
Detection and Molecular Characterization of Adenoviruses in Captive and Free-Roaming African Green Monkeys (Chlorocebus sabaeus): Evidence for Possible Recombination and Cross-Species Transmission
 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
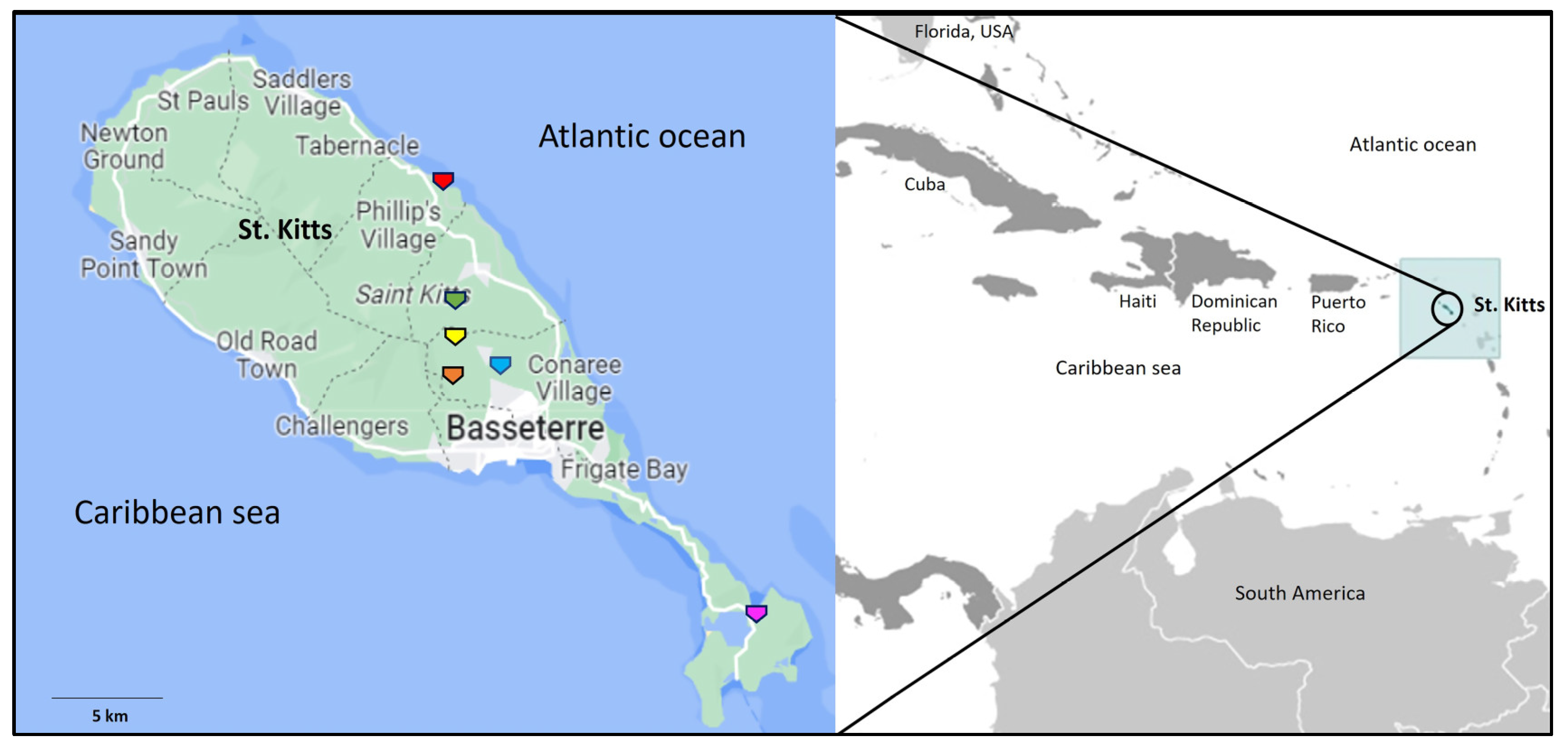
2.1. Sampling
2.2. Amplification of AdV DNA
2.3. Nucleotide Sequencing
2.4. Sequence Analysis
2.5. GenBank Accession Numbers
3. Results
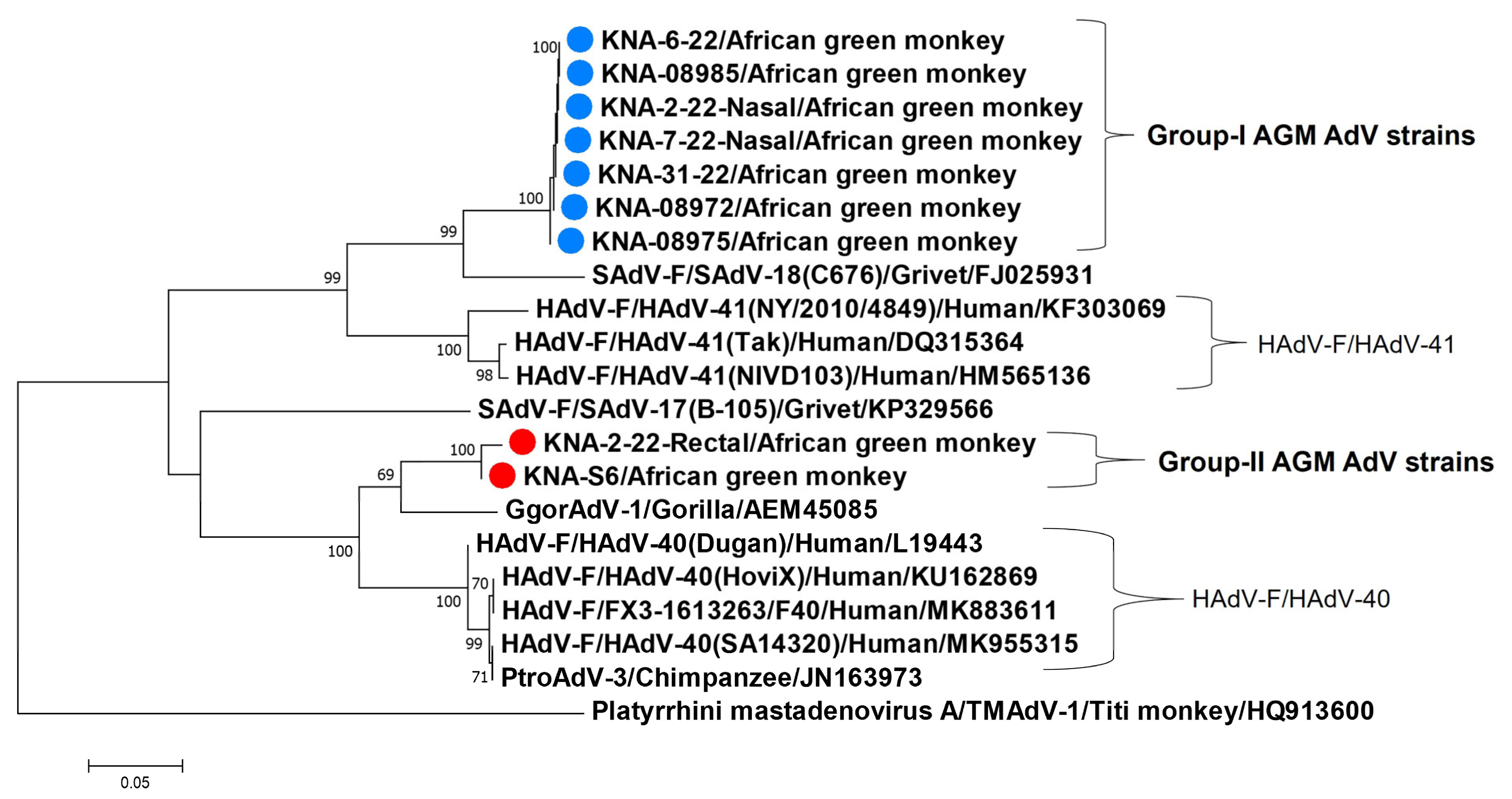
3.1. Detection of AdVs in AGMs
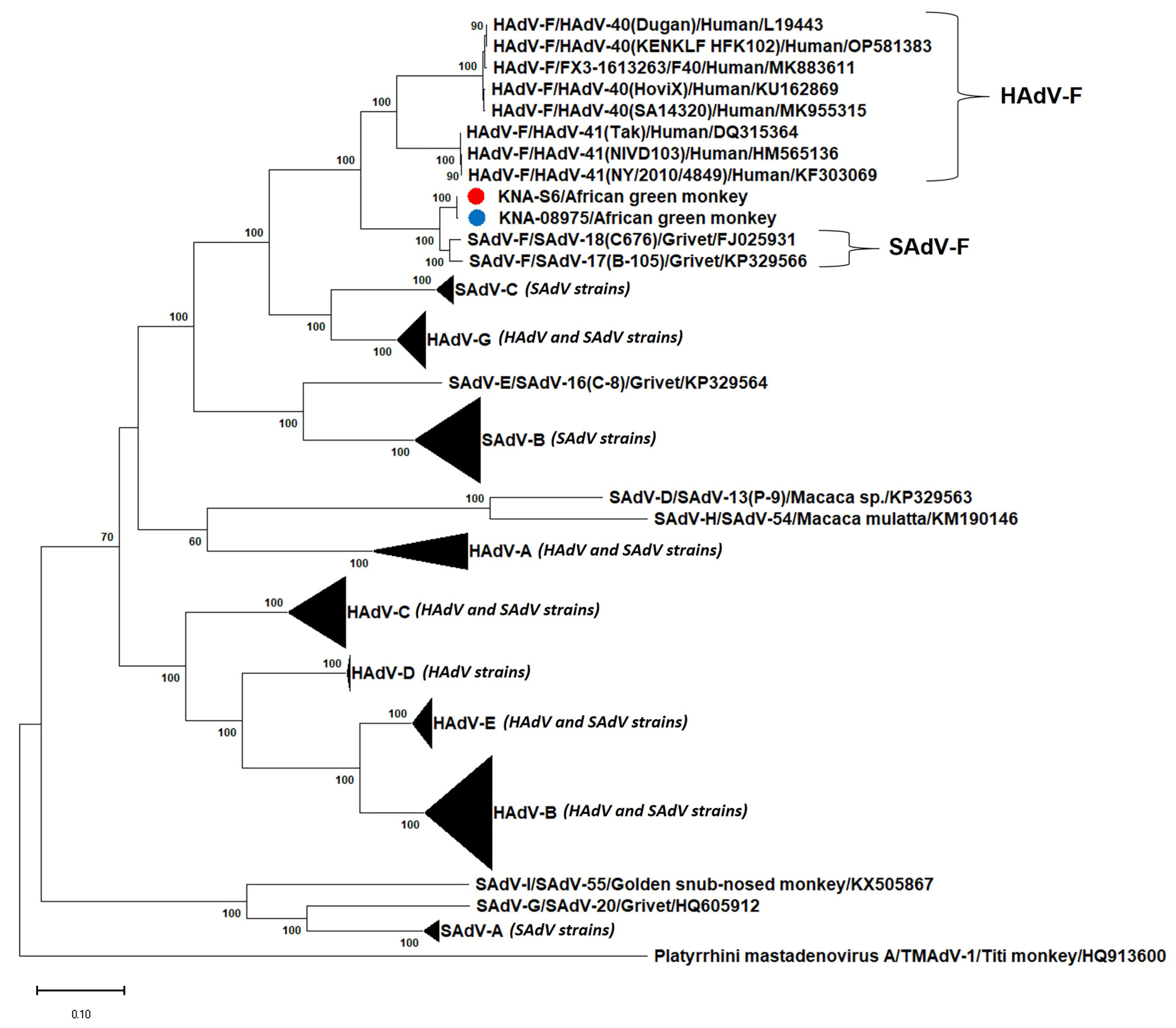
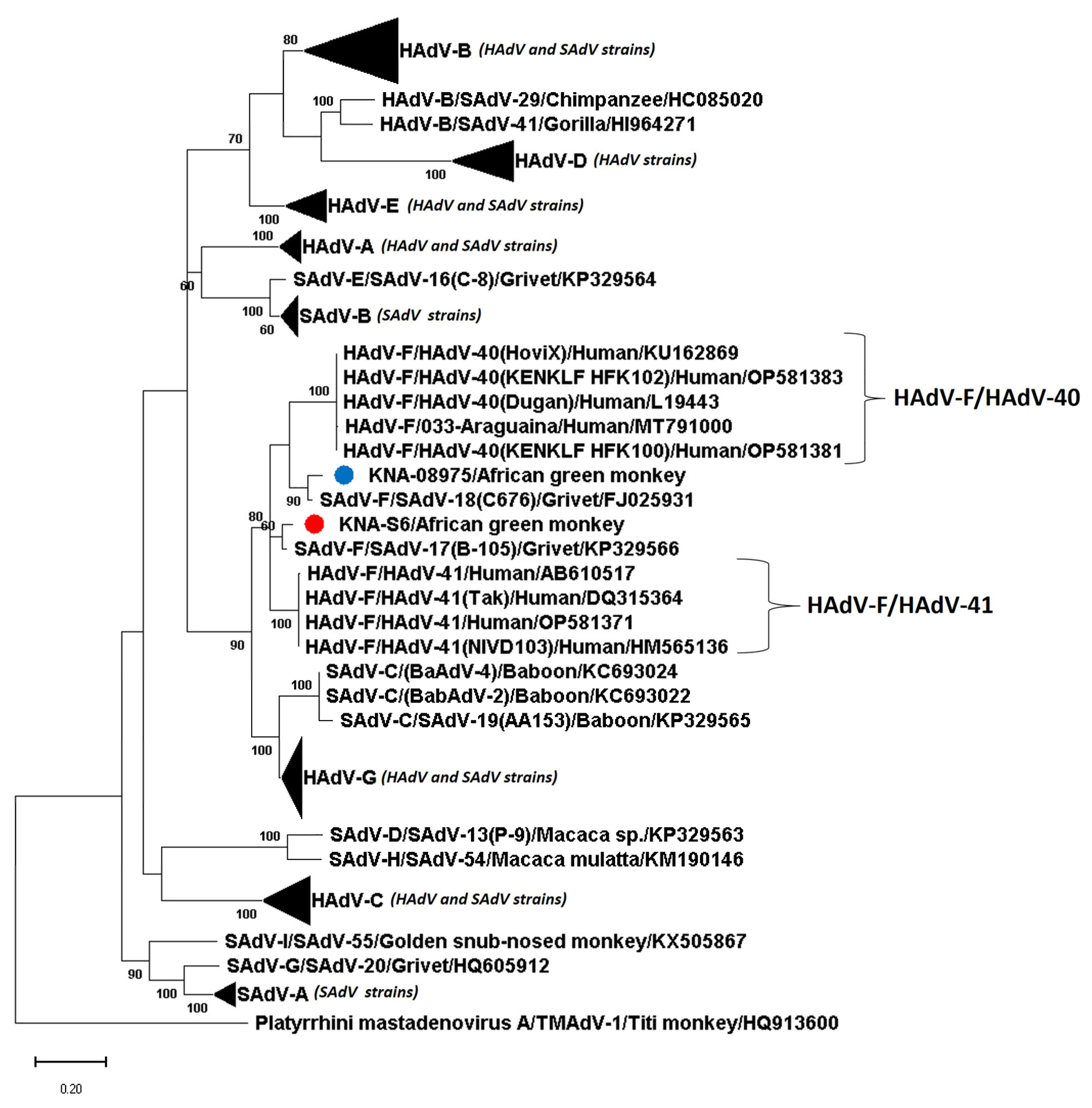
3.2. Analysis of the Partial Pol and Hexon Sequences of AGM AdV Strains
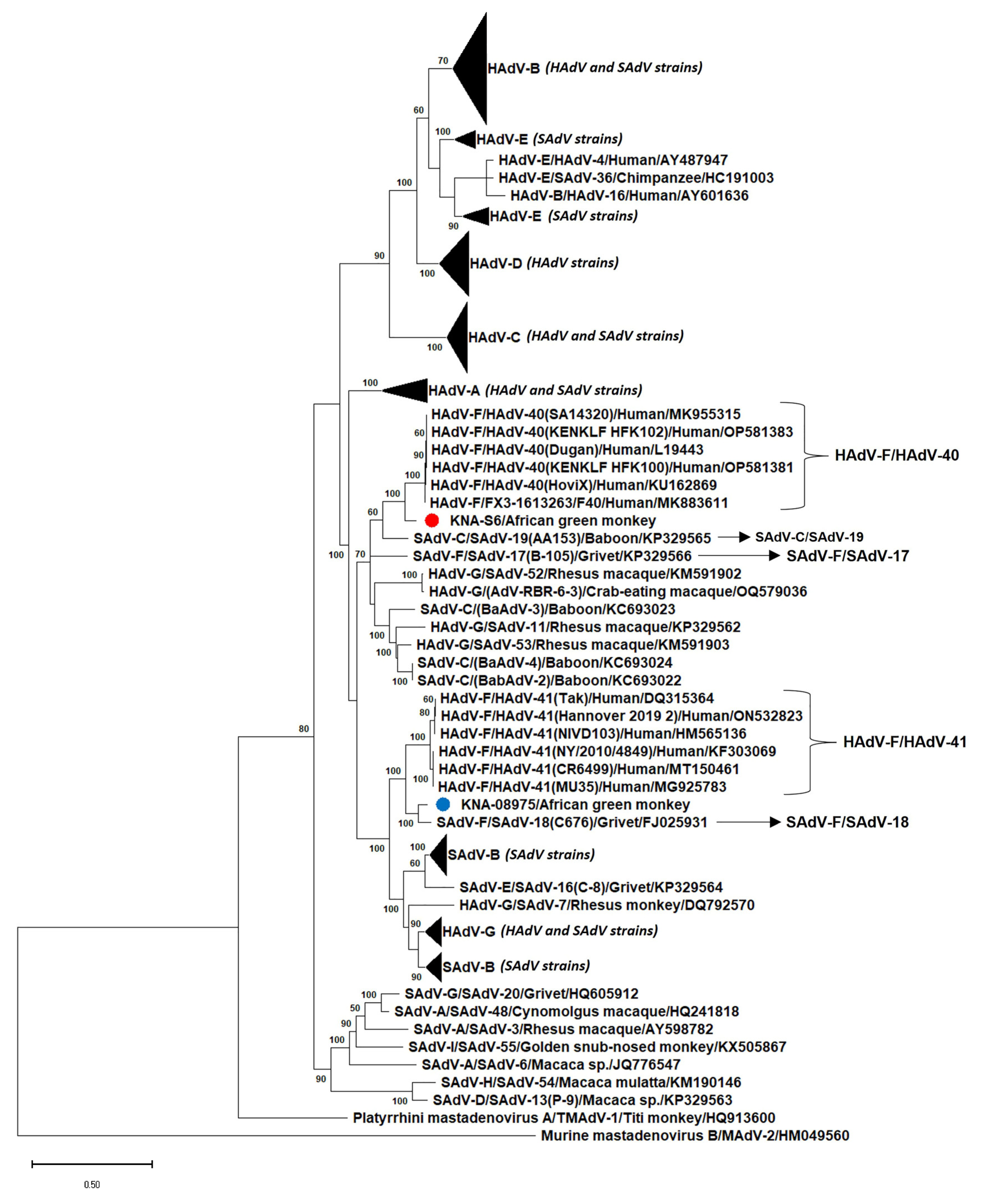
3.3. Analysis of the Nearly Complete Pol, Complete Hexon and Partial Penton Coding Sequences of AGM AdV Strains
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Benkő, M.; Aoki, K.; Arnberg, N.; Davison, A.J.; Echavarría, M.; Hess, M.; Jones, M.S.; Kaján, G.L.; Kajon, A.E.; Mittal, S.K.; et al. ICTV Virus Taxonomy Profile: Adenoviridae 2022. J. Gen. Virol. 2022, 103, 001721. [Google Scholar] [CrossRef]
- Harrach, B.; Benkő, M. Adenoviruses (Adenoviridae). In Encyclopedia of Virology; Bamford, D.H., Zuckerman, M., Eds.; Academic Press: Amsterdam, The Netherlands, 2021; pp. 3–16. [Google Scholar] [CrossRef]
- Harrach, B.; Tarján, Z.L.; Benkő, M. Adenoviruses across the animal kingdom: A walk in the zoo. FEBS Lett. 2019, 593, 3660–3673. [Google Scholar] [CrossRef]
- Machlachlan, J.; Dubovi, E. Chapter 10—Adenoviridae. In Fenner’s Veterinary Virology, 5th ed.; Academic Press: Amsterdam, The Netherlands, 2016; pp. 217–227. [Google Scholar] [CrossRef]
- Pellegrinelli, L.; Uceda Renteria, S.C.; Ceriotti, F.; Ammoni, E.; Galli, C.; Seiti, A.; Castiglioni, S.; Cereda, D.; Binda, S.; Pariani, E. Wastewater Surveillance Captured an Increase in Adenovirus Circulation in Milan (Italy) during the First Quarter of 2022. Viruses 2022, 14, 2351. [Google Scholar] [CrossRef]
- Hoeben, R.C.; Uil, T.G. Adenovirus DNA replication. Cold Spring Harb. Perspect. Biol. 2013, 5, a013003. [Google Scholar] [CrossRef] [Green Version]
- Gallardo, J.; Pérez-Illana, M.; Martín-González, N.; San Martín, C. Adenovirus Structure: What Is New? Int. J. Mol. Sci. 2021, 22, 5240. [Google Scholar] [CrossRef] [PubMed]
- Bányai, K.; Esona, M.D.; Liu, A.; Wang, Y.; Tu, X.; Jiang, B. Molecular detection of novel adenoviruses in fecal specimens of captive monkeys with diarrhea in China. Vet. Microbiol. 2010, 142, 416–419. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.C.; Yagi, S.; Kelly, K.R.; Mendoza, S.P.; Tarara, R.P.; Canfield, D.R.; Maninger, N.; Rosenthal, A.; Spinner, A.; Bales, K.L.; et al. Cross-species transmission of a novel adenovirus associated with a fulminant pneumonia outbreak in a new world monkey colony. PLoS Pathog. 2011, 7, e1002155. [Google Scholar] [CrossRef]
- Kim, C.S.; Sueltenfuss, E.S.; Kalter, S.S. Isolation and characterization of simian adenoviruses isolated in association with an outbreak of pneumoenteritis in vervet monkeys (Cercopithecus aethiops). J. Infect. Dis. 1967, 117, 292–300. [Google Scholar] [CrossRef]
- Vasileva, V.A.; Ivanov, M.T.; Rumel, N.B.; D’Yachenko, A.G.; Kakubava, V.V.; Danelyan, G.A. Isolation and biological characterization of an adenovirus of rhesus macaques. Acta Biol. Medica Ger. 1978, 37, 1281–1287. [Google Scholar]
- Zöller, M.; Mätz-Rensing, K.; Kaup, F.J. Adenoviral hepatitis in a SIV-infected rhesus monkey (Macaca mulatta). J. Med. Primatol. 2008, 37, 184–187. [Google Scholar] [CrossRef]
- Chandler, F.W.; McClure, H.M. Adenoviral pancreatitis in rhesus monkeys: Current knowledge. Vet. Pathol. Suppl. 1982, 7, 171–180. [Google Scholar] [CrossRef] [Green Version]
- Kaján, G.L.; Doszpoly, A.; Tarján, Z.L.; Vidovszky, M.Z.; Papp, T. Virus–Host Coevolution with a Focus on Animal and Human DNA Viruses. J. Mol. Evol. 2020, 88, 41–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.; Ismail, A.M.; Dehghan, S.; Rajaiya, J.; Allard, M.W.; Lim, H.C.; Dyer, D.W.; Chodosh, J.; Seto, D. Genomics-based re-examination of the taxonomy and phylogeny of human and simian Mastadenoviruses: An evolving whole genomes approach, revealing putative zoonosis, anthroponosis, and amphizoonosis. Cladistics 2020, 36, 358–373. [Google Scholar] [CrossRef] [PubMed]
- Podgorski, I.I.; Pantó, L.; Papp, T.; Harrach, B.; Benkö, M. Genome analysis of four Old World monkey adenoviruses supports the proposed species classification of primate adenoviruses and reveals signs of possible homologous recombination. J. Gen. Virol. 2016, 97, 1604–1614. [Google Scholar] [CrossRef] [PubMed]
- Pantó, L.; Podgorski, I.I.; Jánoska, M.; Márkó, O.; Harrach, B. Taxonomy proposal for Old World monkey adenoviruses: Characterisation of several non-human, non-ape primate adenovirus lineages. Arch. Virol. 2015, 160, 3165–3177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podgorski, I.I.; Pantó, L.; Földes, K.; de Winter, I.; Jánoska, M.; Sós, E.; Chenet, B.; Harrach, B.; Benkő, M. Adenoviruses of the most ancient primate lineages support the theory on virus-host co-evolution. Acta Vet. Hung. 2018, 66, 474–487. [Google Scholar] [CrossRef] [Green Version]
- Wevers, D.; Metzger, S.; Babweteera, F.; Bieberbach, M.; Boesch, C.; Cameron, K.; Couacy-Hymann, E.; Cranfield, M.; Gray, M.; Harris, L.A.; et al. Novel adenoviruses in free-roaming primates: A high level of genetic diversity and evidence of zoonotic transmissions. J. Virol. 2011, 85, 10774–10784. [Google Scholar] [CrossRef] [Green Version]
- Morris, S.J.; Sebastian, S.; Spencer, A.J.; Gilbert, S.C. Simian adenoviruses as vaccine vectors. Future Virol. 2016, 11, 649–659. [Google Scholar] [CrossRef] [Green Version]
- Bots, S.T.F.; Hoeben, R.C. Non-Human Primate-Derived Adenoviruses for Future Use as Oncolytic Agents? Int. J. Mol. Sci. 2020, 21, 4821. [Google Scholar] [CrossRef]
- Borkenhagen, L.K.; Fieldhouse, J.K.; Seto, D.; Gray, G.C. Are adenoviruses zoonotic? A systematic review of the evidence. Emerg. Microbes Infect. 2019, 8, 1679–1687. [Google Scholar] [CrossRef] [Green Version]
- Medkour, H.; Amona, I.; Akiana, J.; Davoust, B.; Bitam, I.; Levasseur, A.; Tall, M.L.; Diatta, G.; Sokhna, C.; Hernandez-Aguilar, R.A.; et al. Adenovirus Infections in African Humans and Free-roaming Non-Human Primates: Great Diversity and Cross-Species Transmission. Viruses 2020, 12, 657. [Google Scholar] [CrossRef]
- Bots, S.T.F.; Kemp, V.; Dautzenberg, I.J.C.; Hoeben, R.C. Genome Analyses of Ten New Ape Adenoviruses with Similarity to Human Mastadenovirus C. Int. J. Mol. Sci. 2022, 23, 9832. [Google Scholar] [CrossRef]
- Kosoltanapiwat, N.; Tongshoob, J.; Ampawong, S.; Reamtong, O.; Prasittichai, L.; Yindee, M.; Tongthainan, D.; Tulayakul, P.; Boonnak, K. Simian adenoviruses: Molecular and serological survey in monkeys and humans in Thailand. One Health 2022, 15, 100434. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, C.A.; Hervé-Claude, L.P.; Cruz-Martinez, L.; Craig, S. Understanding Community Perceptions of the St. Kitts’ “Monkey Problem” by Adapting Harm Reduction Concepts and Methods. Front. Ecol. Evol. 2022, 10, 904797. Available online: https://www.frontiersin.org/articles/10.3389/fevo.2022.904797 (accessed on 20 May 2023). [CrossRef]
- Dore, K. Vervets in the Caribbean. In The International Encyclopedia of Primatology; Fuentes, A., Bezanson, B., Campbell, C.J., DiFiore, A.F., Elton, S., Estrada, A., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2017; pp. 1–2. [Google Scholar] [CrossRef]
- Dore, K.M. Ethnoprimatology without conservation: The political ecology of farmer–green monkey (Chlorocebus sabaeus) relations in St. Kitts, West Indies. Int. J. Primatol. 2018, 39, 918–944. [Google Scholar] [CrossRef]
- Dore, K.M.; Gallagher, C.A.; Mill, A.C. Telemetry-Based Assessment of Home Range to Estimate the Abundance of Invasive Green Monkeys on St. Kitts. Caribb. J. Sci. 2023, 53, 1–17. [Google Scholar] [CrossRef]
- Navarro, R.; Aung, M.S.; Cruz, K.; Ketzis, J.; Gallagher, C.A.; Beierschmitt, A.; Malik, Y.S.; Kobayashi, N.; Ghosh, S. Whole genome analysis provides evidence for porcine-to-simian interspecies transmission of rotavirus-A. Infect. Genet. Evol. 2017, 49, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Richins, T.; Sapp, S.G.H.; Ketzis, J.K.; Willingham, A.L.; Mukaratirwa, S.; Qvarnstrom, Y.; Barratt, J.L.N. Genetic characterization of Strongyloides fuelleborni infecting free-roaming African vervets (Chlorocebus aethiops sabaeus) on the Caribbean island of St. Kitts. Int. J. Parasitol. Parasites Wildl. 2023, 20, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.; Walkush, J.; Shim, D.; Cruz, K.; Ketzis, J. Molecular species identification of Trichuris trichiura in African green monkey on St. Kitts, West Indies. Vet. Parasitol. Reg. Stud. Rep. 2018, 11, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Hoefer, A.; Boyen, F.; Beierschmitt, A.; Moodley, A.; Roberts, M.C.; Butaye, P. Methicillin-Resistant and Methicillin-Susceptible Staphylococcus from Vervet Monkeys (Chlorocebus sabaeus) in Saint Kitts. Antibiotics 2021, 10, 290. [Google Scholar] [CrossRef]
- Rajeev, S.; Conan, A.; Pratt, N.; Beierschmitt, A.; Palmour, R. High Leptospira seroprevalence in captive and free-roaming-caught vervet monkeys (Chlorocebus sabeus) on the Caribbean island of Saint Kitts. J. Vet. Diagn. Investig. 2017, 29, 930–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallagher, C.; Beierschmitt, A.; Cruz, K.; Choo, J.; Ketzis, J. Should monkeys wash their hands and feet: A pilot-study on sources of zoonotic parasite exposure. One Health 2019, 7, 100088. [Google Scholar] [CrossRef] [PubMed]
- Goulet, N.R.; Schriver, P.W.; Liu, O.C.; Beiler, J.M. Isolation and propagation of adenoviruses in African green monkey kidney cultures. J. Infect. Dis. 1965, 115, 429–435. [Google Scholar] [CrossRef]
- Lawrence, M.S.; Foellmer, H.G.; Elsworth, J.D.; Kim, J.H.; Leranth, C.; Kozlowski, D.A.; Bothwell, A.L.; Davidson, B.L.; Bohn, M.C.; Redmond, D.E. Inflammatory responses and their impact on beta-galactosidase transgene expression following adenovirus vector delivery to the primate caudate nucleus. Gene Ther. 1999, 6, 1368–1379. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.; Vandenberghe, L.H.; Kryazhimskiy, S.; Grant, R.; Calcedo, R.; Yuan, X.; Keough, M.; Sandhu, A.; Wang, Q.; Medina-Jaszek, C.A.; et al. Isolation and characterization of adenoviruses persistently shed from the gastrointestinal tract of non-human primates. PLoS Pathog. 2009, 5, e1000503. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and Analysis of Recombination Patterns in Virus Genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef] [Green Version]
- Kleymann, A.; Soto, E.; Illanes, O.; Malik, Y.S.; Fuentealba, C.; Ghosh, S. High rates of detection and complete genomic analysis of porcine circovirus 2 (PCV2) in the Lesser Antilles island of St. Kitts: Identification of PCV2b-PCV2d recombinants. Transbound. Emerg. Dis. 2020, 67, 2282–2289. [Google Scholar] [CrossRef]
- Wellehan, J.F.X.; Johnson, A.J.; Harrach, B.; Benkö, M.; Pessier, A.P.; Johnson, C.M.; Garner, M.M.; Childress, A.; Jacobson, E.R. Detection and Analysis of Six Lizard Adenoviruses by Consensus Primer PCR Provides Further Evidence of a Reptilian Origin for the Atadenoviruses. J. Virol. 2004, 78, 13366–13369. [Google Scholar] [CrossRef] [Green Version]
- Gainor, K.; Becker, A.A.M.J.; Malik, Y.S.; Ghosh, S. First Report on Detection and Molecular Characterization of Adenoviruses in the Small Indian Mongoose (Urva auropunctata). Viruses 2021, 13, 2194. [Google Scholar] [CrossRef]
- Gainor, K.; Fortuna, Y.C.; Alakkaparambil, A.S.; González, W.; Malik, Y.S.; Ghosh, S. High Rates of Detection and Molecular Characterization of Porcine Adenovirus Serotype 5 (Porcine mastadenovirus C) from Diarrheic Pigs. Pathogens 2022, 11, 1210. [Google Scholar] [CrossRef]
- Roy, S.; Sandhu, A.; Medina, A.; Clawson, D.S.; Wilson, J.M. Adenoviruses in fecal samples from asymptomatic rhesus macaques, United States. Emerg. Infect. Dis. 2012, 18, 1081–1088. [Google Scholar] [CrossRef] [PubMed]
- Podgorski, I.I.; Harrach, B.; Benkő, M.; Papp, T. Characterization of monkey adenoviruses with three fiber genes. Infect. Genet. Evol. 2023, 108, 105403. [Google Scholar] [CrossRef]
- Kalter, S.S. Enteric viruses of nonhuman primates. Vet. Pathol. Suppl. 1982, 7, 33–43. [Google Scholar] [CrossRef]
- Hoffert, W.R.; Bates, M.E.; Cheever, F.S. Study of enteric viruses of simian origin. Am. J. Epidemiol. 1958, 68, 15–30. [Google Scholar] [CrossRef]
- Heberling, R.L.; Cheever, F.S. Enteric viruses of monkeys. Ann. N. Y. Acad. Sci. 1960, 85, 942–950. [Google Scholar] [CrossRef] [PubMed]
- Lange, C.E.; Niama, F.R.; Cameron, K.; Olson, S.H.; Aime Nina, R.; Ondzie, A.; Bounga, G.; Smith, B.R.; Pante, J.; Reed, P.; et al. First evidence of a new simian adenovirus clustering with Human mastadenovirus F viruses. Virol. J. 2019, 16, 147. [Google Scholar] [CrossRef]
- Götting, J.; Cordes, A.K.; Steinbrück, L.; Heim, A. Molecular phylogeny of human adenovirus type 41 lineages. Virus. Evol. 2022, 8, veac098. [Google Scholar] [CrossRef]
- Saint-Pierre, C.G.; Conei Valencia, D.; Lizama, L.; Vargas Zuñiga, D.; Avendaño Carvajal, L.F.; Ampuero Llanos, S. An Old Acquaintance: Could Adenoviruses Be Our Next Pandemic Threat? Viruses 2023, 15, 330. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AGM Number | Age, Gender | Adenovirus Positive Sample | Date of Sample Collection | Sampling/Trapping Location in St. Kitts |
|---|---|---|---|---|
| KNA-S2 | Juvenile, male | Feces | 12 July 2014 | Southeast Peninsula 1 |
| KNA-S6 | Adult, male | Feces | 15 November 2014 | Southeast Peninsula 1 |
| KNA-08972 | Not available | Feces | 9 September 2015 | Behavioral Science Foundation 2 |
| KNA-08973 | Not available | Feces | 2 October 2015 | Behavioral Science Foundation 2 |
| KNA-08975 | Juvenile, male | Feces | 19 October 2015 | Behavioral Science Foundation 2 |
| KNA-08977 | Adult, male | Feces | 19 October 2015 | Behavioral Science Foundation 2 |
| KNA-08980 | Not available | Feces | 9 October 2015 | Behavioral Science Foundation 2 |
| KNA-08982 | Not available | Feces | 6 October 2015 | Behavioral Science Foundation 2 |
| KNA-08984 | Not available | Feces | 9 October 2015 | Behavioral Science Foundation 2 |
| KNA-08985 | Adult, female | Feces | 15 October 2015 | Behavioral Science Foundation 2 |
| KNA-08994 | Adult, female | Feces | 15 October 2015 | Behavioral Science Foundation 2 |
| KNA-08995 | Not available | Feces | 27 October 2015 | Behavioral Science Foundation 2 |
| KNA-1-22 | Adult, male | Nasal swab | 24 September 2022 | Monkey hill 1 |
| KNA-2-22 | Juvenile, male | Nasal and rectal swab | 24 September 2022 | Monkey hill 1 |
| KNA-3-22 | Adult, female | Rectal swab | 24 September 2022 | Monkey hill 1 |
| KNA-5-22 | Juvenile, male | Nasal swab | 24 September 2022 | Monkey hill 1 |
| KNA-6-22 | Adult, female | Rectal swab | 24 September 2022 | Monkey hill 1 |
| KNA-7-22 | Juvenile, male | Nasal and rectal swab | 24 September 2022 | Monkey hill 1 |
| KNA-9-22 | Adult, male | Nasal and rectal swab | 24 September 2022 | Monkey hill 1 |
| KNA-11-22 | Adult, female | Nasal swab | 15 December 2022 | Monkey hill 1 |
| KNA-13-22 | Juvenile, female | Rectal swab | 15 December 2022 | Monkey hill 1 |
| KNA-14-22 | Adult, male | Rectal swab | 15 December 2022 | Monkey hill 1 |
| KNA-15-22 | Juvenile, male | Nasal swab | 15 December2022 | Monkey hill 1 |
| KNA-16-22 | Adult, male | Nasal swab | 15 December2022 | Monkey hill 1 |
| KNA-22-22 | Adult, female | Rectal swab | 19 December2022 | Cedar Grove 3 |
| KNA-25-22 | Adult, female | Nasal swab | 19 December2022 | Cedar Grove 3 |
| KNA-31-22 | Adult, male | Rectal swab | 20 December2022 | Cedar Grove 3 |
| KNA-38-22 | Adult, male | Rectal swab | 29 December2022 | Green hill 3 |
| Putative Adenovirus Protein | Deduced aa Identities (%) | |
|---|---|---|
| KNA-S6 | KNA-08975 | |
| DNA-dependent DNA polymerase (Pol) 1 |
|
|
| Hexon 2 |
|
|
| Penton 3 |
|
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mancuso, D.M.; Gainor, K.; Dore, K.M.; Gallagher, C.A.; Cruz, K.; Beierschmitt, A.; Malik, Y.S.; Ghosh, S. Detection and Molecular Characterization of Adenoviruses in Captive and Free-Roaming African Green Monkeys (Chlorocebus sabaeus): Evidence for Possible Recombination and Cross-Species Transmission. Viruses 2023, 15, 1605. https://doi.org/10.3390/v15071605
Mancuso DM, Gainor K, Dore KM, Gallagher CA, Cruz K, Beierschmitt A, Malik YS, Ghosh S. Detection and Molecular Characterization of Adenoviruses in Captive and Free-Roaming African Green Monkeys (Chlorocebus sabaeus): Evidence for Possible Recombination and Cross-Species Transmission. Viruses. 2023; 15(7):1605. https://doi.org/10.3390/v15071605
Chicago/Turabian StyleMancuso, Diana M., Kerry Gainor, Kerry M. Dore, Christa A. Gallagher, Katalina Cruz, Amy Beierschmitt, Yashpal S. Malik, and Souvik Ghosh. 2023. "Detection and Molecular Characterization of Adenoviruses in Captive and Free-Roaming African Green Monkeys (Chlorocebus sabaeus): Evidence for Possible Recombination and Cross-Species Transmission" Viruses 15, no. 7: 1605. https://doi.org/10.3390/v15071605