
Detection and Molecular Characterization of Animal Adenovirus and Astrovirus from Western Maharashtra, India
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Nucleic Acid Extraction
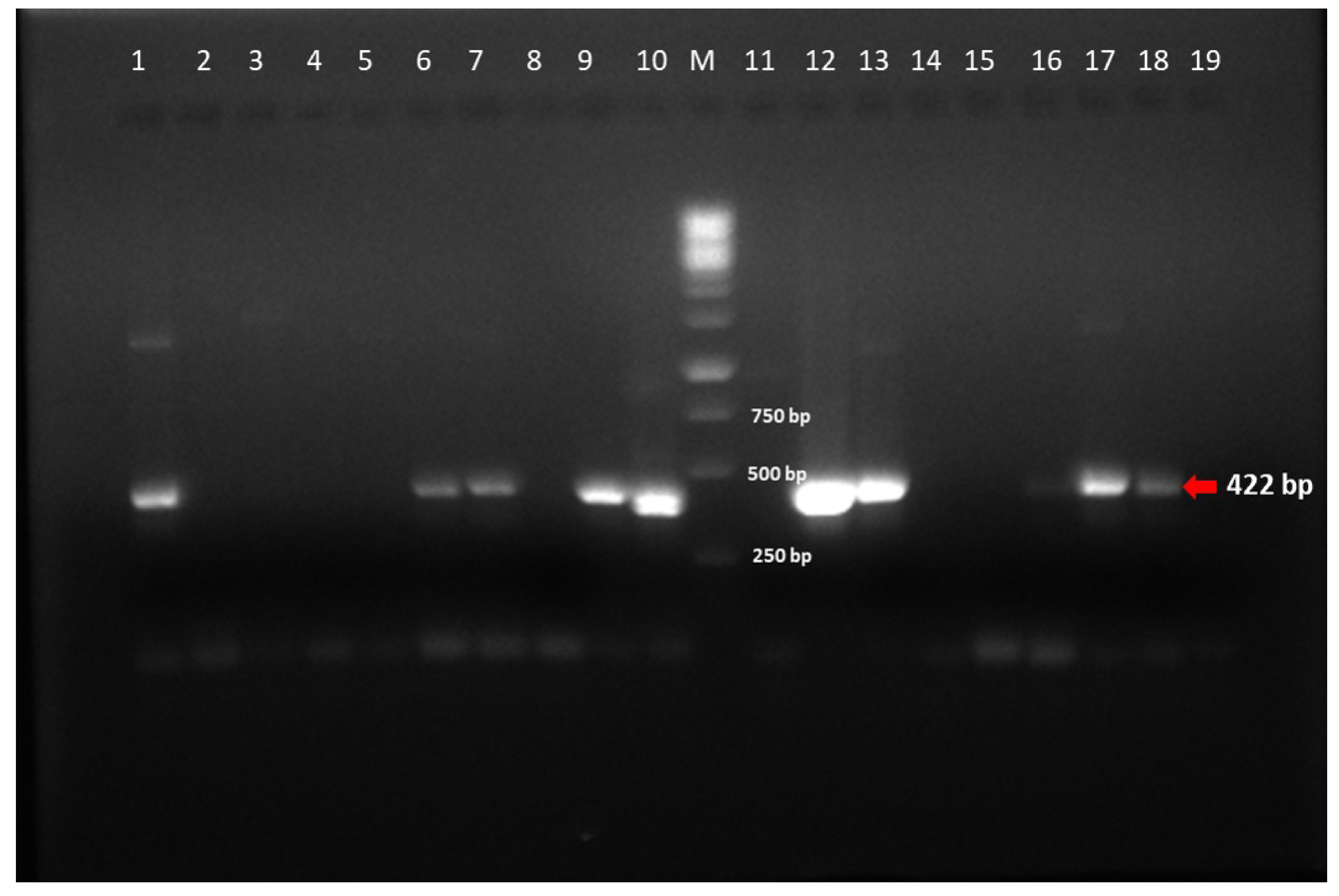
2.3. Conventional PCR for Adenovirus Detection
2.4. One-Step RT-PCR for Astrovirus Detection
2.5. Sequencing and Phylogenetic Analysis
3. Results
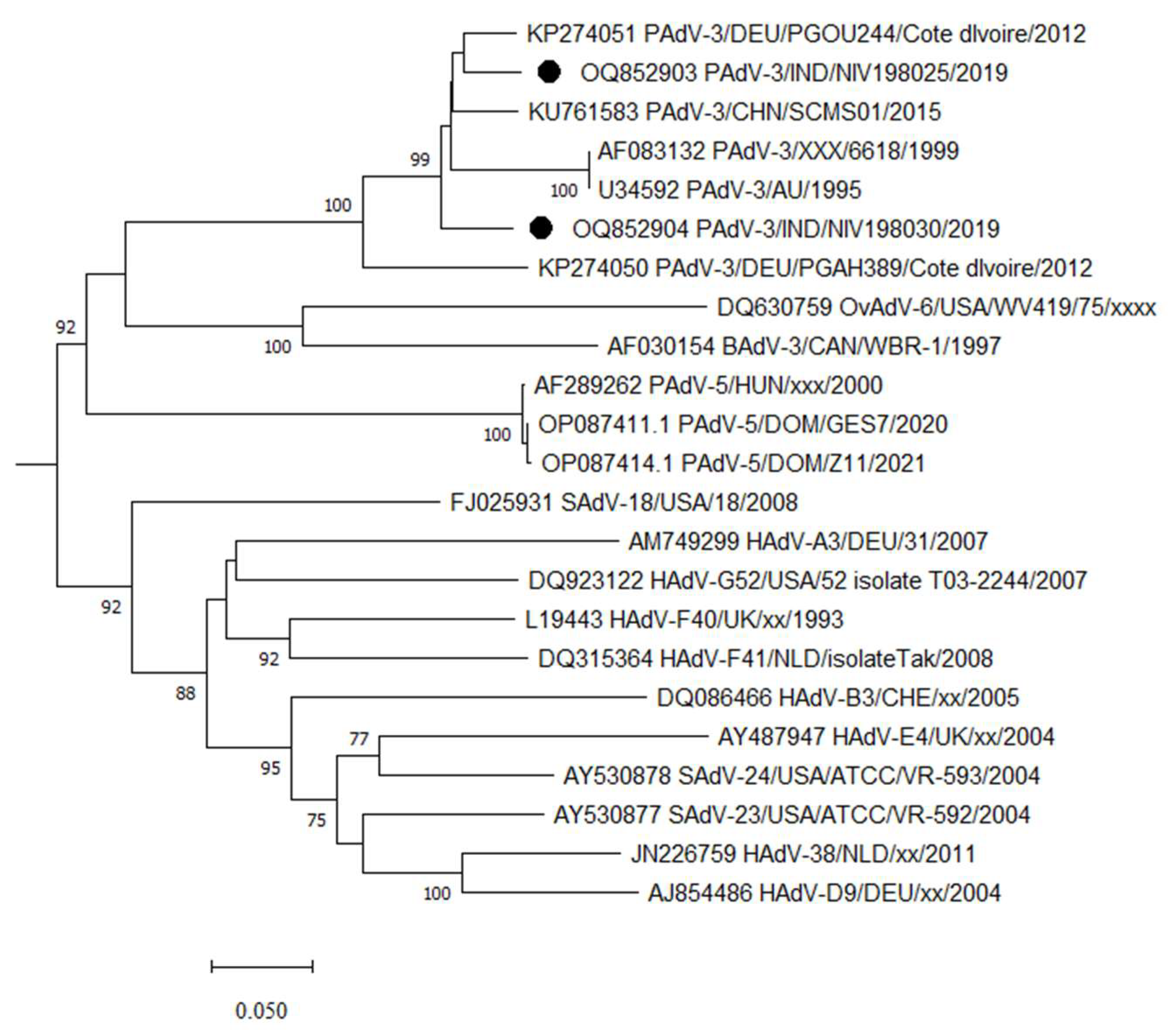
3.1. Adenovirus Detection and Phylogenetic Analysis
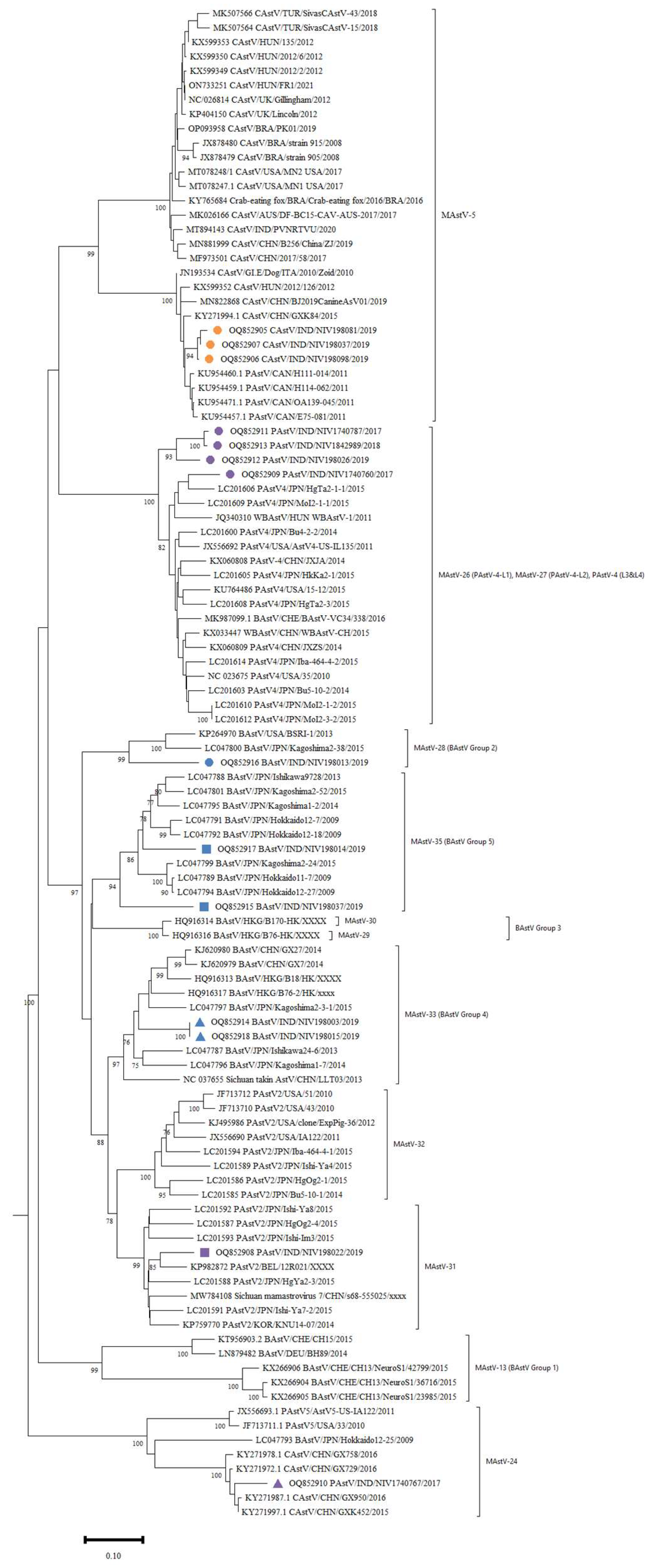
3.2. Astrovirus Detection and Phylogenetic Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Castells, M.; Colina, R. Viral Enteritis in Cattle: To Well Known Viruses and Beyond. Microbiol. Res. 2021, 12, 663–682. [Google Scholar] [CrossRef]
- Rawal, G.; Linhares, D.C.L. Scoping review on the epidemiology, diagnostics and clinical significance of porcine astroviruses. Transbound. Emerg. Dis. 2021, 69, 974–985. [Google Scholar] [CrossRef] [PubMed]
- Gaba, A.; Ayalew, L.E.; Tikoo, S.K. Animal Adenoviruses. In Recent Advances in Animal Virology; Malik, Y.S., Singh, R.K., Yadav, M.P., Eds.; Springer: Singapore, 2019; ISBN 978-981-13-9072-2. [Google Scholar]
- Benkő, M.; Aoki, K.; Arnberg, N.; Davison, A.J.; Echavarría, M.; Hess, M.; Jones, M.S.; Kaján, G.L.; Kajon, A.E.; Mittal, S.K.; et al. ICTV Report Consortium. ICTV Virus Taxonomy Profile: Adenoviridae 2022. J. Gen. Virol. 2022, 103, 001721. [Google Scholar] [CrossRef] [PubMed]
- Kumthip, K.; Khamrin, P.; Kongkaew, A.; Vachirachewin, R.; Malasao, R.; Ushijima, H.; Maneekarn, N. Molecular epidemiology and characterization of porcine adenoviruses in pigs with diarrhoea in Thailand. Infect. Genet. Evol. 2019, 67, 73–77. [Google Scholar] [CrossRef]
- Zhu, Q.; Li, B.; Sun, D. Bovine Astrovirus-A Comprehensive Review. Viruses 2022, 14, 1217. [Google Scholar] [CrossRef]
- Bosch, A.; King, A.M.; Lefkowitz, E.; Adams, M.J.; Carstens, E.B. Family Astroviridae. Virus Taxonomy: Classification and Nomenclature of Viruses (Ninth Report of the International Committee on the Taxonomy of Viruses), 9th ed.; Academic Press: New York, NY, USA, 2011. [Google Scholar]
- 20th Livestock Census. 2019. Available online: https://pib.gov.in/PressReleaseIframePage.aspx?PRID=1813802 (accessed on 1 July 2023).
- Iturriza Gómara, M.; Wong, C.; Blome, S.; Desselberger, U.; Gray, J. Molecular characterization of VP6 genes of human rotavirus isolates: Correlation of genogroups with subgroups and evidence of independent segregation. J. Virol. 2002, 76, 6596–6601. [Google Scholar] [CrossRef] [Green Version]
- Sawant, P.M.; Digraskar, S.S.; Gopalkrishna, V. Molecular characterization of unusual G10P[33], G6P[14] genomic constellations of group A rotavirus and evidence of zooanthroponosis in bovines. Infect. Genet. Evol. 2020, 84, 104385. [Google Scholar] [CrossRef]
- Sibley, S.D.; Goldberg, T.L.; Pedersen, J.A. Detection of Known and Novel Adenoviruses in Cattle Wastes via Broad-Spectrum Primers. Appl. Environ. Microbiol. 2011, 77, 5001–5008. [Google Scholar] [CrossRef] [Green Version]
- Chu, D.K.W.; Poon, L.L.M.; Guan, Y.; Peiris, J.S.M. Novel Astroviruses in Insectivorous Bats. J. Virol. 2008, 82, 9107–9114. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Qi, M.; Jiang, C.; Peng, Y.; Peng, Q.; Chen, Y.; Hu, C.; Chen, J.; Chen, X.; Chen, H.; et al. Prevalence of bovine astroviruses and their genotypes in sampled Chinese calves with and without diarrhoea. J. Gen. Virol. 2021, 2, 001640. [Google Scholar] [CrossRef]
- Ito, M.; Kuroda, M.; Masuda, T.; Akagami, M.; Haga, K.; Tsuchiaka, S.; Kishimoto, M.; Naoi, Y.; Sano, K.; Omatsu, T.; et al. Whole genome analysis of porcine astroviruses detected in Japanese pigs reveals genetic diversity and possible intra-genotypic recombination. Infect. Genet. Evol. 2017, 50, 38–48. [Google Scholar] [CrossRef]
- Zhou, H.; Liu, L.; Li, R.; Wang, G.; Wei, Z.; Ouyang, K.; Huang, W.; Chen, Y. Detection and genetic characterization of canine astroviruses in pet dogs in Guangxi, China. Virol. J. 2017, 14, 156. [Google Scholar] [CrossRef]
- Gainor, K.; Fortuna, Y.C.; Alakkaparambil, A.S.; González, W.; Malik, Y.S.; Ghosh, S. High Rates of Detection and Molecular Characterization of Porcine Adenovirus Serotype 5 (Porcine mastadenovirus C) from Diarrheic Pigs. Pathogens 2022, 11, 1210. [Google Scholar] [CrossRef]
- Vu, D.-L.; Bosch, A.; Pintó, R.M.; Guix, S. Epidemiology of Classic and Novel Human Astrovirus: Gastroenteritis and Beyond. Viruses 2017, 9, 33. [Google Scholar] [CrossRef] [Green Version]
- Borkenhagen, L.K.; Fieldhouse, J.K.; Seto, D.; Gray, G.C. Are adenoviruses zoonotic? A systematic review of the evidence. Emerg. Microbes Infect. 2019, 8, 1679–1687. [Google Scholar] [CrossRef] [Green Version]
- Mittal, S.K.; Ahi, Y.S.; Vemula, S.V. Xenogenic Adenoviral Vectors, Chapter 19. In Adenoviral Vectors for Gene Therapy, 2nd ed.; Curiel, D.T., Ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 495–528. ISBN 9780128002766. [Google Scholar]
- De Benedictis, P.; Schultz-Cherry, S.; Burnham, A.; Cattoli, G. Astrovirus infections in humans and animals—Molecular biology, genetic diversity, and interspecies transmissions. Infect. Genet. Evol. 2011, 11, 1529–1544. [Google Scholar] [CrossRef]
- Kumthip, K.; Khamrin, P.; Saikruang, W.; Kongkaew, A.; Vachirachewin, R.; Ushijima, H.; Maneekarn, N. Detection and genetic characterization of porcine astroviruses in piglets with and without diarrhoea in Thailand. Arch. Virol. 2018, 163, 1823–1829. [Google Scholar] [CrossRef]
- Li, J.-S.; Li, M.-Z.; Zheng, L.-S.; Liu, N.; Li, D.D.; Duan, Z.J. Identification and genetic characterization of two porcine astroviruses from domestic piglets in China. Arch. Virol. 2015, 160, 3079–3084. [Google Scholar] [CrossRef]
- Kattoor, J.J.; Malik, Y.S.; Saurabh, S.; Sircar, S.; Vinodhkumar, O.R.; Bora, D.P.; Dhama, K.; Ghosh, S.; Banyai, K.; Touil, N.; et al. First report and genetic characterization of porcine astroviruses of lineage 4 and 2 in diarrhoeic pigs in India. Transbound. Emerg. Dis. 2019, 66, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Kour, R.; Kumar, P.; Jindal, N.; Minhas, S.K.; Kumar, R.; Gupta, A.K.; Malik, A. Molecular detection and characterization reveals circulation of multiple genotypes of porcine astrovirus in Haryana, India. Arch. Virol. 2021, 166, 2847–2852. [Google Scholar] [CrossRef]
- Nagai, M.; Omatsu, T.; Aoki, H.; Otomaru, K.; Uto, T.; Koizumi, M.; Minami-Fukuda, F.; Takai, H.; Murakami, T.; Masuda, T.; et al. Full genome analysis of bovine astrovirus from fecal samples of cattle in Japan: Identification of possible interspecies transmission of bovine astrovirus. Arch. Virol. 2015, 160, 2491–2501. [Google Scholar] [CrossRef] [PubMed]
- Dema, A.; Tallapally, M.R.; Ganji, V.K.; Buddala, B.; Kodi, H.; Ramidi, A.; Yella, N.R.; Putty, K.A. Comprehensive molecular survey of viral pathogens associated with canine gastroenteritis. Arch. Virol. 2023, 168, 36. [Google Scholar] [CrossRef] [PubMed]
- Oem, J.K.; An, D.J. Phylogenetic analysis of bovine astrovirus in Korean cattle. Virus Genes 2014, 48, 372–375. [Google Scholar] [CrossRef] [PubMed]
- Sharp, C.P.; Gregory, W.F.; Mason, C.; Bronsvoort, B.M.D.; Beard, P.M. High prevalence and diversity of bovine astroviruses in the faeces of healthy and diarrhoeic calves in South West Scotland. Vet. Microbiol. 2015, 178, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Akdag, A.I.; Gupta, S.; Khan, N.; Upadhayay, A.; Ray, P. Epidemiology and clinical features of rotavirus, adenovirus, and astrovirus infections and coinfections in children with acute gastroenteritis prior to rotavirus vaccine introduction in Meerut, North India. J. Med. Virol. 2020, 92, 1102–1109. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Location | Health Condition | Sample Numbers | Virus Detected |
|---|---|---|---|---|
| Porcine (n = 48) | Chandkhed (22) | Diarrheic (18) | NIV1740787 | AstV, RVA |
| NIV1842989 | AstV | |||
| NIV1740785, NIV1740786, NIV1740788 | RVA | |||
| Apparently healthy (4) | Negative for RVA, AstV, AdV | |||
| Nandur (7) | Diarrheic (5) | NIV198025 | AstV, AdV | |
| NIV198030 | AstV, AdV | |||
| Apparently healthy (2) | NIV198026 | AstV | ||
| Shirwal (19) | Diarrheic (5) | Negative for RVA, AstV, AdV | ||
| Apparently healthy (14) | ||||
| Bovine (n = 65) | Manchar (48) | Diarrheic (41) | NIV1841887-890, NIV1841892-897, NIV1841899, NIV1841901, NIV1841904-907, NIV1841909 | RVA |
| Apparently healthy (7) | Negative for RVA, AstV, AdV | |||
| Tathwade (17) | Diarrheic (14) | NIV198003, NIV198013,NIV198015 | AstV | |
| NIV198014 | AstV, RVA | |||
| NIV198004, NIV198011 | RVA | |||
| Apparently healthy (3) | NIV198007 | AstV | ||
| Canine (n = 80) | Kasarsai | Diarrheic (55) | NIV198081, NIV198098 | AstV |
| Apparently healthy (25) | NIV198036 | AstV | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sawant, P.M.; Waghchaure, R.B.; Shinde, P.A.; Palikondawar, A.P.; Lavania, M. Detection and Molecular Characterization of Animal Adenovirus and Astrovirus from Western Maharashtra, India. Viruses 2023, 15, 1679. https://doi.org/10.3390/v15081679
Sawant PM, Waghchaure RB, Shinde PA, Palikondawar AP, Lavania M. Detection and Molecular Characterization of Animal Adenovirus and Astrovirus from Western Maharashtra, India. Viruses. 2023; 15(8):1679. https://doi.org/10.3390/v15081679
Chicago/Turabian StyleSawant, Pradeep M., Rishabh B. Waghchaure, Pooja A. Shinde, Avani P. Palikondawar, and Mallika Lavania. 2023. "Detection and Molecular Characterization of Animal Adenovirus and Astrovirus from Western Maharashtra, India" Viruses 15, no. 8: 1679. https://doi.org/10.3390/v15081679