E., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; Volume 1. [Google Scholar]
- King, A.; Lefkowitz, E.J.; Adams, M.J.; Carstens, E.B. Virus Taxonomy. Ninth Report of the International Committee on Taxonomy of Viruses; Academic Press: San Diego, CA, USA, 2011. [Google Scholar]
- Mata, C.P.; Rodríguez, J.M.; Suzuki, N.; Castón, J.R. Structure and Assembly of Double-Stranded RNA Mycoviruses. Adv. Virus Res. 2020, 108, 213–247. [Google Scholar]
- Crawford, S.E.; Ramani, S.; Tate, J.E.; Parashar, U.D.; Svensson, L.; Hagbom, M.; Franco, M.A.; Greenberg, H.B.; O’Ryan, M.; Kang, G.; et al. Rotavirus Infection. Nat. Rev. Dis. Primers 2017, 3, 17083. [Google Scholar] [CrossRef]
- Rodríguez, J.M.; Luque, D. Structural Insights into Rotavirus Entry. In Advances in Experimental Medicine and Biology; Greber, U.F., Ed.; Springer: Cham, Switzerland, 2018; pp. 45–68. [Google Scholar]
- Dermody, T.S.; Parker, J.S.; Sherry, B. Orthoreoviruses. In Fields Virology; Knipe, D.M., Howley, P.M., Cohen, J.I., Griffin, D.E., Lamb, R.A., Martin, M.A., Roizman, B., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; Volume 1. [Google Scholar]
- Roy, P. Orbiviruses. In Fields Virology; Knipe, D.M., Howley, P.M., Cohen, J.I., Griffin, D.E., Lamb, R.A., Martin, M.A., Roizman, B., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; Volume 1. [Google Scholar]
- Estes, M.K.; Greenberg, H.B. Rotaviruses. In Fields Virology; Knipe, D.M., Howley, P.M., Cohen, J.I., Griffin, D.E., Lamb, R.A., Martin, M.A., Roizman, B., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; Volume 1. [Google Scholar]
- Du, Y.; Chen, C.; Zhang, X.; Yan, D.; Jiang, D.; Liu, X.; Yang, M.; Ding, C.; Lan, L.; Hecht, R.; et al. Global Burden and Trends of Rotavirus Infection-Associated Deaths from 1990 to 2019: An Observational Trend Study. Virol. J. 2022, 19, 166. [Google Scholar] [CrossRef]
- Bishop, R.F.; Davidson, G.P.; Holmes, I.H.; Ruck, B.J. Virus Particles in Epithelial Cells of Duodenal Mucosa from Children with Acute Non-Bacterial Gastroenteritis. Lancet 1973, 2, 1281–1283. [Google Scholar] [CrossRef]
- Troeger, C.; Khalil, I.A.; Rao, P.C.; Cao, S.; Blacker, B.F.; Ahmed, T.; Armah, G.; Bines, J.E.; Brewer, T.G.; Colombara, D.V.; et al. Rotavirus Vaccination and the Global Burden of Rotavirus Diarrhea among Children Younger Than 5 Years. JAMA Pediatr. 2018, 172, 958–965. [Google Scholar] [CrossRef]
- Troeger, C.; Blacker, B.F.; Rao, P.C.; Khalil, I.; Brown, A.; Reiner, R.C., Jr.; Fullman, N.; Thompson, R.L.; Abajobir, A.; Ahmed, M.; et al. Estimates of Global, Regional, and National Morbidity, Mortality, and Aetiologies of Diarrhoeal Diseases: A Systematic Analysis for the Global Burden of Disease Study 2015. Lancet Infect. Dis. 2017, 17, 909–948. [Google Scholar] [CrossRef]
- Pesavento, J.B.; Crawford, S.E.; Estes, M.K.; Prasad, B. V Rotavirus Proteins: Structure and Assembly. Curr. Top. Microbiol. Immunol. 2006, 309, 189–219. [Google Scholar]
- Flewett, T.H.; Bryden, A.S.; Davies, H.; Woode, G.N.; Bridger, J.C.; Derrick, J.M. Relation between Viruses from Acute Gastroenteritis of Children and Newborn Calves. Lancet 1974, 2, 61–63. [Google Scholar] [CrossRef]
- Prasad, B.V.; Wang, G.J.; Clerx, J.P.; Chiu, W. Three-Dimensional Structure of Rotavirus. J. Mol. Biol. 1988, 199, 269–275. [Google Scholar] [CrossRef]
- Settembre, E.C.; Chen, J.Z.; Dormitzer, P.R.; Grigorieff, N.; Harrison, S.C. Atomic Model of an Infectious Rotavirus Particle. EMBO J. 2011, 30, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Jenni, S.; Salgado, E.N.; Herrmann, T.; Li, Z.; Grant, T.; Grigorieff, N.; Trapani, S.; Estrozi, L.F.; Harrison, S.C. In Situ Structure of Rotavirus VP1 RNA-Dependent RNA Polymerase. J. Mol. Biol. 2019, 431, 3124–3138. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Settembre, E.; Xu, C.; Dormitzer, P.R.; Bellamy, R.; Harrison, S.C.; Grigorieff, N. Near-Atomic Resolution Using Electron Cryomicroscopy and Single-Particle Reconstruction. Proc. Natl. Acad. Sci. USA 2008, 105, 1867–1872. [Google Scholar] [CrossRef]
- McClain, B.; Settembre, E.; Temple, B.R.; Bellamy, A.R.; Harrison, S.C. X-ray Crystal Structure of the Rotavirus Inner Capsid Particle at 3.8 A Resolution. J. Mol. Biol. 2010, 397, 587–599. [Google Scholar] [CrossRef] [PubMed]
- Ding, K.; Celma, C.C.; Zhang, X.; Chang, T.; Shen, W.; Atanasov, I.; Roy, P.; Zhou, Z.H. In Situ Structures of Rotavirus Polymerase in Action and Mechanism of MRNA Transcription and Release. Nat. Commun. 2019, 10, 2216. [Google Scholar] [CrossRef]
- Periz, J.; Celma, C.; Jing, B.; Pinkney, J.N.M.M.; Roy, P.; Kapanidis, A.N. Rotavirus mRNAS Are Released by Transcript-Specific Channels in the Double-Layered Viral Capsid. Proc. Natl. Acad. Sci. USA 2013, 110, 12042–12047. [Google Scholar] [CrossRef]
- Mathieu, M.; Petitpas, I.; Navaza, J.; Lepault, J.; Kohli, E.; Pothier, P.; Prasad, B.V.; Cohen, J.; Rey, F.A. Atomic Structure of the Major Capsid Protein of Rotavirus: Implications for the Architecture of the Virion. EMBO J. 2001, 20, 1485–1497. [Google Scholar] [CrossRef]
- Chen, J.Z.; Settembre, E.C.; Aoki, S.T.; Zhang, X.; Bellamy, A.R.; Dormitzer, P.R.; Harrison, S.C.; Grigorieff, N. Molecular Interactions in Rotavirus Assembly and Uncoating Seen by High-Resolution Cryo-EM. Proc. Natl. Acad. Sci. USA 2009, 106, 10644–10648. [Google Scholar] [CrossRef]
- Dormitzer, P.R.; Greenberg, H.B.; Harrison, S.C. Purified Recombinant Rotavirus VP7 Forms Soluble, Calcium-Dependent Trimers. Virology 2000, 277, 420–428. [Google Scholar] [CrossRef]
- Jiménez-Zaragoza, M.; Yubero, M.; Martín-Forero, E.; Castón, J.R.; Reguera, D.; Luque, D.; de Pablo, P.J.; Rodríguez, J.M. Biophysical Properties of Single Rotavirus Particles Account for the Functions of Protein Shells in a Multilayered Virus. Elife 2018, 7, e37295. [Google Scholar] [CrossRef]
- McDonald, S.M.; Patton, J.T. Rotavirus VP2 Core Shell Regions Critical for Viral Polymerase Activation. J. Virol. 2011, 85, 3095–3105. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.Q.; Estes, M.K.; Charpilienne, A.; Cohen, J. The N Terminus of Rotavirus VP2 Is Necessary for Encapsidation of VP1 and VP3. J. Virol. 1998, 72, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Yu, X.; Crawford, S.E.; Moreno, R.; Jakana, J.; Sankaran, B.; Anish, R.; Kaundal, S.; Hu, L.; Estes, M.K.; et al. 2.7 Å Cryo-EM Structure of Rotavirus Core Protein VP3, a Unique Capping Machine with a Helicase Activity. Sci. Adv. 2020, 6, eaay6410. [Google Scholar] [CrossRef] [PubMed]
- Bass, D.M.; Baylor, M.R.; Chen, C.; Mackow, E.M.; Bremont, M.; Greenberg, H.B. Liposome-Mediated Transfection of Intact Viral Particles Reveals That Plasma Membrane Penetration Determines Permissivity of Tissue Culture Cells to Rotavirus. J. Clin. Investig. 1992, 90, 2313–2320. [Google Scholar] [CrossRef]
- Charpilienne, A.; Lepault, J.; Rey, F.; Cohen, J. Identification of Rotavirus VP6 Residues Located at the Interface with VP2 That Are Essential for Capsid Assembly and Transcriptase Activity. J. Virol. 2002, 76, 7822–7831. [Google Scholar] [CrossRef]
- Cohen, J.; Laporte, J.; Charpilienne, A.; Scherrer, R. Activation of Rotavirus RNA Polymerase by Calcium Chelation. Arch. Virol. 1979, 60, 177–186. [Google Scholar] [CrossRef]
- Trask, S.D.; Dormitzer, P.R. Assembly of Highly Infectious Rotavirus Particles Recoated with Recombinant Outer Capsid Proteins. J. Virol. 2006, 80, 11293–11304. [Google Scholar] [CrossRef]
- Aoki, S.T.; Settembre, E.C.; Trask, S.D.; Greenberg, H.B.; Harrison, S.C.; Dormitzer, P.R. Structure of Rotavirus Outer-Layer Protein VP7 Bound with a Neutralizing Fab. Science 2009, 324, 1444–1447. [Google Scholar] [CrossRef]
- Clark, S.M.; Roth, J.R.; Clark, M.L.; Barnett, B.B.; Spendlove, R.S. Trypsin Enhancement of Rotavirus Infectivity: Mechanism of Enhancement. J. Virol. 1981, 39, 816–822. [Google Scholar] [CrossRef] [PubMed]
- Estes, M.K.; Graham, D.Y.; Mason, B.B. Proteolytic Enhancement of Rotavirus Infectivity: Molecular Mechanisms. J. Virol. 1981, 39, 879–888. [Google Scholar] [CrossRef] [PubMed]
- Estes, M.K.; Graham, D.Y.; Smith, E.M.; Gerba, C.P. Rotavirus Stability and Inactivation. J. Gen. Virol. 1979, 43, 403–409. [Google Scholar] [CrossRef]
- Helenius, A. Virus Entry: Looking Back and Moving Forward. J. Mol. Biol. 2018, 430, 1853–1862. [Google Scholar] [CrossRef] [PubMed]
- Trask, S.D.; McDonald, S.M.; Patton, J.T. Structural Insights into the Coupling of Virion Assembly and Rotavirus Replication. Nat. Rev. Microbiol. 2012, 10, 165–177. [Google Scholar] [CrossRef]
- Bamford, D.H.; Mindich, L. Viral Molecular Machines: Replication Systems within the Inner Cores of dsRNA Viruses. Virus Res. 2004, 101, 1–100. [Google Scholar] [CrossRef]
- Abdelhakim, A.H.; Salgado, E.N.; Fu, X.; Pasham, M.; Nicastro, D.; Kirchhausen, T.; Harrison, S.C. Structural Correlates of Rotavirus Cell Entry. PLoS Pathog. 2014, 10, e1004355. [Google Scholar] [CrossRef]
- Dormitzer, P.R.; Nason, E.B.; Prasad, B.V.V.; Harrison, S.C. Structural Rearrangements in the Membrane Penetration Protein of a Non-Enveloped Virus. Nature 2004, 430, 1053–1058. [Google Scholar] [CrossRef]
- Sun, X.; Li, D.; Duan, Z. Structural Basis of Glycan Recognition of Rotavirus. Front. Mol. Biosci. 2021, 8, 658029. [Google Scholar] [CrossRef]
- Dormitzer, P.R.; Sun, Z.Y.J.; Wagner, G.; Harrison, S.C. The Rhesus Rotavirus VP4 Sialic Acid Binding Domain Has a Galectin Fold with a Novel Carbohydrate Binding Site. EMBO J. 2002, 21, 885–897. [Google Scholar] [CrossRef]
- Díaz-Salinas, M.A.; Romero, P.; Espinosa, R.; Hoshino, Y.; López, S.; Arias, C.F. The Spike Protein VP4 Defines the Endocytic Pathway Used by Rotavirus to Enter MA104 Cells. J. Virol. 2013, 87, 1658–1663. [Google Scholar] [CrossRef]
- Herrmann, T.; Torres, R.; Salgado, E.N.; Berciu, C.; Stoddard, D.; Nicastro, D.; Jenni, S.; Harrison, S.C. Functional Refolding of the Penetration Protein on a Non-Enveloped Virus. Nature 2021, 590, 666–670. [Google Scholar] [CrossRef] [PubMed]
- Ludert, J.E.; Krishnaney, A.A.; Burns, J.W.; Vo, P.T.; Greenberg, H.B. Cleavage of Rotavirus VP4 In Vivo. J. Gen. Virol. 1996, 77, 391–395. [Google Scholar] [CrossRef]
- Li, Z.; Baker, M.L.; Jiang, W.; Estes, M.K.; Prasad, B.V.V. Rotavirus Architecture at Subnanometer Resolution. J. Virol. 2009, 83, 1754–1766. [Google Scholar] [CrossRef] [PubMed]
- Shah, P.N.M.; Gilchrist, J.B.; Forsberg, B.O.; Burt, A.; Howe, A.; Mosalaganti, S.; Wan, W.; Radecke, J.; Chaban, Y.; Sutton, G.; et al. Characterization of the Rotavirus Assembly Pathway in Situ Using Cryoelectron Tomography. Cell Host Microbe 2023, 31, 604–615.e4. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, J.M.; Chichón, F.J.; Martín-Forero, E.; González-Camacho, F.; Carrascosa, J.L.; Castón, J.R.; Luque, D. New Insights into Rotavirus Entry Machinery: Stabilization of Rotavirus Spike Conformation Is Independent of Trypsin Cleavage. PLoS Pathog. 2014, 10, e1004157. [Google Scholar] [CrossRef]
- Kaljot, K.T.; Shaw, R.D.; Rubin, D.H.; Greenberg, H.B. Infectious Rotavirus Enters Cells by Direct Cell Membrane Penetration, Not by Endocytosis. J. Virol. 1988, 62, 1136–1144. [Google Scholar] [CrossRef]
- Arias, C.F.; López, S. Rotavirus Cell Entry: Not so Simple after All. Curr. Opin. Virol. 2021, 48, 42–48. [Google Scholar] [CrossRef]
- Ciarlet, M.; Crawford, S.E.; Cheng, E.; Blutt, S.E.; Rice, D.A.; Bergelson, J.M.; Estes, M.K. VLA-2 (A2β1) Integrin Promotes Rotavirus Entry into Cells but Is Not Necessary for Rotavirus Attachment. J. Virol. 2002, 76, 1109–1123. [Google Scholar] [CrossRef]
- Ramani, S.; Hu, L.; Venkataram Prasad, B.V.; Estes, M.K. Diversity in Rotavirus-Host Glycan Interactions: A “Sweet” Spectrum. Cell. Mol. Gastroenterol. Hepatol. 2016, 2, 263–273. [Google Scholar] [CrossRef]
- Sánchez-Tacuba, L.; Kawagishi, T.; Feng, N.; Jiang, B.; Ding, S.; Greenberg, H.B. The Role of the VP4 Attachment Protein in Rotavirus Host Range Restriction in an In Vivo Suckling Mouse Model. J. Virol. 2022, 96, e0055022. [Google Scholar] [CrossRef]
- Patra, U.; Mukhopadhyay, U.; Mukherjee, A.; Dutta, S.; Chawla-Sarkar, M. Treading a HOSTile Path: Mapping the Dynamic Landscape of Host Cell–Rotavirus Interactions to Explore Novel Host-Directed Curative Dimensions. Virulence 2021, 12, 1022–1062. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Ding, S.; Fenga, N.; Mooney, N.; Ooia, Y.S.; Ren, L.; Diep, J.; Kelly, M.R.; Yasukawa, L.L.; Patton, J.T.; et al. Drebrin Restricts Rotavirus Entry by Inhibiting Dynamin-Mediated Endocytosis. Proc. Natl. Acad. Sci. USA 2017, 114, E3642–E3651. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, M.; Isa, P.; Sánchez-San Martin, C.; Pérez-Vargas, J.; Espinosa, R.; Arias, C.F.; López, S. Different Rotavirus Strains Enter MA104 Cells through Different Endocytic Pathways: The Role of Clathrin-Mediated Endocytosis. J. Virol. 2010, 84, 9161–9169. [Google Scholar] [CrossRef]
- Díaz-Salinas, M.A.; Silva-Ayala, D.; López, S.; Arias, C.F. Rotaviruses Reach Late Endosomes and Require the Cation-Dependent Mannose-6-Phosphate Receptor and the Activity of Cathepsin Proteases to Enter the Cell. J. Virol. 2014, 88, 4389–4402. [Google Scholar] [CrossRef]
- Silva-Ayala, D.; López, T.; Gutiérrez, M.; Perrimon, N.; López, S.; Arias, C.F.; Estes, M.K. Genome-Wide RNAi Screen Reveals a Role for the ESCRT Complex in Rotavirus Cell Entry. Proc. Natl. Acad. Sci. USA 2013, 110, 10270–10275. [Google Scholar] [CrossRef]
- Wolf, M.; Deal, E.M.; Greenberg, H.B. Rhesus Rotavirus Trafficking during Entry into MA104 Cells Is Restricted to the Early Endosome Compartment. J. Virol. 2012, 86, 4009–4013. [Google Scholar] [CrossRef]
- Soliman, M.; Seo, J.Y.; Kim, D.S.; Kim, J.Y.; Park, J.G.; Alfajaro, M.M.; Baek, Y.B.; Cho, E.H.; Kwon, J.; Choi, J.S.; et al. Activation of PI3K, Akt, and ERK during Early Rotavirus Infection Leads to V-ATPase-Dependent Endosomal Acidification Required for Uncoating. PLoS Pathog. 2018, 14, e1006820. [Google Scholar] [CrossRef] [PubMed]
- Salgado, E.N.; Upadhyayula, S.; Harrison, S.C. Single-Particle Detection of Transcription Following Rotavirus Entry. J. Virol. 2017, 91, e00651-17. [Google Scholar] [CrossRef]
- Salgado, E.N.; Garcia Rodriguez, B.; Narayanaswamy, N.; Krishnan, Y.; Harrison, S.C. Visualization of Calcium Ion Loss from Rotavirus during Cell Entry. J. Virol. 2018, 92, e01327-18. [Google Scholar] [CrossRef]
- Yoder, J.D.; Trask, S.D.; Vo, P.T.; Binka, M.; Feng, N.; Harrison, S.C.; Greenberg, H.B.; Dormitzer, P.R. VP5* Rearranges When Rotavirus Uncoats. J. Virol. 2009, 83, 11372–11377. [Google Scholar] [CrossRef]
- Santiana, M.; Ghosh, S.; Ho, B.A.; Rajasekaran, V.; Du, W.L.; Mutsafi, Y.; De Jésus-Diaz, D.A.; Sosnovtsev, S.V.; Levenson, E.A.; Parra, G.I.; et al. Vesicle-Cloaked Virus Clusters Are Optimal Units for Inter-Organismal Viral Transmission. Cell Host Microbe 2018, 24, 208–220.e8. [Google Scholar] [CrossRef]
- Gallegos, C.O.; Patton, J.T. Characterization of Rotavirus Replication Intermediates: A Model for the Assembly of Single-Shelled Particles. Virology 1989, 172, 616–627. [Google Scholar] [CrossRef]
- Lu, X.; McDonald, S.M.; Tortorici, M.A.; Tao, Y.J.; Vasquez-Del Carpio, R.; Nibert, M.L.; Patton, J.T.; Harrison, S.C. Mechanism for Coordinated RNA Packaging and Genome Replication by Rotavirus Polymerase VP1. Structure 2008, 16, 1678–1688. [Google Scholar] [CrossRef]
- Tortorici, M.A.; Broering, T.J.; Nibert, M.L.; Patton, J.T. Template Recognition and Formation of Initiation Complexes by the Replicase of a Segmented Double-Stranded RNA Virus. J. Biol. Chem. 2003, 278, 32673–32682. [Google Scholar] [CrossRef] [PubMed]
- Patton, J.T. Rotavirus VP1 Alone Specifically Binds to the 3’ End of Viral mRNA, but the Interaction Is Not Sufficient to Initiate Minus-Strand Synthesis. J. Virol. 1996, 70, 7940–7947. [Google Scholar] [CrossRef]
- Papa, G.; Borodavka, A.; Desselberger, U. Viroplasms: Assembly and Functions of Rotavirus Replication Factories. Viruses 2021, 13, 1349. [Google Scholar] [CrossRef] [PubMed]
- Strauss, S.; Acker, J.; Papa, G.; Desiró, D.; Schueder, F.; Borodavka, A.; Jungmann, R. Principles of RNA Recruitment to Viral Ribonucleoprotein Condensates in a Segmented dsRNA Virus. Elife 2023, 12, e68670. [Google Scholar] [CrossRef] [PubMed]
- Patton, J.T.; Silvestri, L.S.; Tortorici, M.A.; Vasquez-Del Carpio, R.; Taraporewala, Z.F. Rotavirus Genome Replication and Morphogenesis: Role of the Viroplasm. Curr. Top. Microbiol. Immunol. 2006, 309, 169–187. [Google Scholar]
- Fabbretti, E.; Afrikanova, I.; Vascotto, F.; Burrone, O.R. Two Non-Structural Rotavirus Proteins, NSP2 and NSP5, Form Viroplasm-like Structures In Vivo. J. Gen. Virol. 1999, 80, 333–339. [Google Scholar] [CrossRef]
- Campagna, M.; Eichwald, C.; Vascotto, F.; Burrone, O.R. RNA Interference of Rotavirus Segment 11 mRNA Reveals the Essential Role of NSP5 in the Virus Replicative Cycle. J. Gen. Virol. 2005, 86, 1481–1487. [Google Scholar] [CrossRef]
- Criglar, J.M.; Anish, R.; Hu, L.; Crawford, S.E.; Sankaran, B.; Prasad, B.V.V.; Estes, M.K. Phosphorylation Cascade Regulates the Formation and Maturation of Rotaviral Replication Factories. Proc. Natl. Acad. Sci. USA 2018, 115, E12015–E12023. [Google Scholar] [CrossRef] [PubMed]
- Criglar, J.M.; Hu, L.; Crawford, S.E.; Hyser, J.M.; Broughman, J.R.; Prasad, B.V.V.; Estes, M.K. A Novel Form of Rotavirus NSP2 and Phosphorylation-Dependent NSP2-NSP5 Interactions Are Associated with Viroplasm Assembly. J. Virol. 2014, 88, 786–798. [Google Scholar] [CrossRef] [PubMed]
- Criglar, J.M.; Crawford, S.E.; Estes, M.K. Plasmid-Based Reverse Genetics for Probing Phosphorylation-Dependent Viroplasm Formation in Rotaviruses. Virus Res. 2021, 291, 198193. [Google Scholar] [CrossRef]
- Gaunt, E.R.; Zhang, Q.; Cheung, W.; Wakelam, M.J.O.; Lever, A.M.L.; Desselberger, U. Lipidome Analysis of Rotavirus-Infected Cells Confirms the Close Interaction of Lipid Droplets with Viroplasms. J. Gen. Virol. 2013, 94, 1576–1586. [Google Scholar] [CrossRef] [PubMed]
- Cheung, W.; Gill, M.; Esposito, A.; Kaminski, C.F.; Courousse, N.; Chwetzoff, S.; Trugnan, G.; Keshavan, N.; Lever, A.; Desselberger, U. Rotaviruses Associate with Cellular Lipid Droplet Components to Replicate in Viroplasms, and Compounds Disrupting or Blocking Lipid Droplets Inhibit Viroplasm Formation and Viral Replication. J. Virol. 2010, 84, 6782–6798. [Google Scholar] [CrossRef]
- Crawford, S.E.; Desselberger, U. Lipid Droplets Form Complexes with Viroplasms and Are Crucial for Rotavirus Replication. Curr. Opin. Virol. 2016, 19, 11–15. [Google Scholar] [CrossRef]
- Criglar, J.M.; Estes, M.K.; Crawford, S.E. Rotavirus-Induced Lipid Droplet Biogenesis Is Critical for Virus Replication. Front. Mater. 2022, 13, 836870. [Google Scholar] [CrossRef]
- Eichwald, C.; Arnoldi, F.; Laimbacher, A.S.; Schraner, E.M.; Fraefel, C.; Wild, P.; Burrone, O.R.; Ackermann, M. Rotavirus Viroplasm Fusion and Perinuclear Localization Are Dynamic Processes Requiring Stabilized Microtubules. PLoS ONE 2012, 7, e47947. [Google Scholar] [CrossRef]
- Campagna, M.; Marcos-Villar, L.; Arnoldi, F.; de la Cruz-Herrera, C.F.; Gallego, P.; González-Santamaría, J.; González, D.; Lopitz-Otsoa, F.; Rodriguez, M.S.; Burrone, O.R.; et al. Rotavirus Viroplasm Proteins Interact with the Cellular SUMOylation System: Implications for Viroplasm-Like Structure Formation. J. Virol. 2013, 87, 807–817. [Google Scholar] [CrossRef] [PubMed]
- Chandra, P.; Banerjee, S.; Saha, P.; Chawla-Sarkar, M.; Patra, U. Sneaking into the Viral Safe-Houses: Implications of Host Components in Regulating Integrity and Dynamics of Rotaviral Replication Factories. Front. Cell. Infect. Microbiol. 2022, 12, 977799. [Google Scholar] [CrossRef] [PubMed]
- Geiger, F.; Acker, J.; Papa, G.; Wang, X.; Arter, W.E.; Saar, K.L.; Erkamp, N.A.; Qi, R.; Bravo, J.P.K.; Strauss, S.; et al. Liquid–Liquid Phase Separation Underpins the Formation of Replication Factories in Rotaviruses. EMBO J. 2021, 40, e107711. [Google Scholar] [CrossRef] [PubMed]
- Suárez, Y.G.; Martínez, J.L.; Hernández, D.T.; Hernández, H.O.; Pérez-Delgado, A.; Méndez, M.; Wood, C.D.; Rendon-Mancha, J.M.; Silva-Ayala, D.; López, S.; et al. Nanoscale Organization of Rotavirus Replication Machineries. Elife 2019, 8, e42906. [Google Scholar] [CrossRef]
- Banani, S.F.; Lee, H.O.; Hyman, A.A.; Rosen, M.K. Biomolecular Condensates: Organizers of Cellular Biochemistry. Nat. Rev. Mol. Cell Biol. 2017, 18, 285–298. [Google Scholar] [CrossRef]
- Borodavka, A.; Desselberger, U.; Patton, J.T. Genome Packaging in Multi-Segmented dsRNA Viruses: Distinct Mechanisms with Similar Outcomes. Curr. Opin. Virol. 2018, 33, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Desselberger, U. What Are the Limits of the Packaging Capacity for Genomic RNA in the Cores of Rotaviruses and of Other Members of the Reoviridae? Virus Res. 2020, 276, 197822. [Google Scholar] [CrossRef]
- Moreno-Contreras, J.; Sánchez-Tacuba, L.; Arias, C.F.; López, S. Mature Rotavirus Particles Contain Equivalent Amounts of 7meGpppG-Capped and Noncapped Viral Positive-Sense RNAs. J. Virol. 2022, 96, e0115122. [Google Scholar] [CrossRef]
- Borodavka, A.; Dykeman, E.C.; Schrimpf, W.; Lamb, D.C. Protein-Mediated RNA Folding Governs Sequence-Specific Interactions between Rotavirus Genome Segments. Elife 2017, 6, e27453. [Google Scholar] [CrossRef]
- Sutton, G.; Sun, D.; Fu, X.; Kotecha, A.; Hecksel, C.W.; Clare, D.K.; Zhang, P.; Stuart, D.I.; Boyce, M. Assembly Intermediates of Orthoreovirus Captured in the Cell. Nat. Commun. 2020, 11, 4445. [Google Scholar] [CrossRef]
- Long, C.P.; McDonald, S.M. Rotavirus Genome Replication: Some Assembly Required. PLoS Pathog. 2017, 13, e1006242. [Google Scholar] [CrossRef]
- Papa, G.; Venditti, L.; Braga, L.; Schneider, E.; Giacca, M.; Petris, G.; Burrone, O.R. CRISPR-Csy4-Mediated Editing of Rotavirus Double-Stranded RNA Genome. Cell Rep. 2020, 32, 108205. [Google Scholar] [CrossRef] [PubMed]
- Ayala-Breton, C.; Arias, M.; Espinosa, R.; Romero, P.; Arias, C.F.; López, S. Analysis of the Kinetics of Transcription and Replication of the Rotavirus Genome by RNA Interference. J. Virol. 2009, 83, 8819–8831. [Google Scholar] [CrossRef] [PubMed]
- Stacy-Phipps, S.; Patton, J.T. Synthesis of Plus- and Minus-Strand RNA in Rotavirus-Infected Cells. J. Virol. 1987, 61, 3479–3484. [Google Scholar] [CrossRef] [PubMed]
- Crawford, S.E.; Hyser, J.M.; Utama, B.; Estes, M.K. Autophagy Hijacked through Viroporin-Activated Calcium/Calmodulin-Dependent Kinase Kinase-β Signaling Is Required for Rotavirus Replication. Proc. Natl. Acad. Sci. USA 2012, 109, E3405–E3413. [Google Scholar] [CrossRef]
- Crawford, S.E.; Criglar, J.M.; Liu, Z.; Broughman, J.R.; Estes, M.K. COPII Vesicle Transport Is Required for Rotavirus NSP4 Interaction with the Autophagy Protein LC3 II and Trafficking to Viroplasms. J. Virol. 2019, 94, e01341-19. [Google Scholar] [CrossRef]
- Jensen, D.; Schekman, R. COPII-Mediated Vesicle Formation at a Glance. J. Cell Sci. 2011, 124 Pt 1, 1–4. [Google Scholar] [CrossRef]
- Peña-Martinez, C.; Rickman, A.D.; Heckmann, B.L. Beyond Autophagy: LC3-Associated Phagocytosis and Endocytosis. Sci. Adv. 2022, 8, eabn1702. [Google Scholar] [CrossRef]
- Taylor, J.A.; O’Brien, J.A.; Yeager, M. The Cytoplasmic Tail of NSP4, the Endoplasmic Reticulum-Localized Non-Structural Glycoprotein of Rotavirus, Contains Distinct Virus Binding and Coiled Coil Domains. EMBO J. 1996, 15, 4469–4476. [Google Scholar] [CrossRef]
- O’Brien, J.A.; Taylor, J.A.; Bellamy, A.R. Probing the Structure of Rotavirus NSP4: A Short Sequence at the Extreme C Terminus Mediates Binding to the Inner Capsid Particle. J. Virol. 2000, 74, 5388–5394. [Google Scholar] [CrossRef]
- Au, K.S.; Mattion, N.M.; Estes, M.K. A Subviral Particle Binding Domain on the Rotavirus Nonstructural Glycoprotein Ns28. Virology 1993, 194, 665–673. [Google Scholar] [CrossRef]
- Maass, D.R.; Atkinson, P.H. Rotavirus Proteins VP7, NS28, and VP4 Form Oligomeric Structures. J. Virol. 1990, 64, 2632–2641. [Google Scholar] [CrossRef]
- Musalem, C.; Espejo, R.T. Release of Progeny Virus from Cells Infected with Simian Rotavirus SA11. J. Gen. Virol. 1985, 66, 2715–2724. [Google Scholar] [CrossRef] [PubMed]
- Jourdan, N.; Maurice, M.; Delautier, D.; Quero, A.M.; Servin, A.L.; Trugnan, G. Rotavirus Is Released from the Apical Surface of Cultured Human Intestinal Cells through Nonconventional Vesicular Transport That Bypasses the Golgi Apparatus. J. Virol. 1997, 71, 8268–8278. [Google Scholar] [CrossRef] [PubMed]
- Kerviel, A.; Zhang, M.; Altan-Bonnet, N. A New Infectious Unit: Extracellular Vesicles Carrying Virus Populations. Annu. Rev. Cell Dev. Biol. 2021, 37, 171–197. [Google Scholar] [CrossRef]
- Vora, A.; Zhou, W.; Londono-Renteria, B.; Woodson, M.; Sherman, M.B.; Colpitts, T.M.; Neelakanta, G.; Sultana, H. Arthropod EVs Mediate Dengue Virus Transmission through Interaction with a Tetraspanin Domain Containing Glycoprotein Tsp29Fb. Proc. Natl. Acad. Sci. USA 2018, 115, E6604–E6613. [Google Scholar] [CrossRef]
- Labadie, T.; Roy, P. A Non-Enveloped Arbovirus Released in Lysosome-Derived Extracellular Vesicles Induces Super-Infection Exclusion. PLoS Pathog. 2020, 16, e1009015. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Woodson, M.; Neupane, B.; Bai, F.; Sherman, M.B.; Choi, K.H.; Neelakanta, G.; Sultana, H. Exosomes Serve as Novel Modes of Tick-Borne Flavivirus Transmission from Arthropod to Human Cells and Facilitates Dissemination of Viral RNA and Proteins to the Vertebrate Neuronal Cells. PLoS Pathog. 2018, 14, e1006764. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Du, W.; Hagemeijer, M.C.; Takvorian, P.M.; Pau, C.; Cali, A.; Brantner, C.A.; Stempinski, E.S.; Connelly, P.S.; Ma, H.C.; et al. Phosphatidylserine Vesicles Enable Efficient En Bloc Transmission of Enteroviruses. Cell 2015, 160, 619–630. [Google Scholar] [CrossRef]
- Ramakrishnaiah, V.; Thumann, C.; Fofana, I.; Habersetzer, F.; Pan, Q.; De Ruiter, P.E.; Willemsen, R.; Demmers, J.A.A.; Raj, V.S.; Jenster, G.; et al. Exosome-Mediated Transmission of Hepatitis C Virus between Human Hepatoma Huh7.5 Cells. Proc. Natl. Acad. Sci. USA 2013, 110, 13109–13113. [Google Scholar] [CrossRef]
- Altan-Bonnet, N.; Perales, C.; Domingo, E. Extracellular Vesicles: Vehicles of En Bloc Viral Transmission. Virus Res. 2019, 265, 143–149. [Google Scholar] [CrossRef]
- Altan-Bonnet, N. Extracellular Vesicles Are the Trojan Horses of Viral Infection. Curr. Opin. Microbiol. 2016, 32, 77–81. [Google Scholar] [CrossRef]
- Shirogane, Y.; Watanabe, S.; Yanagi, Y. Cooperation between Different Variants: A Unique Potential for Virus Evolution. Virus Res. 2019, 264, 68–73. [Google Scholar] [CrossRef]
- Iša, P.; Pérez-Delgado, A.; Quevedo, I.R.; López, S.; Arias, C.F. Rotaviruses Associate with Distinct Types of Extracellular Vesicles. Viruses 2020, 12, 763. [Google Scholar] [CrossRef] [PubMed]
- Carafoli, E.; Krebs, J. Why Calcium? How Calcium Became the Best Communicator. J. Biol. Chem. 2016, 291, 20849–20857. [Google Scholar] [CrossRef]
- Zhou, Y.; Frey, T.K.; Yang, J.J. Viral Calciomics: Interplays between Ca2+ and Virus. Cell Calcium 2009, 46, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Gajardo, R.; Vende, P.; Poncet, D.; Cohen, J. Two Proline Residues Are Essential in the Calcium-Binding Activity of Rotavirus VP7 Outer Capsid Protein. J. Virol. 1997, 71, 2211–2216. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, M.C.; Charpilienne, A.; Liprandi, F.; Gajardo, R.; Michelangeli, F.; Cohen, J. The Concentration of Ca2+ That Solubilizes Outer Capsid Proteins from Rotavirus Particles Is Dependent on the Strain. J. Virol. 1996, 70, 4877–4883. [Google Scholar] [CrossRef]
- Bican, P.; Cohen, J.; Charpilienne, A.; Scherrer, R. Purification and Characterization of Bovine Rotavirus Cores. J. Virol. 1982, 43, 1113–1117. [Google Scholar] [CrossRef]
- Chandran, K.; Zhang, X.; Olson, N.H.; Walker, S.B.; Chappell, J.D.; Dermody, T.S.; Baker, T.S.; Nibert, M.L. Complete In Vitro Assembly of the Reovirus Outer Capsid Produces Highly Infectious Particles Suitable for Genetic Studies of the Receptor-Binding Protein. J. Virol. 2001, 75, 5335–5342. [Google Scholar] [CrossRef]
- Chandran, K.; Parker, J.S.L.; Ehrlich, M.; Kirchhausen, T.; Nibert, M.L. The δ Region of Outer Capsid Protein μ1 Undergoes Conformational Change and Release from Reovirus Particles during Cell Entry. J. Virol. 2003, 77, 13361–13375. [Google Scholar] [CrossRef]
- Odegard, A.L.; Chandran, K.; Liemann, S.; Harrison, S.C.; Nibert, M.L. Disulfide Bonding among Μ1 Trimers in Mammalian Reovirus Outer Capsid: A Late and Reversible Step in Virion Morphogenesis. J. Virol. 2003, 77, 5389–5400. [Google Scholar] [CrossRef] [PubMed]
- Odegard, A.L.; Chandran, K.; Zhang, X.; Parker, J.S.L.; Baker, T.S.; Nibert, M.L. Putative Autocleavage of Outer Capsid Protein Μ1, Allowing Release of Myristoylated Peptide Μ1N during Particle Uncoating, Is Critical for Cell Entry by Reovirus. J. Virol. 2004, 78, 8732–8745. [Google Scholar] [CrossRef] [PubMed]
- Komoto, S.; Fukuda, S.; Ide, T.; Ito, N.; Sugiyama, M.; Yoshikawa, T.; Murata, T.; Taniguchi, K. Generation of Recombinant Rotaviruses Expressing Fluorescent Proteins by Using an Optimized Reverse Genetics System. J. Virol. 2018, 92, e00588-18. [Google Scholar] [CrossRef] [PubMed]
- Komoto, S.; Fukuda, S.; Murata, T.; Taniguchi, K. Human Rotavirus Reverse Genetics Systems to Study Viral Replication and Pathogenesis. Viruses 2021, 13, 1791. [Google Scholar] [CrossRef]
- Chen, D.; Ramig, R.F. Rescue of Infectivity by in Vitro Transcapsidation of Rotavirus Single- Shelled Particles. Virology 1993, 192, 422–429. [Google Scholar] [CrossRef]
- Grgacic, E.V.L.; Anderson, D.A. Virus-like Particles: Passport to Immune Recognition. Methods 2006, 40, 60–65. [Google Scholar] [CrossRef]
- Crawford, S.E.; Labbé, M.; Cohen, J.; Burroughs, M.H.; Zhou, Y.J.; Estes, M.K.; Labbe, M.; Cohen, J.; Burroughs, M.H.; Zhou, Y.J.; et al. Characterization of Virus-like Particles Produced by the Expression of Rotavirus Capsid Proteins in Insect Cells. J. Virol. 1994, 68, 5945–5952. [Google Scholar] [CrossRef]
- Rodríguez-Limas, W.A.; Tyo, K.E.J.; Nielsen, J.; Ramírez, O.T.; Palomares, L.A. Molecular and Process Design for Rotavirus-like Particle Production in Saccharomyces cerevisiae. Microb Cell Fact 2011, 10, 33. [Google Scholar] [CrossRef]
- Yang, Y.M.; Li, X.; Yang, H.; Qian, Y.; Zhang, Y.; Fang, R.X.; Chen, X.Y. Immunogenicity and Virus-like Particle Formation of Rotavirus Capsid Proteins Produced in Transgenic Plants. Sci. China Life Sci. 2011, 54, 82–89. [Google Scholar] [CrossRef]
- Saldana, S.; Esquivel Guadarrama, F.; Olivera Flores Tde, J.; Arias, N.; Lopez, S.; Arias, C.; Ruiz-Medrano, R.; Mason, H.; Mor, T.; Richter, L.; et al. Production of Rotavirus-like Particles in Tomato (Lycopersicon esculentum L.) Fruit by Expression of Capsid Proteins VP2 and VP6 and Immunological Studies. Viral Immunol. 2006, 19, 42–53. [Google Scholar] [CrossRef]
- Lee, J.M.; Chung, H.Y.; Kim, K.I.; Yoo, K.H.; Hwang-Bo, J.; Chung, I.S.; Park, J.H. Synthesis of Double-Layered Rotavirus-like Particles Using Internal Ribosome Entry Site Vector System in Stably-Transformed Drosophila melanogaster. Biotechnol. Lett. 2011, 33, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Laimbacher, A.S.; Esteban, L.E.; Castello, A.A.; Abdusetir Cerfoglio, J.C.; Argüelles, M.H.; Glikmann, G.; D’Antuono, A.; Mattion, N.; Berois, M.; Arbiza, J.; et al. HSV-1 Amplicon Vectors Launch the Production of Heterologous Rotavirus-like Particles and Induce Rotavirus-Specific Immune Responses in Mice. Mol. Ther. 2012, 20, 1810–1820. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Lin, H.; Zhang, Y.; Li, M.; Wang, D.; Che, Y.; Zhu, Y.; Li, S.; Zhang, J.; Ge, S.; et al. Improved Characteristics and Protective Efficacy in an Animal Model of E. coli-Derived Recombinant Double-Layered Rotavirus Virus-like Particles. Vaccine 2014, 32, 1921–1931. [Google Scholar] [CrossRef] [PubMed]
- Pêra, F.F.P.G.; Mutepfa, D.L.R.; Khan, A.M.; Els, J.H.; Mbewana, S.; Van Dijk, A.A.A.; Rybicki, E.P.; Hitzeroth, I.I. Engineering and Expression of a Human Rotavirus Candidate Vaccine in Nicotiana benthamiana. Virol. J. 2015, 12, 205. [Google Scholar] [CrossRef]
- Labbe, M.; Charpilienne, A.; Crawford, S.E.; Estes, M.K.; Cohen, J. Expression of Rotavirus VP2 Produces Empty Corelike Particles. J. Virol. 1991, 65, 2946–2952. [Google Scholar] [CrossRef]
- Zeng, C.Q.; Wentz, M.J.; Cohen, J.; Estes, M.K.; Ramig, R.F. Characterization and Replicase Activity of Double-Layered and Single-Layered Rotavirus-like Particles Expressed from Baculovirus Recombinants. J. Virol. 1996, 70, 2736–2742. [Google Scholar] [CrossRef]
- Vieira, H.L.A.; Estêvão, C.; Roldão, A.; Peixoto, C.C.; Sousa, M.F.Q.; Cruz, P.E.; Carrondo, M.J.T.; Alves, P.M. Triple Layered Rotavirus VLP Production: Kinetics of Vector Replication, MRNA Stability and Recombinant Protein Production. J. Biotechnol. 2005, 120, 72–82. [Google Scholar] [CrossRef]
- Jayaram, H.; Estes, M.K.; Prasad, B.V.V. Emerging Themes in Rotavirus Cell Entry, Genome Organization, Transcription and Replication. Virus Res. 2004, 101, 67–81. [Google Scholar] [CrossRef]
- Estes, M.K.; Crawford, S.E.; Penaranda, M.E.; Petrie, B.L.; Burns, J.W.; Chan, W.K.; Ericson, B.; Smith, G.E.; Summers, M.D. Synthesis and Immunogenicity of the Rotavirus Major Capsid Antigen Using a Baculovirus Expression System. J. Virol. 1987, 61, 1488–1494. [Google Scholar] [CrossRef]
- Lepault, J.; Petitpas, I.; Erk, I.; Navaza, J.; Bigot, D.; Dona, M.; Vachette, P.; Cohen, J.; Rey, F.A. Structural Polymorphism of the Major Capsid Protein of Rotavirus. EMBO J. 2001, 20, 1498–1507. [Google Scholar] [CrossRef]
- Ready, K.F.M.; Sabara, M. In Vitro Assembly of Bovine Rotavirus Nucleocapsid Protein. Virology 1987, 157, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Chang, K.O.; Kim, W.Y.; Saif, L.J. Production of Hybrid Double- or Triple-Layered Virus-like Particles of Group A and C Rotaviruses Using a Baculovirus Expression System. Virology 2002, 302, 1–8. [Google Scholar] [CrossRef]
- Smith, G.E.; Summers, M.D.; Fraser, M.J. Production of Human Beta Interferon in Insect Cells Infected with a Baculovirus Expression Vector. Mol. Cell Biol. 1983, 3, 2156–2165. [Google Scholar]
- Jarvis, D.L. Baculovirus-Insect Cell Expression Systems. Methods Enzymol. 2009, 463, 191–222. [Google Scholar] [PubMed]
- Felberbaum, R.S. The Baculovirus Expression Vector System: A Commercial Manufacturing Platform for Viral Vaccines and Gene Therapy Vectors. Biotechnol. J. 2015, 10, 702–714. [Google Scholar] [CrossRef] [PubMed]
- Irons, S.L.; Chambers, A.C.; Lissina, O.; King, L.A.; Possee, R.D. Protein Production Using the Baculovirus Expression System. Curr. Protoc. Protein Sci. 2018, 91, 5.5.1–5.5.22. [Google Scholar] [CrossRef]
- Beljelarskaya, S.N. Baculovirus Expression Systems for Production of Recombinant Proteins in Insect and Mammalian Cells. Mol. Biol. 2011, 45, 123–138. [Google Scholar] [CrossRef]
- Clark, K.B.; Lin, S.C.; Humphrey, C.; Foytich, K.; Esona, M.; Wang, Y.; Liu, M.; Jiang, B. Expression and Characterization of Human Group C Rotavirus Virus-like Particles in Insect Cells. Virology 2009, 387, 267–272. [Google Scholar] [CrossRef]
- Zhou, H.; Guo, L.; Wang, M.; Qu, J.; Zhao, Z.; Wang, J.; Hung, T. Prime Immunization with Rotavirus VLP 2/6 Followed by Boosting with an Adenovirus Expressing VP6 Induces Protective Immunization against Rotavirus in Mice. Virol. J. 2011, 8, 3. [Google Scholar] [CrossRef]
- O’Neal, C.M.; Crawford, S.E.; Estes, M.K.; Conner, M.E. Rotavirus Virus-like Particles Administered Mucosally Induce Protective Immunity. J. Virol. 1997, 71, 8707–8717. [Google Scholar] [CrossRef]
- Parez, N.; Fourgeux, C.; Mohamed, A.; Dubuquoy, C.; Pillot, M.; Dehee, A.; Charpilienne, A.; Poncet, D.; Schwartz-Cornil, I.; Garbarg-Chenon, A. Rectal Immunization with Rotavirus Virus-Like Particles Induces Systemic and Mucosal Humoral Immune Responses and Protects Mice against Rotavirus Infection. J. Virol. 2006, 80, 1752–1761. [Google Scholar] [CrossRef] [PubMed]
- Agnello, D.; Hervé, C.A.; Lavaux, A.; Darniot, M.; Guillon, P.; Charpilienne, A.; Pothier, P. Intrarectal Immunization with Rotavirus 2/6 Virus-Like Particles Induces an Antirotavirus Immune Response Localized in the Intestinal Mucosa and Protects against Rotavirus Infection in Mice. J. Virol. 2006, 80, 3823–3832. [Google Scholar] [CrossRef] [PubMed]
- Changotra, H.; Vij, A. Rotavirus Virus-like Particles (RV-VLPs) Vaccines: An Update. Rev. Med. Virol. 2017, 27, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kushnir, N.; Streatfield, S.J.; Yusibov, V. Virus-like Particles as a Highly Efficient Vaccine Platform: Diversity of Targets and Production Systems and Advances in Clinical Development. Vaccine 2012, 31, 58–83. [Google Scholar] [CrossRef]
- Noad, R.; Roy, P. Virus-like Particles as Immunogens. Trends Microbiol. 2003, 11, 438–444. [Google Scholar] [CrossRef]
- Heinimäki, S.; Hankaniemi, M.M.; Sioofy-Khojine, A.B.; Laitinen, O.H.; Hyöty, H.; Hytönen, V.P.; Vesikari, T.; Blazevic, V. Combination of Three Virus-Derived Nanoparticles as a Vaccine against Enteric Pathogens; Enterovirus, Norovirus and Rotavirus. Vaccine 2019, 37, 7509–7518. [Google Scholar] [CrossRef]
- Fiore, L.; Greenberg, H.B.; Mackow, E.R. The VP8 Fragment of VP4 Is the Rhesus Rotavirus Hemagglutinin. Virology 1991, 181, 553–563. [Google Scholar] [CrossRef]
- Zhao, Q.; Chen, W.; Chen, Y.; Zhang, L.; Zhang, J.; Zhang, Z. Self-Assembled Virus-like Particles from Rotavirus Structural Protein VP6 for Targeted Drug Delivery. Bioconjug. Chem. 2011, 22, 346–352. [Google Scholar] [CrossRef]
- Vicente, T.; Roldão, A.; Peixoto, C.; Carrondo, M.J.T.; Alves, P.M. Large-Scale Production and Purification of VLP-Based Vaccines. J. Invertebr. Pathol. 2011, 107, S42–S48. [Google Scholar] [CrossRef]
- Trugnan, G.; Cortes-Perez, N.G.; Sapin, C.; Jaffrelo, L.; Daou, S.; Grill, J.P.; Langella, P.; Seksik, P.; Beaugerie, L.; Chwetzoff, S. Rotavirus-like Particles: A Novel Nanocarrier for the Gut. J. Biomed. Biotechnol. 2010, 2010. [Google Scholar]
- Peralta, A.; Molinari, P.; Taboga, O. Chimeric Recombinant Rotavirus-like Particles as a Vehicle for the Display of Heterologous Epitopes. Virol. J. 2009, 6, 1–9. [Google Scholar] [CrossRef]
- Charpilienne, A.; Nejmeddine, M.; Berois, M.; Parez, N.; Neumann, E.; Hewat, E.; Trugnan, G.; Cohen, J. Individual Rotavirus-like Particles Containing 120 Molecules of Fluorescent Protein Are Visible in Living Cells. J. Biol. Chem. 2001, 276, 29361–29367. [Google Scholar] [CrossRef] [PubMed]
- Plascencia-Villa, G.; Mena, J.A.; Castro-Acosta, R.M.; Fabián, J.C.; Ramírez, O.T.; Palomares, L.A. Strategies for the Purification and Characterization of Protein Scaffolds for the Production of Hybrid Nanobiomaterials. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2011, 879, 1105–1111. [Google Scholar] [CrossRef]
- Kanai, Y.; Komoto, S.; Kawagishi, T.; Nouda, R.; Nagasawa, N.; Onishi, M.; Matsuura, Y.; Taniguchi, K.; Kobayashi, T. Entirely Plasmid-Based Reverse Genetics System for Rotaviruses. Proc. Natl. Acad. Sci. USA 2017, 114, 2343–2348. [Google Scholar] [CrossRef]
- Komoto, S.; Sasaki, J.; Taniguchi, K. Reverse Genetics System for Introduction of Site-Specific Mutations into the Double-Stranded RNA Genome of Infectious Rotavirus. Proc. Natl. Acad. Sci. USA 2006, 103, 4646–4651. [Google Scholar] [CrossRef]
- Komoto, S.; Fukuda, S.; Murata, T.; Taniguchi, K. Reverse Genetics System for Human Rotaviruses. Microbiol. Immunol. 2020, 64, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Mignot, T.; Nollmann, M. Biology across Scales: From Atomic Processes to Bacterial Communities through the Lens of the Microscope. FEMS Microbiol. Rev. 2021, 45, fuab009. [Google Scholar] [CrossRef]
- Philip, A.A.; Patton, J.T. Expression of Separate Heterologous Proteins from the Rotavirus NSP3 Genome Segment Using a Translational 2A Stop-Restart Element. J. Virol. 2020, 94. [Google Scholar] [CrossRef] [PubMed]
- Philip, A.A.; Patton, J.T. Rotavirus as an Expression Platform of Domains of the SARS-CoV-2 Spike Protein. Vaccines 2021, 9, 449. [Google Scholar] [CrossRef]
- Kawagishi, T.; Sánchez-Tacuba, L.; Feng, N.; Costantini, V.P.; Tan, M.; Jiang, X.; Green, K.Y.; Vinjé, J.; Ding, S.; Greenberg, H.B. Mucosal and Systemic Neutralizing Antibodies to Norovirus Induced in Infant Mice Orally Inoculated with Recombinant Rotaviruses. Proc. Natl. Acad. Sci. USA 2023, 120, e2214421120. [Google Scholar] [CrossRef]
- Zhu, Y.; Sánchez-Tacuba, L.; Hou, G.; Kawagishi, T.; Feng, N.; Greenberg, H.B.; Ding, S. A Recombinant Murine-like Rotavirus with Nano-Luciferase Expression Reveals Tissue Tropism, Replication Dynamics, and Virus Transmission. Front. Immunol. 2022, 13. [Google Scholar] [CrossRef]
- Yamasaki, M.; Kanai, Y.; Wakamura, Y.; Kotaki, T.; Minami, S.; Nouda, R.; Nurdin, J.A.; Kobayashi, T. Characterization of Sialic Acid-Independent Simian Rotavirus Mutants in Viral Infection and Pathogenesis. J. Virol. 2023, 97, e0139722. [Google Scholar] [CrossRef]
- Kanai, Y.; Kobayashi, T. Rotavirus Reverse Genetics Systems: Development and Application. Virus Res. 2021, 295, 198296. [Google Scholar] [CrossRef] [PubMed]
- Kanai, Y.; Kobayashi, T. FAST Proteins: Development and Use of Reverse Genetics Systems for Reoviridae Viruses. Annu. Rev. Virol. 2021, 8, 515–536. [Google Scholar] [CrossRef]
- Kanai, Y.; Onishi, M.; Kawagishi, T.; Pannacha, P.; Nurdin, J.A.; Nouda, R.; Yamasaki, M.; Lusiany, T.; Khamrin, P.; Okitsu, S.; et al. Reverse Genetics Approach for Developing Rotavirus Vaccine Candidates Carrying VP4 and VP7 Genes Cloned from Clinical Isolates of Human Rotavirus. J. Virol. 2020, 95, e01374-20. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Patton, J.T.; Desselberger, U. Species A Rotavirus Reverse Genetics: Achievements and Prospects. Virus Res. 2021, 306, 198583. [Google Scholar] [CrossRef] [PubMed]
- Desselberger, U. Potential of Plasmid Only Based Reverse Genetics of Rotavirus for the Development of Next-Generation Vaccines. Curr. Opin. Virol. 2020, 44, 1–6. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Asensio-Cob, D.; Rodríguez, J.M.; Luque, D. Rotavirus Particle Disassembly and Assembly In Vivo and In Vitro. Viruses 2023, 15, 1750. https://doi.org/10.3390/v15081750
Asensio-Cob D, Rodríguez JM, Luque D. Rotavirus Particle Disassembly and Assembly In Vivo and In Vitro. Viruses. 2023; 15(8):1750. https://doi.org/10.3390/v15081750
Chicago/Turabian StyleAsensio-Cob, Dunia, Javier M. Rodríguez, and Daniel Luque. 2023. "Rotavirus Particle Disassembly and Assembly In Vivo and In Vitro" Viruses 15, no. 8: 1750. https://doi.org/10.3390/v15081750