
Genomic and Pathologic Characterization of the First FAdV-C Serotype 4 Isolate from Black-Necked Crane

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fecal Samples from Wading Birds
2.2. PCR Protocols for Virus Identification
2.3. Virus Isolation Using LMH and Determination of TCID50
2.4. Determination of Complete Genome
2.5. Sequence Aligment Analysis
2.6. Viral Challenge Study with Mallard Ducks
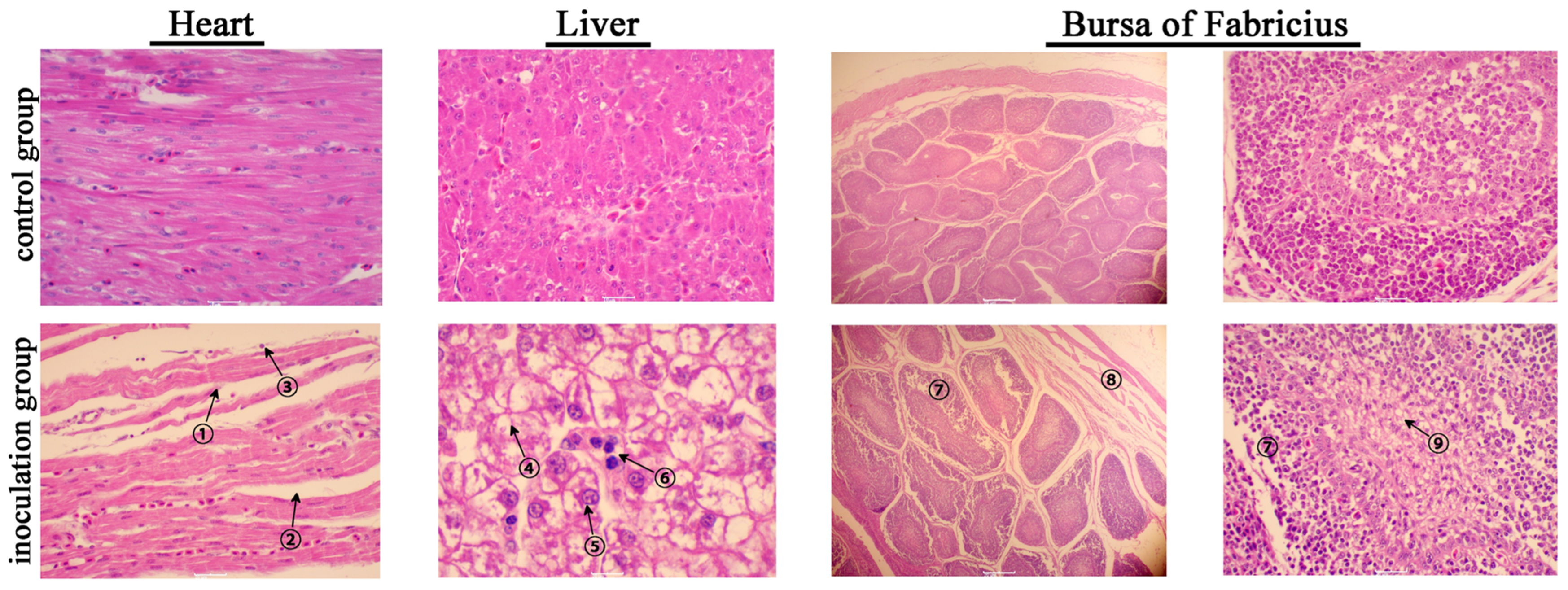
2.7. Histopathologic Examination
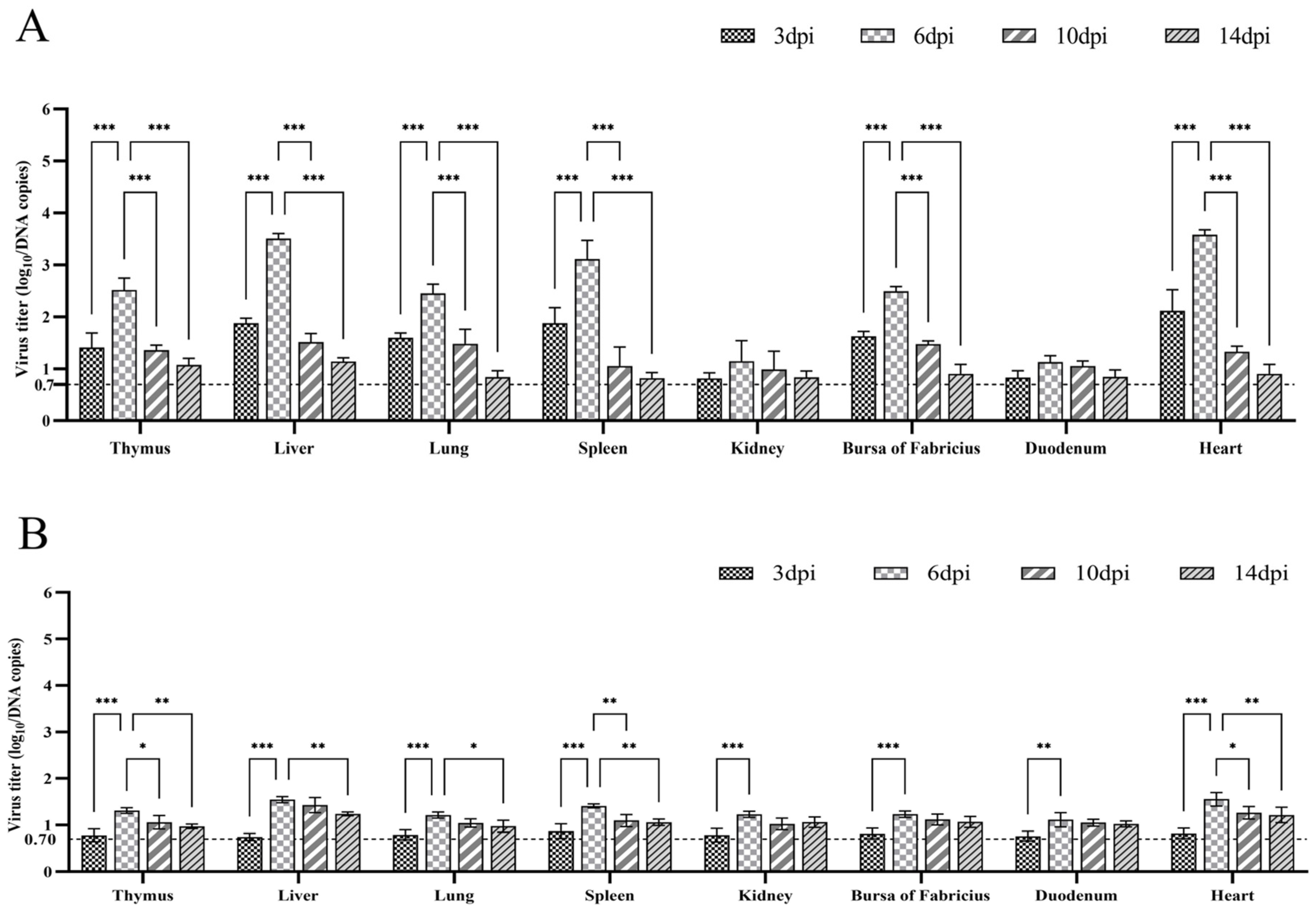
2.8. Quantification of Viral Titers in Tissues
2.9. Statistical Analysis
3. Results
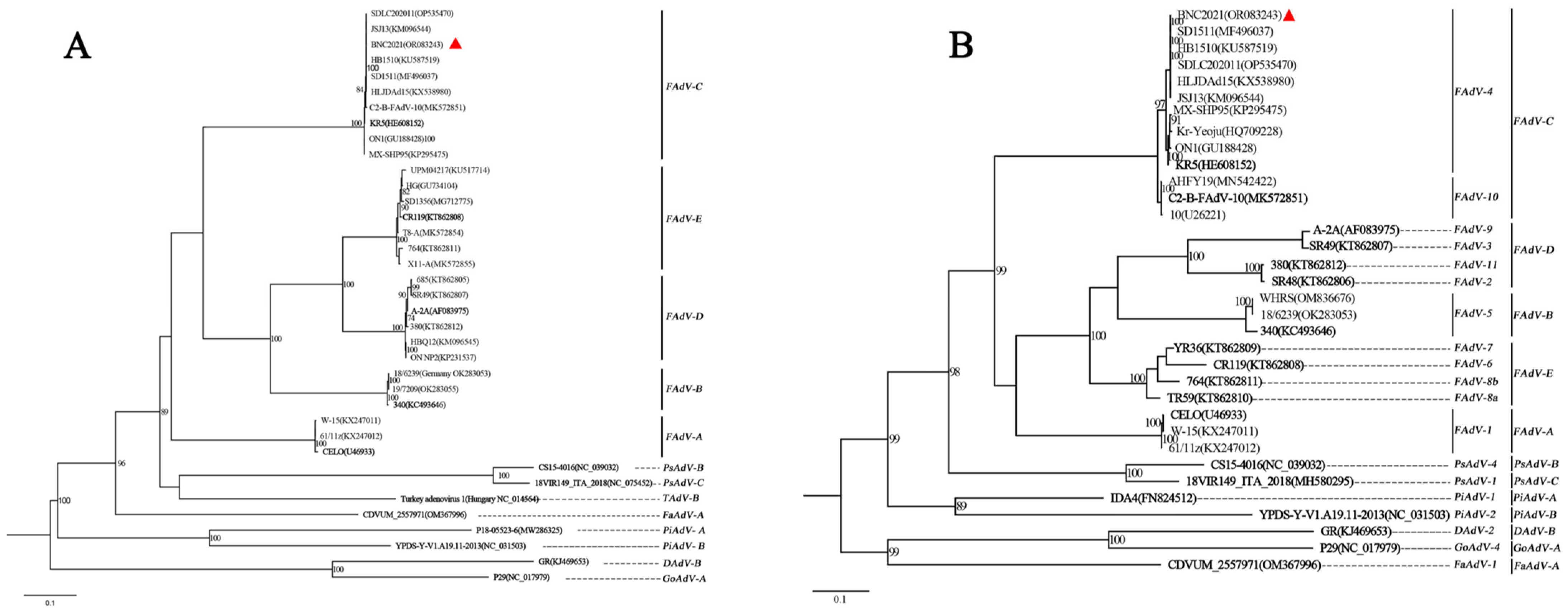
3.1. Identification of the Isolated Strain BNC2021
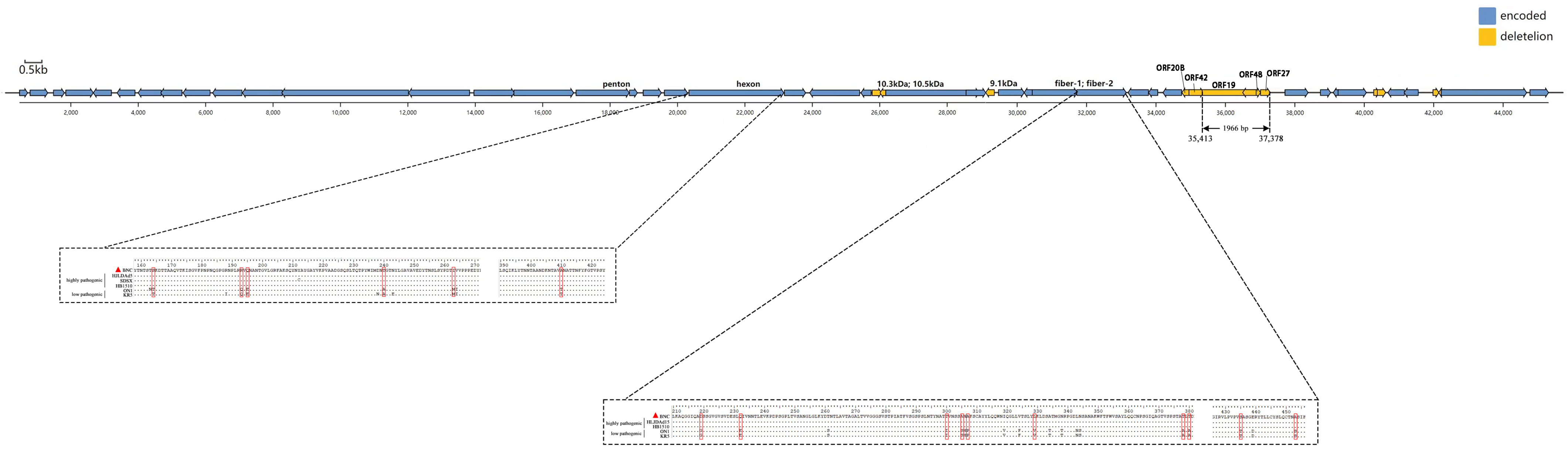
3.2. Genomic Analysis of BNC2021
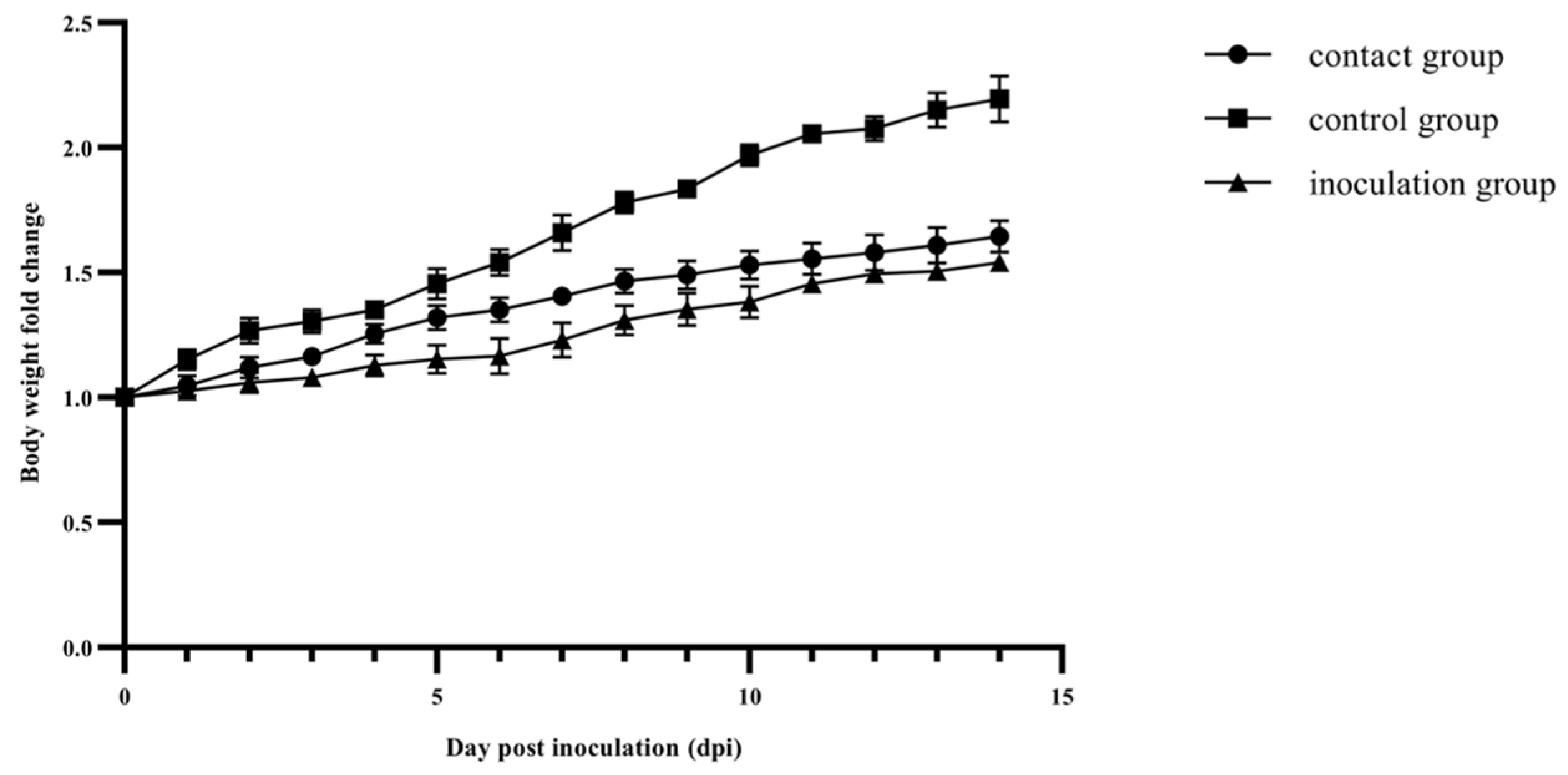
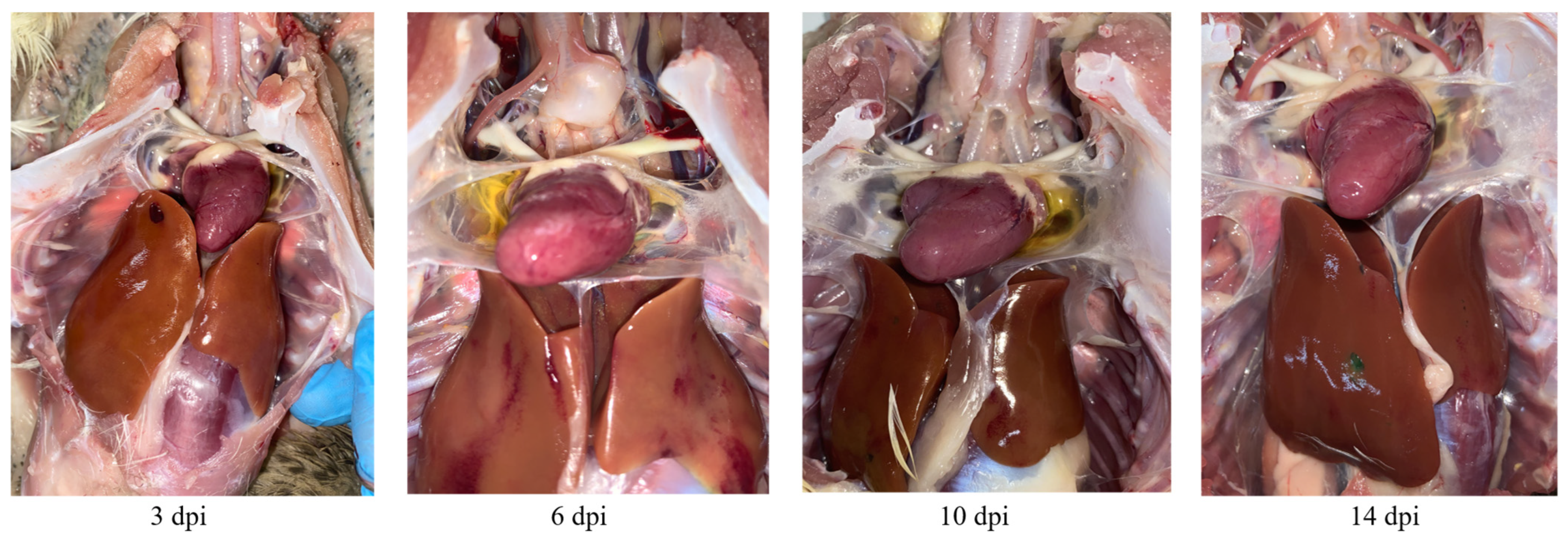
3.3. Pathogenicity Assessment of BNC2021 in Mallard Ducks
3.4. Viral Titers in Tissues from Mallard Ducks Challenged with BNC2021
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Benkő, M.; Aoki, K.; Arnberg, N.; Davison, A.J.; Echavarría, M.; Hess, M.; Jones, M.S.; Kaján, G.L.; Kajon, A.E.; Mittal, S.K.; et al. ICTV Virus Taxonomy Profile: Adenoviridae 2022. J. Gen. Virol. 2022, 103, 001721. [Google Scholar] [CrossRef] [PubMed]
- El-Shall, N.A.; El-Hamid, H.S.A.; Elkady, M.F.; Ellakany, H.F.; Elbestawy, A.R.; Gado, A.R.; Geneedy, A.M.; Hasan, M.E.; Jaremko, M.; Selim, S.; et al. Epidemiology, pathology, prevention, and control strategies of inclusion body hepatitis and hepatitis-hydropericardium syndrome in poultry: A comprehensive review. Front. Vet. Sci. 2022, 9, 963199. [Google Scholar] [CrossRef] [PubMed]
- Schachner, A.; Matos, M.; Grafl, B.; Hess, M. Fowl adenovirus-induced diseases and strategies for their control—A review on the current global situation. Avian Pathol. 2018, 47, 111–126. [Google Scholar] [CrossRef] [PubMed]
- Harrach, B.; Tarján, Z.L.; Benkő, M. Adenoviruses across the animal kingdom: A walk in the zoo. FEBS Lett. 2019, 593, 3660–3673. [Google Scholar] [CrossRef]
- Li, C.; Li, H.; Wang, D.; Wang, J.; Wang, Y.; Wang, S.; Li, J.; Liu, P.; Wang, J.; Xu, S.; et al. Characterization of fowl adenoviruses isolated between 2007 and 2014 in China. Vet. Microbiol. 2016, 197, 62–67. [Google Scholar] [CrossRef]
- Chen, L.; Yin, L.; Zhou, Q.; Peng, P.; Du, Y.; Liu, L.; Zhang, Y.; Xue, C.; Cao, Y. Epidemiological investigation of fowl adenovirus infections in poultry in China during 2015-2018. BMC Vet. Res. 2019, 15, 271. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, Q.; Wang, S.; Zhang, F.; Zhao, C.; Chen, Q.; Zhao, R.; Guo, P.; Ju, L.; Li, J.; Hou, G.; et al. Molecular epidemiology analysis of fowl adenovirus detected from apparently healthy birds in eastern China. BMC Vet. Res. 2023, 19, 5. [Google Scholar] [CrossRef]
- Hess, M. Avidenovirus infections. In Diseases of Poultry, 14th ed.; Swayne, D.E., Boulianne, M., Logue, C.M., McDougald, L.R., Nair, V., Suarez, D.L., Wit, S.D., Grimes, T., Johnson, D., Kromm, M., et al., Eds.; Wiley-Blackwell: Ames, IA, USA, 2020; pp. 321–332. [Google Scholar]
- Mohamed, M.H.A.; El-Sabagh, I.M.; Abdelaziz, A.M.; Al-Ali, A.M.; Alramadan, M.; Lebdah, M.A.; Ibrahim, A.M.; Al-Ankari, A.S. Molecular characterization of fowl aviadenoviruses species D and E associated with inclusion body hepatitis in chickens and falcons indicates possible cross-species transmission. Avian Pathol. 2018, 47, 384–390. [Google Scholar] [CrossRef]
- Yang, Q.; Xue, X.; Zhang, Z.; Wu, M.J.; Ji, J.; Wang, W.; Yin, H.; Li, S.; Dai, H.; Duan, B.; et al. Clade 2.3.4.4b H5N8 Subtype Avian Influenza Viruses Were Identified from the Common Crane Wintering in Yunnan Province, China. Viruses 2022, 15, 38. [Google Scholar] [CrossRef]
- Meulemans, G.; Boschmans, M.; Berg, T.P.; Decaesstecker, M. Polymerase chain reaction combined with restriction enzyme analysis for detection and differentiation of fowl adenoviruses. Avian Pathol. 2001, 30, 655–660. [Google Scholar] [CrossRef]
- Mo, K.K.; Lyu, C.F.; Cao, S.S.; Li, X.; Xing, G.; Yan, Y.; Zheng, X.J.; Liao, M.; Zhou, J.Y. Pathogenicity of an FAdV-4 isolate to chickens and its genomic analysis. J. Zhejiang Univ. Sci. B 2019, 20, 740–752. [Google Scholar] [CrossRef]
- Wang, B.; Song, C.; Yang, P.; Song, M.; Zhao, S.; Qiao, Q.; Wang, Z.; Zhao, J. The Role of Hexon Amino Acid 188 Varies in Fowl Adenovirus Serotype 4 Strains with Different Virulence. Microbiol. Spectr. 2022, 10, e0149322. [Google Scholar] [CrossRef] [PubMed]
- Lei, C.; Yang, J.; Hu, J.; Sun, X. On the Calculation of TCID(50) for Quantitation of Virus Infectivity. Virol. Sin. 2021, 36, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Yu, G.; Niu, Y.; Cai, Y.; Liu, S. Airborne Transmission of a Serotype 4 Fowl Adenovirus in Chickens. Viruses 2019, 11, 262. [Google Scholar] [CrossRef] [Green Version]
- Schachner, A.; Marek, A.; Grafl, B.; Hess, M. Detailed molecular analyses of the hexon loop-1 and fibers of fowl aviadenoviruses reveal new insights into the antigenic relationship and confirm that specific genotypes are involved in field outbreaks of inclusion body hepatitis. Vet. Microbiol. 2016, 186, 13–20. [Google Scholar] [CrossRef]
- Kumar, R.; Kumar, V.; Asthana, M.; Shukla, S.K.; Chandra, R. Isolation and identification of a fowl adenovirus from wild Black Kites (Milvus migrans). J. Wildl. Dis. 2010, 46, 272–276. [Google Scholar] [CrossRef]
- Harrach, B.; Megyeri, A.; Papp, T.; Ursu, K.; Boldogh, S.A.; Kaján, G.L. A screening of wild bird samples enhances our knowledge about the biodiversity of avian adenoviruses. Vet. Res. Commun. 2023, 47, 297–303. [Google Scholar] [CrossRef]
- Yang, F.; Zhang, Y.Q. Quantities and distributions of the Black-necked Crane (Grus nigricollis) and other large waterfowls on the Yunnan and Guizhou Plateau. Zoo. Res. 2014, 35, 80–85. [Google Scholar]
- Qian, F.W.; Wu, H.Q.; Gao, L.B.; Zhang, H.G.; Li, F.S.; Zhong, X.Y.; Yang, X.J.; Zheng, G.M. Migration routes and stopover sites on Black-necked Cranes determined by satellite tracking. J. Field Ornithol. 2009, 80, 19–26. [Google Scholar] [CrossRef]
- Li, L.; Luo, L.; Luo, Q.; Zhang, T.; Zhao, K.; Wang, H.; Zhang, R.; Lu, Q.; Pan, Z.; Shao, H.; et al. Genome Sequence of a Fowl Adenovirus Serotype 4 Strain Lethal to Chickens, Isolated from China. Genome Announc. 2016, 4, e00140-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Q.; Wang, J.; Gao, Y.; Cui, H.; Liu, C.; Qi, X.; Zhang, Y.; Wang, Y.; Wang, X. The Natural Large Genomic Deletion Is Unrelated to the Increased Virulence of the Novel Genotype Fowl Adenovirus 4 Recently Emerged in China. Viruses 2018, 10, 494. [Google Scholar] [CrossRef] [Green Version]
- Mo, J. Historical Investigation of Fowl Adenovirus Outbreaks in South Korea from 2007 to 2021: A Comprehensive Review. Viruses 2021, 13, 2256. [Google Scholar] [CrossRef] [PubMed]
- Niczyporuk, J.S.; Kozdruń, W.; Czekaj, H.; Styś-Fijoł, N.; Piekarska, K. Detection of fowl adenovirus D strains in wild birds in Poland by Loop-Mediated Isothermal Amplification (LAMP). BMC Vet. Res. 2020, 16, 58. [Google Scholar] [CrossRef]
- Pan, Q.; Yang, Y.; Shi, Z.; Liu, L.; Gao, Y.; Qi, X.; Liu, C.; Zhang, Y.; Cui, H.; Wang, X. Different Dynamic Distribution in Chickens and Ducks of the Hypervirulent, Novel Genotype Fowl Adenovirus Serotype 4 Recently Emerged in China. Front. Microbiol. 2017, 8, 1005. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Dou, Y.; Zheng, X.; Tang, Y.; Zhang, M.; Zhang, Y.; Wang, Z.; Diao, Y. Hydropericardium Hepatitis Syndrome Emerged in Cherry Valley Ducks in China. Transbound. Emerg. Dis. 2017, 64, 1262–1267. [Google Scholar] [CrossRef]
- Pan, Q.; Liu, L.; Wang, Y.; Zhang, Y.; Qi, X.; Liu, C.; Gao, Y.; Wang, X.; Cui, H. The first whole genome sequence and pathogenicity characterization of a fowl adenovirus 4 isolated from ducks associated with inclusion body hepatitis and hydropericardium syndrome. Avian Pathol. 2017, 46, 571–578. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Wang, Z.; Chen, H.; Niu, X.; Dou, Y.; Yang, J.; Tang, Y.; Diao, Y. Serological and Pathogenic Analyses of Fowl Adenovirus Serotype 4 (FAdV-4) Strain in Muscovy Ducks. Front. Microbiol. 2018, 9, 1163. [Google Scholar] [CrossRef]
- Wei, Z.; Liu, H.; Diao, Y.; Li, X.; Zhang, S.; Gao, B.; Tang, Y.; Hu, J.; Diao, Y. Pathogenicity of fowl adenovirus (FAdV) serotype 4 strain SDJN in Taizhou geese. Avian Pathol. 2019, 48, 477–485. [Google Scholar] [CrossRef]
- Tang, Z.; Liu, M.; Gao, Z.; Li, M.; Cao, J.; Ye, H.; Song, S.; Yan, L. Pathogenicity and virus shedding ability of fowl adenovirus serotype 4 to ducks. Vet. Microbiol. 2022, 264, 109302. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, R.; Tian, K.; Wang, Z.; Yang, X.; Gao, D.; Zhang, Y.; Fu, J.; Wang, H.; Zhao, J. Fiber2 and hexon genes are closely associated with the virulence of the emerging and highly pathogenic fowl adenovirus 4. Emerg. Microbes Infect. 2018, 7, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Q.; Liu, L.; Gao, Y.; Liu, C.; Qi, X.; Zhang, Y.; Wang, Y.; Li, K.; Gao, L.; Wang, X.; et al. Characterization of a hypervirulent fowl adenovirus 4 with the novel genotype newly prevalent in China and establishment of reproduction infection model of hydropericardium syndrome in chickens. Poult. Sci. 2017, 96, 1581–1588. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, X.; Yang, Q.; Wu, M.J.; Zhang, Z.; Song, J.; Wang, W.; Yang, J.; Ji, J.; Zhang, Y.; Dai, H.; et al. Genomic and Pathologic Characterization of the First FAdV-C Serotype 4 Isolate from Black-Necked Crane. Viruses 2023, 15, 1653. https://doi.org/10.3390/v15081653
Xue X, Yang Q, Wu MJ, Zhang Z, Song J, Wang W, Yang J, Ji J, Zhang Y, Dai H, et al. Genomic and Pathologic Characterization of the First FAdV-C Serotype 4 Isolate from Black-Necked Crane. Viruses. 2023; 15(8):1653. https://doi.org/10.3390/v15081653
Chicago/Turabian StyleXue, Xiaoyan, Qinhong Yang, Ming J. Wu, Zhenxing Zhang, Jianling Song, Wei Wang, Jia Yang, Jia Ji, Yongxian Zhang, Hongyang Dai, and et al. 2023. "Genomic and Pathologic Characterization of the First FAdV-C Serotype 4 Isolate from Black-Necked Crane" Viruses 15, no. 8: 1653. https://doi.org/10.3390/v15081653