
Antigenic Characterization of Infectious Bronchitis Virus in the South China during 2021–2022
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus Isolation and Identification
2.2. Cloning and Sequencing of S1
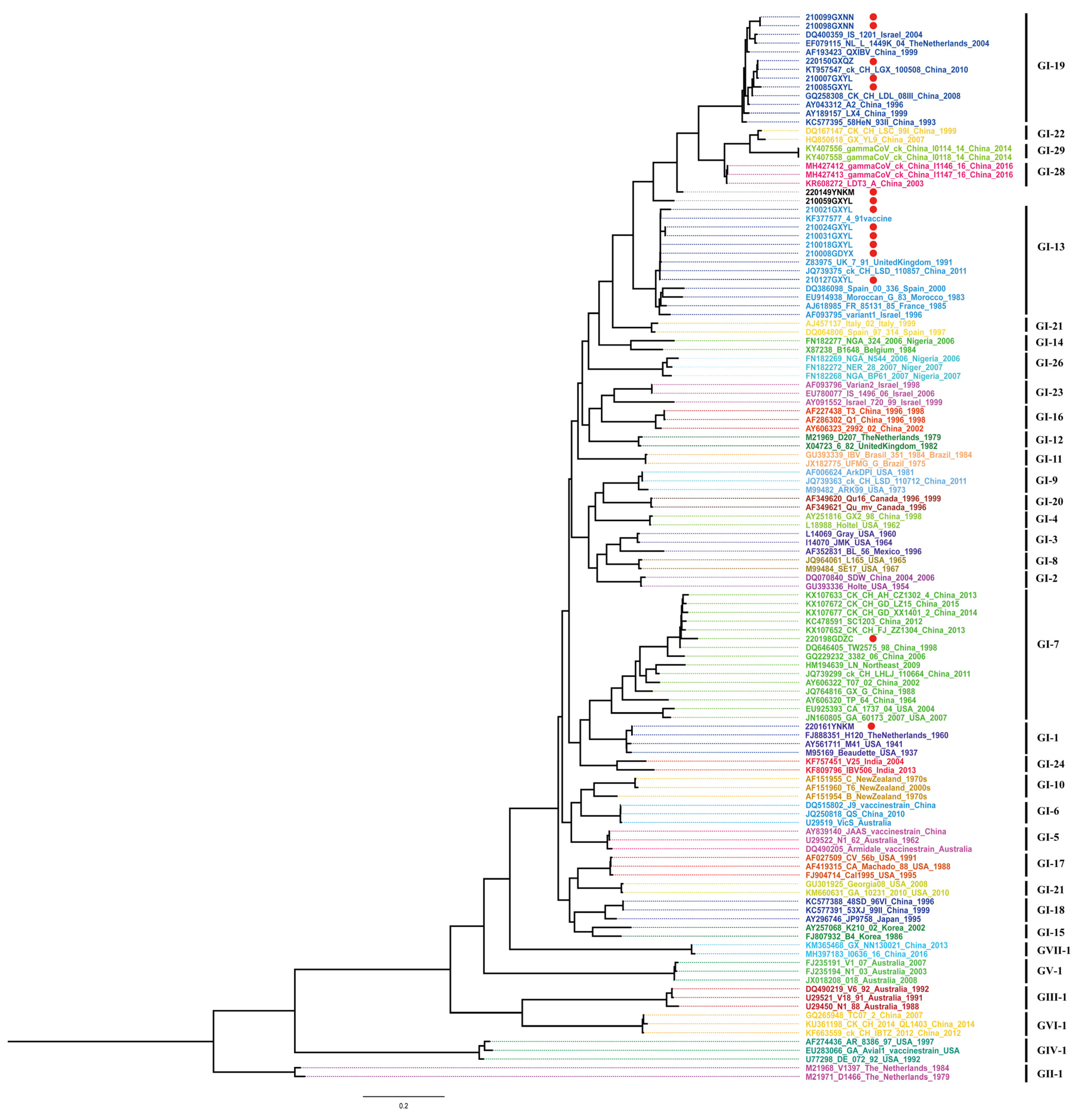
2.3. Phylogenetic Analysis
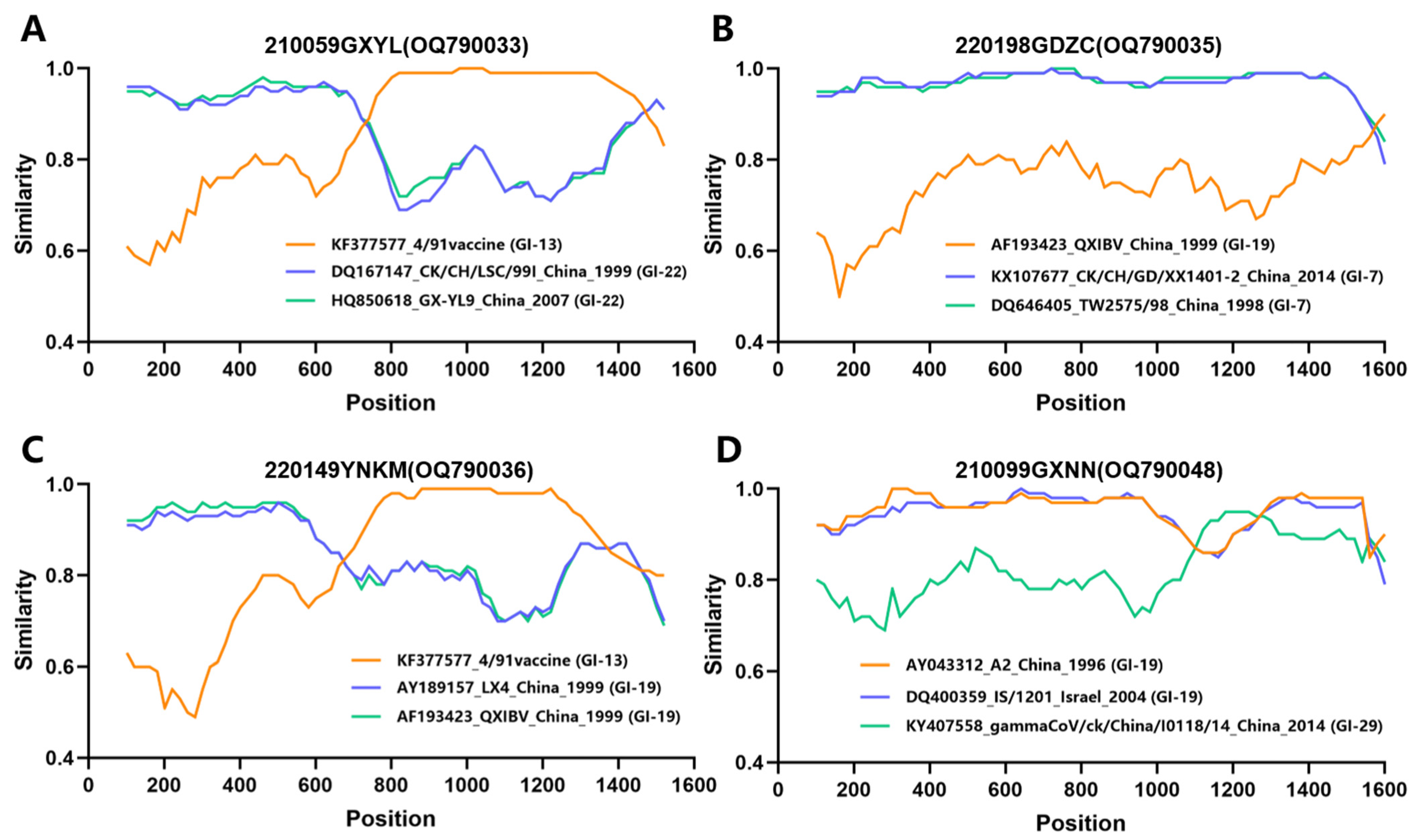
2.4. Recombination Analysis
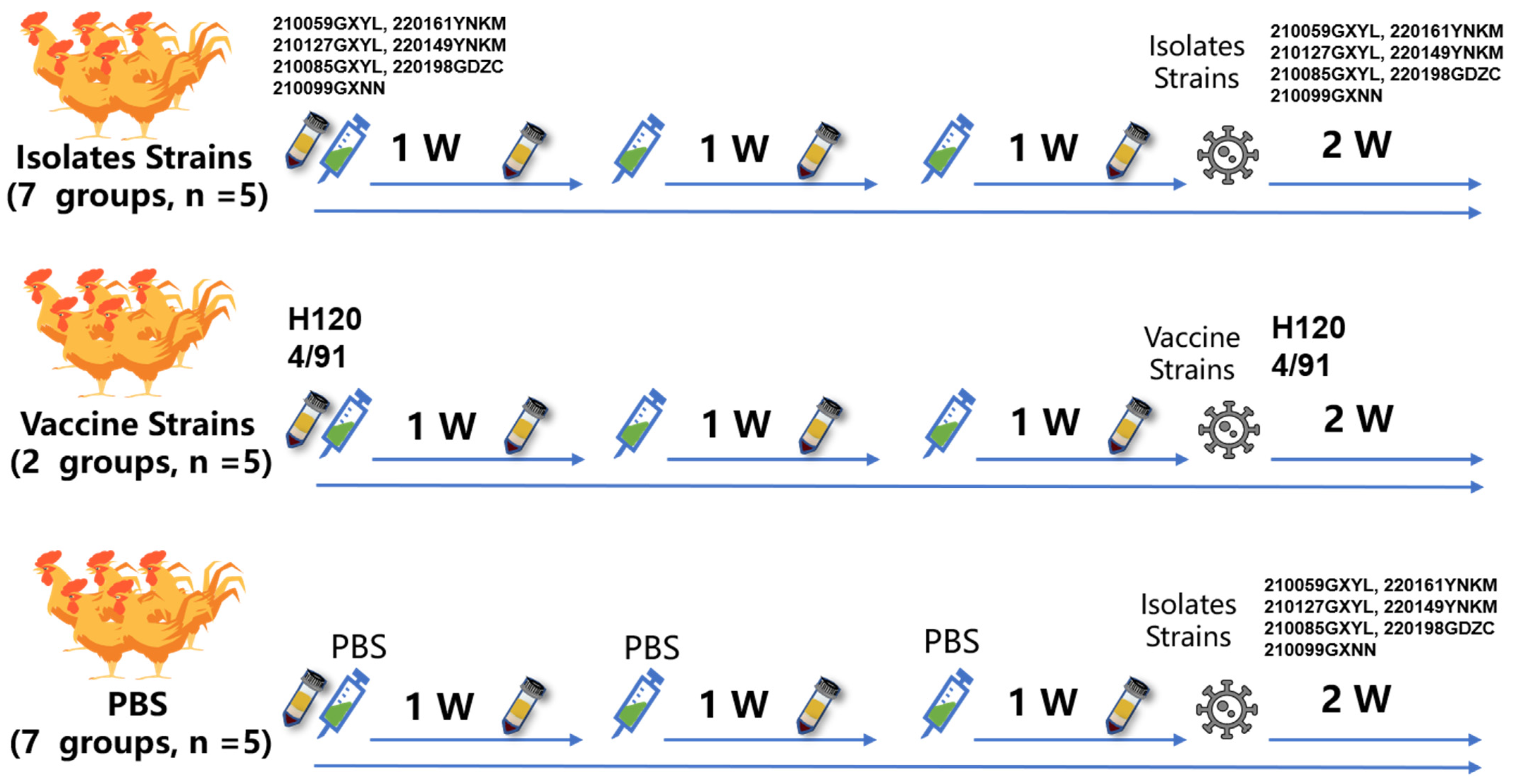
2.5. Inactivated IBV Vaccine and Testing
2.6. Antigenic Correlation Coefficient
3. Results
3.1. Epidemic Information and S1 Sequences of Clinical Samples
3.2. Evolutionary Genetic and Recombination Analysis of S1
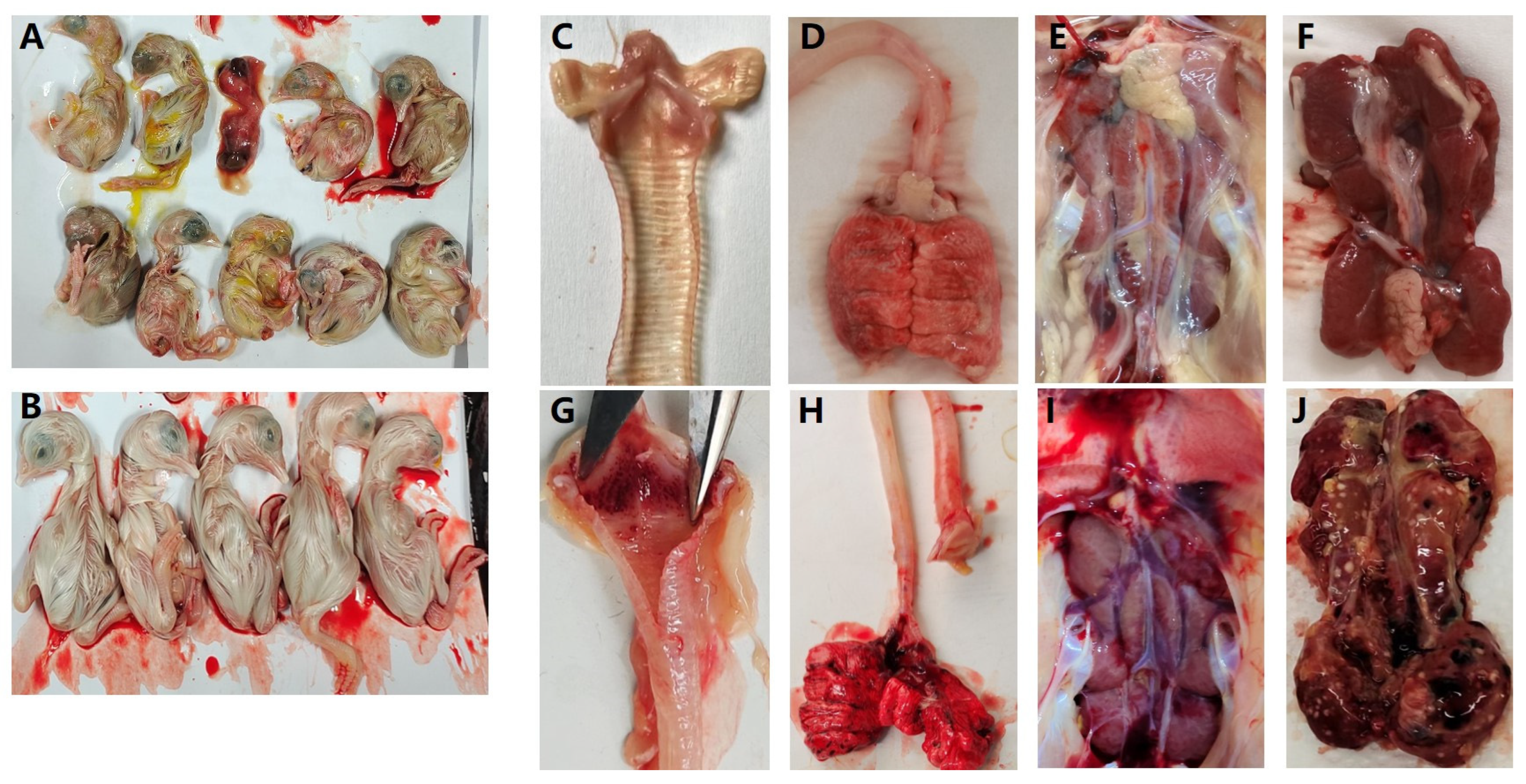
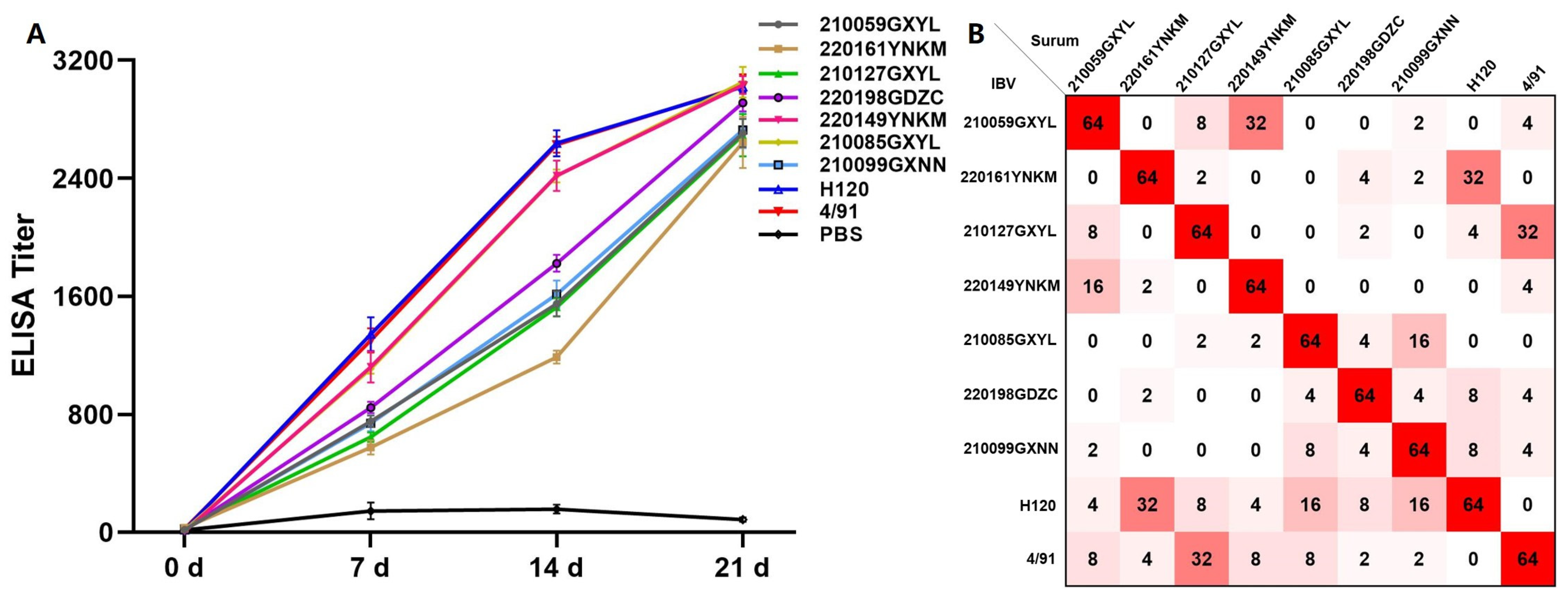
3.3. Antigenic Characterization of Isolate Strains
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sjaak de Wit, J.J.; Cook, J.K.; van der Heijden, H.M. Infectious bronchitis virus variants: A review of the history, current situation and control measures. Avian Pathol. 2011, 40, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Cook, J.K.A.; Jackwood, M.; Jones, R.C. The long view: 40 years of infectious bronchitis research. Avian Pathol. 2012, 41, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Bande, F.; Arshad, S.S.; Bejo, M.H.; Moeini, H.; Omar, A.R. Progress and Challenges toward the Development of Vaccines against Avian Infectious Bronchitis. J. Immunol. Res. 2015, 2015, 424860. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, D. Coronaviruses in poultry and other birds. Avian Pathol. 2005, 34, 439–448. [Google Scholar] [CrossRef]
- Cavanagh, D. Coronavirus avian infectious bronchitis virus. Vet. Res. 2007, 38, 281–297. [Google Scholar] [CrossRef]
- Shang, J.; Zheng, Y.; Yang, Y.; Liu, C.; Geng, Q.; Luo, C.; Zhang, W.; Li, F. Cryo-EM structure of infectious bronchitis coronavirus spike protein reveals structural and functional evolution of coronavirus spike proteins. PLoS Pathog. 2018, 14, e1007009. [Google Scholar] [CrossRef]
- Ali, A.; Ojkic, D.; Elshafiee, E.A.; Shany, S.; El-Safty, M.M.; Shalaby, A.A.; Abdul-Careem, M.F. Genotyping and In Silico Analysis of Delmarva (DMV/1639) Infectious Bronchitis Virus (IBV) Spike 1 (S1) Glycoprotein. Genes 2022, 13, 1617. [Google Scholar] [CrossRef]
- Kabinger, F.; Stiller, C.; Schmitzova, J.; Dienemann, C.; Kokic, G.; Hillen, H.S.; Hobartner, C.; Cramer, P. Mechanism of molnupiravir-induced SARS-CoV-2 mutagenesis. Nat. Struct. Mol. Biol. 2021, 28, 740–746. [Google Scholar] [CrossRef]
- Xu, L.; Han, Z.; Jiang, L.; Sun, J.; Zhao, Y.; Liu, S. Genetic diversity of avian infectious bronchitis virus in China in recent years. Infect. Genet. Evol. 2018, 66, 82–94. [Google Scholar] [CrossRef]
- Xia, X. Domains and Functions of Spike Protein in SARS-CoV-2 in the Context of Vaccine Design. Viruses 2021, 13, 109. [Google Scholar] [CrossRef]
- Valastro, V.; Holmes, E.C.; Britton, P.; Fusaro, A.; Jackwood, M.W.; Cattoli, G.; Monne, I. S1 gene-based phylogeny of infectious bronchitis virus: An attempt to harmonize virus classification. Infect. Genet. Evol. 2016, 39, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Xu, L.; Ren, M.; Shen, J.; Han, Z.; Sun, J.; Zhao, Y.; Liu, S. Novel genotype of infectious bronchitis virus isolated in China. Vet. Microbiol. 2019, 230, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Feng, K.; Xue, Y.; Wang, F.; Chen, F.; Shu, D.; Xie, Q. Analysis of S1 gene of avian infectious bronchitis virus isolated in southern China during 2011–2012. Virus Genes 2014, 49, 292–303. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, H.; Zhao, J.; Guo-Zhong, Z.; Jin, J.-H.; Zhang, G.-Z. Evolution of infectious bronchitis virus in China over the past two decades. J. Gen. Virol. 2016, 97, 1566–1574. [Google Scholar] [CrossRef]
- Lian, J.; Wang, Z.; Xu, Z.; Chen, T.; Shao, G.; Zhang, X.; Qin, J.; Xie, Q.; Lin, W. Distribution and molecular characterization of avian infectious bronchitis virus in southern China. Poult. Sci. 2021, 100, 101169. [Google Scholar] [CrossRef]
- Ren, M.; Sheng, J.; Ma, T.; Xu, L.; Han, Z.; Li, H.; Zhao, Y.; Sun, J.; Liu, S. Molecular and biological characteristics of the infectious bronchitis virus TC07-2/GVI-1 lineage isolated in China. Infect. Genet. Evol. 2019, 75, 103942. [Google Scholar] [CrossRef]
- Chen, L.; Xiang, B.; Hong, Y.; Li, Q.; Du, H.; Lin, Q.; Liao, M.; Ren, T.; Xu, C. Phylogenetic analysis of infectious bronchitis virus circulating in southern China in 2016–2017 and evaluation of an attenuated strain as a vaccine candidate. Arch. Virol. 2021, 166, 73–81. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Martin, D.P. Recombination Detection and Analysis Using RDP3. Bioinform. DNA Seq. Anal. 2009, 537, 185–205. [Google Scholar] [CrossRef]
- Temmam, S.; Vongphayloth, K.; Baquero, E.; Munier, S.; Bonomi, M.; Regnault, B.; Douangboubpha, B.; Karami, Y.; Chrétien, D.; Sanamxay, D.; et al. Bat coronaviruses related to SARS-CoV-2 and infectious for human cells. Nature 2022, 604, 330–336. [Google Scholar] [CrossRef]
- Ji, J.; Gao, Y.; Chen, Q.; Wu, Q.; Xu, X.; Kan, Y.; Yao, L.; Bi, Y.; Xie, Q. Epidemiological investigation of avian infectious bronchitis and locally determined genotype diversity in central China: A 2016–2018 study. Poult. Sci. 2020, 99, 3001–3008. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-W.; Jackwood, M.W. Evidence of genetic diversity generated by recombination among avian coronavirus IBV. Arch. Virol. 2000, 145, 2135–2148. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Chen, W.; Shen, Y.; Xia, J.; Fan, S.; Li, N.; Luo, Y.; Han, X.; Cui, M.; Zhao, Y.; et al. Molecular characterization of infectious bronchitis virus in Southwestern China for the protective efficacy evaluation of four live vaccine strains. Vaccine 2022, 40, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, T.; Han, Z.; Liang, S.; Xu, Y.; Xu, Q.; Chen, Y.; Zhao, Y.; Shao, Y.; Li, H.; et al. Molecular and antigenic characteristics of Massachusetts genotype infectious bronchitis coronavirus in China. Vet. Microbiol. 2015, 181, 241–251. [Google Scholar] [CrossRef]
- Shan, D.; Fang, S.; Han, Z.; Ai, H.; Zhao, W.; Chen, Y.; Jiang, L.; Liu, S. Effects of hypervariable regions in spike protein on pathogenicity, tropism, and serotypes of infectious bronchitis virus. Virus Res. 2018, 250, 104–113. [Google Scholar] [CrossRef]
- Yan, W.; Qiu, R.; Wang, F.; Fu, X.; Li, H.; Cui, P.; Zhai, Y.; Li, C.; Zhang, L.; Gu, K.; et al. Genetic and pathogenic characterization of a novel recombinant avian infectious bronchitis virus derived from GI-1, GI-13, GI-28, and GI-19 strains in Southwestern China. Poult. Sci. 2021, 100, 101210. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IBV Isolates | Accession Number | Province 1 | Date (Day-Month-Year) | Vaccination History | Clinical Type | Length of S1 (nt/aa) 2 | Cleavage Recognition Motifs 3 | Genotype |
|---|---|---|---|---|---|---|---|---|
| 210007GXYL | OQ790027 | Guangxi | 05-08-2021 | H120 + H120 | A | 1620/540 | HRRRR | GI-19 |
| 210008GDYX | OQ790028 | Guangdong | 11-08-2021 | H120 + H120 | A | 1617/539 | RRSRR | GI-13 |
| 210018GXYL | OQ790029 | Guangxi | 06-09-2021 | H120 + H120 | RS | 1617/539 | RRSRR | GI-13 |
| 210021GXYL | OQ790030 | Guangxi | 06-09-2021 | H120 + H120 | I | 1617/539 | HRRRR | GI-13 |
| 210024GXYL | OQ790031 | Guangxi | 08-09-2021 | H120 + H120 | A | 1620/540 | RRSRR | GI-13 |
| 210031GXYL | OQ790032 | Guangxi | 16-09-2021 | H120 | RS | 1620/540 | RRSRR | GI-13 |
| 210059GXYL | OQ790033 | Guangxi | 29-10-2021 | H120 | RS | 1626/542 | SRRLR | — |
| 220198GDZC | OQ790035 | Guangdong | 07-11-2021 | H120 | RS | 1620/540 | HRRRR | GI-7 |
| 220149YNKM | OQ790036 | Yunnan | 18-11-2021 | H120 + H120 | RS | 1617/539 | RRFRR | — |
| 210085GXYL | OQ790037 | Guangxi | 03-11-2021 | H120 + H120 | I | 1620/540 | HRRRR | GI-19 |
| 210127GXYL | OQ790043 | Guangxi | 12-11-2021 | 4/91 | I | 1617/539 | RRSRR | GI-13 |
| 220150GXQZ | OQ790044 | Guangxi | 19-01-2022 | H120 + H120 | NP | 1620/540 | HRRRR | GI-19 |
| 220161YNKM | OQ790045 | Yunnan | 10-03-2022 | H120 | RS | 1611/537 | RRFRR | GI-1 |
| 210098GXNN | OQ790047 | Guangxi | 07-11-2021 | H120 | RS | 1620/540 | HRRRR | GI-19 |
| 210099GXNN | OQ790048 | Guangxi | 07-11-2021 | H120 | RS | 1620/540 | HRRRR | GI-19 |
| aa (%) | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| nt (%) | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | ||
| 1 | *** | 78.7 | 78.9 | 80.4 | 80.6 | 80.6 | 80 | 80.4 | 86.5 | 97.2 | 79.1 | 99.4 | 77.2 | 95 | 94.5 | 77 | 78.9 | 210007GXYL | |
| 2 | 78.6 | *** | 99.4 | 96.5 | 98.2 | 98.2 | 85.8 | 75.6 | 82.1 | 78.6 | 99.1 | 78.7 | 74.8 | 78.6 | 78 | 74.6 | 99.4 | 210008GDYX | |
| 3 | 78.6 | 99.8 | *** | 96.7 | 98.3 | 98.3 | 86 | 75.8 | 82.3 | 78.7 | 99.3 | 78.9 | 75 | 78.7 | 78.2 | 74.8 | 99.6 | 210018GXYL | |
| 4 | 80 | 97.5 | 97.5 | *** | 95.4 | 95.4 | 84.7 | 77.3 | 80.8 | 80.2 | 96.3 | 80.4 | 74.4 | 80 | 79.5 | 74.3 | 97 | 210021GXYL | |
| 5 | 79.8 | 98.8 | 98.8 | 96.5 | *** | 100 | 86.2 | 76 | 83.7 | 80.4 | 98 | 80.6 | 74.8 | 80.3 | 79.7 | 74.6 | 98.3 | 210024GXYL | |
| 6 | 79.8 | 98.8 | 98.8 | 96.5 | 100 | *** | 86.2 | 76 | 83.7 | 80.4 | 98 | 80.6 | 74.8 | 80.3 | 79.7 | 74.6 | 98.3 | 210031GXYL | |
| 7 | 79.9 | 87.9 | 87.9 | 87 | 88.2 | 88.2 | *** | 77.8 | 87.1 | 79.8 | 85.8 | 79.8 | 75.1 | 80 | 79.4 | 74.9 | 86 | 210059GXYL | |
| 8 | 79.8 | 77 | 77 | 78.4 | 77.2 | 77.2 | 78.4 | *** | 77.3 | 80.2 | 75.8 | 80.2 | 80 | 80.4 | 79.9 | 79.8 | 75.8 | 220198GDZC | |
| 9 | 85.5 | 85.8 | 85.8 | 84.6 | 86.6 | 86.6 | 89.3 | 77.9 | *** | 86.1 | 82.5 | 86.5 | 76.5 | 86.7 | 86.1 | 76.3 | 82.3 | 220149YNKM | |
| 10 | 97.3 | 78.6 | 78.6 | 79.9 | 79.8 | 79.8 | 79.9 | 79.4 | 85.5 | *** | 78.9 | 97.4 | 77 | 94.8 | 94.3 | 76.8 | 78.7 | 210085GXYL | |
| 11 | 78.7 | 99.7 | 99.8 | 97.4 | 98.7 | 98.7 | 87.9 | 77.1 | 86 | 78.7 | *** | 79.1 | 75.4 | 78.9 | 78.4 | 75.2 | 99.3 | 210127GXYL | |
| 12 | 99.4 | 78.7 | 78.7 | 80.2 | 80 | 80 | 80 | 79.7 | 85.6 | 97.4 | 78.9 | *** | 76.8 | 94.8 | 94.3 | 76.6 | 78.9 | 220150GXQZ | |
| 13 | 77.7 | 78.6 | 78.6 | 77.8 | 78.2 | 78.2 | 78.4 | 80.6 | 79.3 | 77.9 | 78.7 | 77.7 | *** | 77.7 | 77.2 | 99.6 | 75 | 220161YNKM | |
| 14 | 95.1 | 78.4 | 78.5 | 79.9 | 79.6 | 79.6 | 79.8 | 80.3 | 85.4 | 94.6 | 78.6 | 95.3 | 77.7 | *** | 99.3 | 77.6 | 78.7 | 210098GXNN | |
| 15 | 95.1 | 78.3 | 78.4 | 79.8 | 79.5 | 79.5 | 79.6 | 80.2 | 85.2 | 94.5 | 78.5 | 95.1 | 77.4 | 99.6 | *** | 77 | 78.2 | 210099GXNN | |
| 16 | 77.6 | 78.5 | 78.5 | 77.7 | 78 | 78 | 78.4 | 80.6 | 79.2 | 77.7 | 78.6 | 77.6 | 99.9 | 77.6 | 77.2 | *** | 74.8 | FJ888351_H120 vaccine | |
| 17 | 78.6 | 99.8 | 99.9 | 97.7 | 98.8 | 98.8 | 87.9 | 77 | 85.8 | 78.6 | 99.8 | 78.7 | 78.6 | 78.5 | 78.4 | 78.5 | *** | KF377577_4/91vaccine | |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | |||
| H120 | 4/91 | 210059GXYL | 220161YNKM | 210127GXYL | 220149YNKM | 210085GXYL | 220198GDZC | 210099GXNN | |
|---|---|---|---|---|---|---|---|---|---|
| H120 | 100 | 6.54 | 2.21 | 74.56 | 10.06 | 3.41 | 4.71 | 27.6 | 14.12 |
| 4/91 | 2.1 | 100 | 9.94 | 5.33 | 63.41 | 11.12 | 6.57 | 8.766 | 9.44 |
| 210059GXYL | 3.21 | 21.7 | 100 | 1.05 | 10.49 | 80.03 | 4.21 | 4.25 | 5.46 |
| 220161YNKM | 54.36 | 3.62 | 2.44 | 100 | 14.52 | 11.35 | 12.96 | 21.31 | 15.23 |
| 210127GXYL | 16.11 | 64.43 | 15.46 | 5.1 | 100 | 6.82 | 3.22 | 10.07 | 9.08 |
| 220149YNKM | 1.75 | 18.87 | 59.86 | 4.87 | 6.82 | 100 | 5.97 | 5.41 | 4.12 |
| 210085GXYL | 2.76 | 4.43 | 4.21 | 12.96 | 12.34 | 2.78 | 100 | 16.72 | 35.78 |
| 220198GDZC | 35.1 | 17.58 | 3.67 | 18.65 | 4.62 | 5.41 | 16.72 | 100 | 17.46 |
| 210099GCYL | 37.21 | 21.2 | 5.46 | 7.61 | 9.08 | 4.12 | 24.71 | 17.46 | 100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, W.; Lv, T.; Jiang, W.; Hou, Y.; Wang, Q.; Ren, J.; Fan, L.; Xiang, B.; Lin, Q.; Ding, C.; et al. Antigenic Characterization of Infectious Bronchitis Virus in the South China during 2021–2022. Viruses 2023, 15, 1273. https://doi.org/10.3390/v15061273
Yuan W, Lv T, Jiang W, Hou Y, Wang Q, Ren J, Fan L, Xiang B, Lin Q, Ding C, et al. Antigenic Characterization of Infectious Bronchitis Virus in the South China during 2021–2022. Viruses. 2023; 15(6):1273. https://doi.org/10.3390/v15061273
Chicago/Turabian StyleYuan, Weifeng, Ting Lv, Weiwei Jiang, Yuechi Hou, Qingyi Wang, Jinlian Ren, Lei Fan, Bin Xiang, Qiuyan Lin, Chan Ding, and et al. 2023. "Antigenic Characterization of Infectious Bronchitis Virus in the South China during 2021–2022" Viruses 15, no. 6: 1273. https://doi.org/10.3390/v15061273