
Insecticidal Traits of Variants in a Genotypically Diverse Natural Isolate of Anticarsia Gemmatalis Multiple Nucleopolyhedrovirus (AgMNPV)
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects, Cells, and Virus Isolates
2.2. DNA Extraction from OBs
2.3. Cloning of Genotypic Variants
2.4. Viral DNA Restriction Endonuclease Analysis
2.5. Biological Activity
2.6. Genotypic Variant Selection and Biological Characterization
3. Results
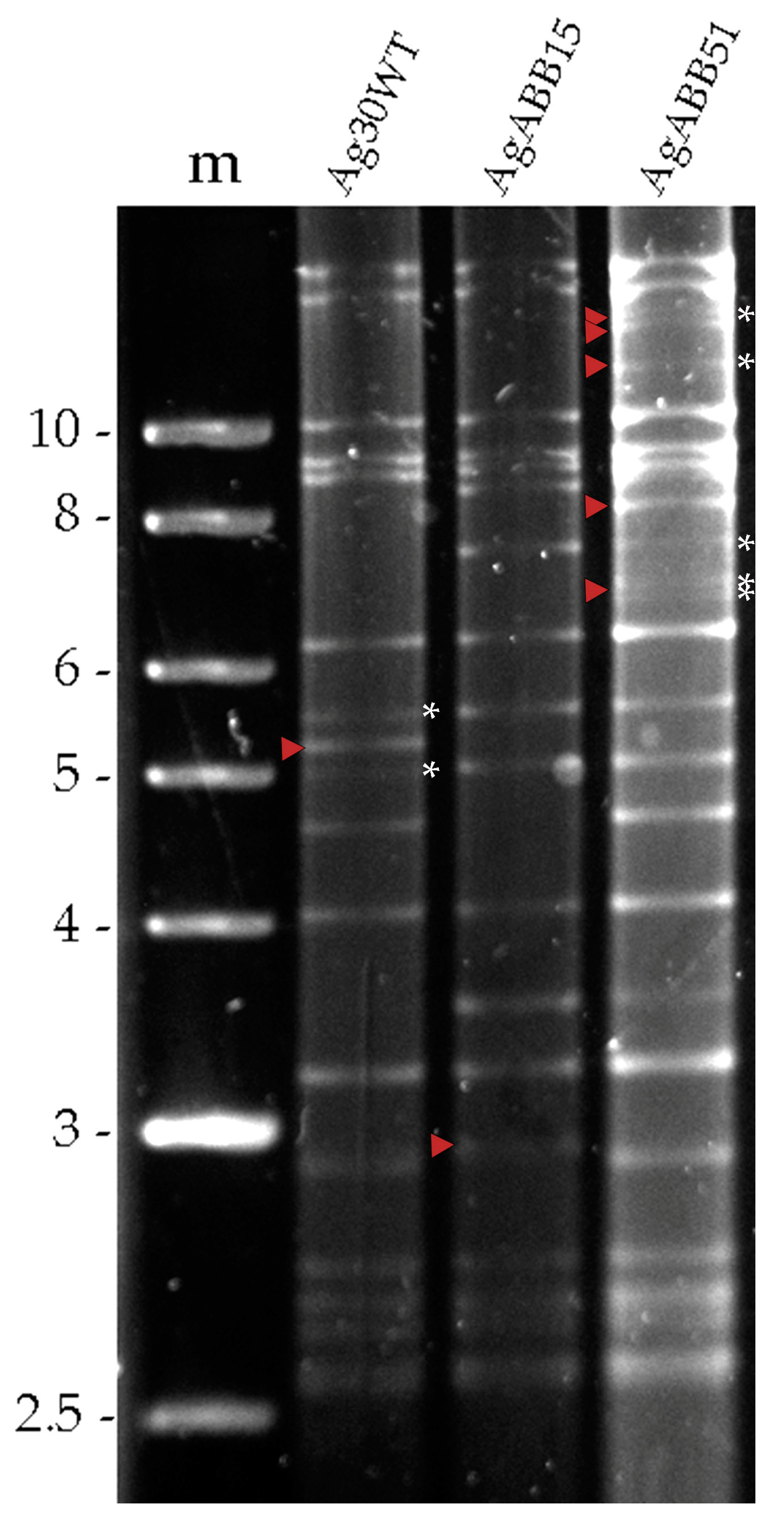
3.1. Identification of AgMNPV Isolates Using Restriction Endonuclease Analysis
3.2. Mortality Response to AgMNPV Isolates
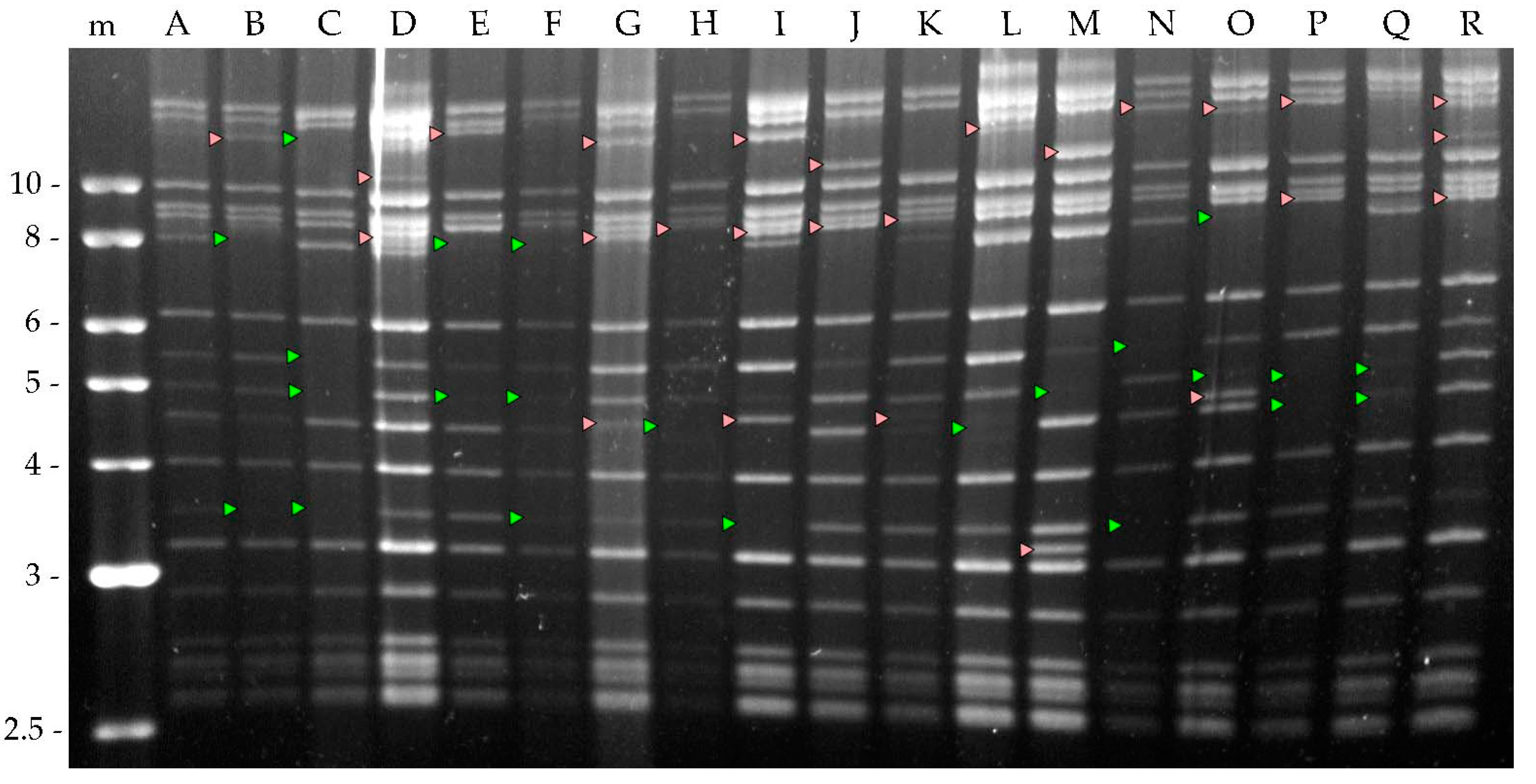
3.3. Biological and Genotypic Characterization of AgABB51
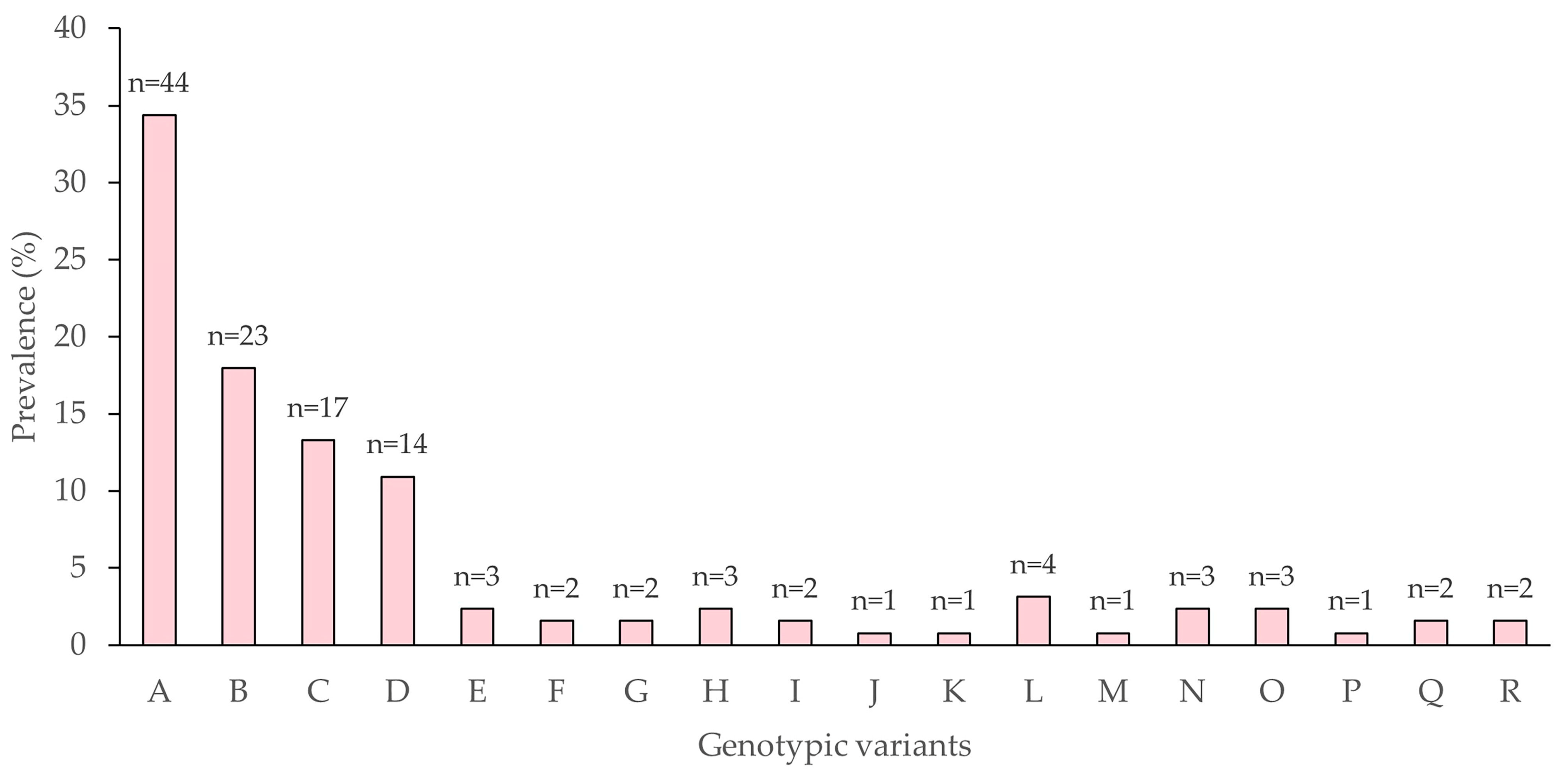
3.4. Biological Characterization of AgABB51 Genotypic Variants
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bueno, R.C.O.; de Bueno, A.F.; Moscardi, F.; Postali Parra, J.R.; Hoffmann-Campo, C.B. Lepidopteran Larva Consumption of Soybean Foliage: Basis for Developing Multiple-Species Economic Thresholds for Pest Management Decisions. Pest Manag. Sci. 2011, 67, 170–174. [Google Scholar] [CrossRef] [Green Version]
- Moscardi, F.; de Carvalho, R.C.Z. Consumo e utilização de folhas da soja por Anticarsia gemmatalis Hüb. (Lepidoptera: Noctuidae) infectada, em diferentes estádios larvais, por seu vírus de poliedrose nuclear. An.-Soc. Entomol. Bras. 1993, 22, 267–280. [Google Scholar] [CrossRef]
- Bayer Crop Science Bayer-Crop Pests Compendium-Anticarsia Gemmatalis by Bayer Crop Science AG. Available online: https://www.agriculture-xprt.com/products/bayer-anticarsia-gemmatalis-617566 (accessed on 24 May 2023).
- Musser, F.R.; Catchot, A.L.; Conley, S.P.; Davis, J.A.; Difonzo, C.; Greene, J.K.; Lorenz, G.M.; Owens, D.; Reisig, D.D.; Roberts, P.; et al. 2019 Soybean Insect Losses in the United States. Midsouth Entomol. 2020, 13, 1–23. [Google Scholar]
- Fernandes, F.O.; Abreu, J.Á.D.; Christ, L.M.; Rosa, A.P.S.A. Da Insecticides Management Used in Soybean for the Control of Anticarsia gemmatalis (Hübner, 1818) (Lepidoptera: Eribidae). J. Agric. Sci. 2018, 10, 223. [Google Scholar] [CrossRef] [Green Version]
- Degrande, P.E.; Vivan, L.M. Pragas Da Soja. Tecnologia e Produção: Soja e Milho 2011/2012; Fundação MT: Rondonópolis, Brazil, 2012; pp. 155–206. [Google Scholar]
- Murúa, M.G.; Vera, M.A.; Herrero, M.I.; Fogliata, S.V.; Michel, A. Defoliation of Soybean Expressing Cry1Ac by Lepidopteran Pests. Insects 2018, 9, 4–8. [Google Scholar] [CrossRef] [Green Version]
- Bueno, R.C.O.F.; Raetano, C.G.; Junior, J.D.; Carvalho, F.K. Integrated Management of Soybean Pests: The Example of Brazil. Outlooks Pest Manag. 2017, 28, 149–153. [Google Scholar] [CrossRef]
- Moscardi, F. Assessment of the Application of Baculoviruses for Control of Lepidoptera. Annu. Rev. Entomol. 1999, 44, 257–289. [Google Scholar] [CrossRef]
- Sosa-Gómez, D.R. Microbial Control of Soybean Pest Insects and Mites. In Microbial Control of Insect and Mite Pests: From Theory to Practice; Elsevier Inc.: Londrina, Brazil, 2017; pp. 199–208. ISBN 978-0-12-803566-5. [Google Scholar]
- Moscardi, F.; de Souza, M.L.; de Castro, M.E.B.; Lara Moscardi, M.; Szewczyk, B. Baculovirus Pesticides: Present State and Future Perspectives. In Microbes and Microbial Technology; Ahmad, I., Ahmad, F., Pichtel, J., Eds.; Springer: New York, NY, USA, 2011; pp. 415–445. ISBN 978-1-4419-7930-8. [Google Scholar]
- Valicente, F.H. Entomopathogenic Viruses. In Natural Enemies of Insect Pests in Neotropical Agroecosystems; Souza, B., Vázquez, L.L., Marucci, R.C., Eds.; Springer: Cham, Switzerland, 2019; pp. 137–150. ISBN 978-3-030-24733-1. [Google Scholar]
- Popham, H.J.R.; Rowley, D.L.; Harrison, R.L. Differential Insecticidal Properties of Spodoptera frugiperda multiple nucleopolyhedrovirus Isolates against Corn-Strain and Rice-Strain Fall Armyworm, and Genomic Analysis of Three Isolates. J. Invertebr. Pathol. 2021, 183, 107561. [Google Scholar] [CrossRef]
- Rowley, D.L.; Popham, H.J.R.; Harrison, R.L. Genetic Variation and Virulence of Nucleopolyhedroviruses Isolated Worldwide from the Heliothine Pests Helicoverpa armigera, Helicoverpa zea, and Heliothis virescens. J. Invertebr. Pathol. 2011, 107, 112–126. [Google Scholar] [CrossRef]
- Clavijo, G.; Williams, T.; Muñoz, D.; Caballero, P.; López-Ferber, M. Mixed Genotype Transmission Bodies and Virions Contribute to the Maintenance of Diversity in an Insect Virus. Proc. Biol. Sci. 2010, 277, 943–951. [Google Scholar] [CrossRef]
- Erlandson, M.A. Genetic Variation in Field Populations of Baculoviruses: Mechanisms for Generating Variation and Its Potential Role in Baculovirus Epizootiology. Virol. Sin. 2009, 24, 458–469. [Google Scholar] [CrossRef]
- de Brito, A.F.; Braconi, C.T.; Weidmann, M.; Dilcher, M.; Pereira Alves, J.M.; Gruber, A.; de Andrade Zanotto, P.M. The Pangenome of the Anticarsia gemmatalis multiple nucleopolyhedrovirus (AgMNPV). Genome Biol. Evol. 2016, 8, 94–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croizier, G.; Ribeiro, H.C.T. Recombination as a Possible Major Cause of Genetic Heterogeneity in Anticarsia gemmatalis nuclear polyhedrosis virus Wild Populations. Virus Res. 1992, 26, 183–196. [Google Scholar] [CrossRef]
- Cabodevilla, O.; Ibañez, I.; Simón, O.; Murillo, R.; Caballero, P.; Williams, T. Occlusion Body Pathogenicity, Virulence and Productivity Traits Vary with Transmission Strategy in a Nucleopolyhedrovirus. Biol. Control 2011, 56, 184–192. [Google Scholar] [CrossRef]
- Harrison, R.L.; Puttler, B.; Popham, H.J.R. Genomic Sequence Analysis of a Fast-Killing Isolate of Spodoptera frugiperda multiple nucleopolyhedrovirus. J. Gen. Virol. 2008, 89, 775–790. [Google Scholar] [CrossRef]
- Aguirre, E.; Beperet, I.; Williams, T.; Caballero, P. Genetic Variability of Chrysodeixis includens nucleopolyhedrovirus (ChinNPV) and the Insecticidal Characteristics of Selected Genotypic Variants. Viruses 2019, 11, 581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del-Angel, C.; Lasa, R.; Rodríguez-del-Bosque, L.A.; Mercado, G.; Beperet, I.; Caballero, P.; Williams, T. Anticarsia gemmatalis nucleopolyhedrovirus from Soybean Crops in Tamaulipas, Mexico: Diversity and Insecticidal Characteristics of Individual Variants and Their Co-Occluded Mixtures. Fla. Entomol. 2018, 101, 404–410. [Google Scholar] [CrossRef]
- Smits, P.H.; Vlak, J.M. Registration of the First Viral Insecticide in the Netherlands: The Development of SPOD-X, Based on Spodoptera exigua nuclear polyhedrosis virus. Meded. Van De Fac. Landbouwwet. Rijksuniv. Gent 1994, 59, 385–392. [Google Scholar]
- Niz, J.M.; Salvador, R.; Ferrelli, M.L.; de Cap, A.S.; Romanowski, V.; Berretta, M.F. Genetic Variants in Argentinean Isolates of Spodoptera frugiperda multiple nucleopolyhedrovirus. Virus Genes 2020, 56, 401–405. [Google Scholar] [CrossRef]
- Masson, T.; Fabre, M.L.; Pidre, M.L.; Niz, J.M.; Berretta, M.F.; Romanowski, V.; Ferrelli, M.L. Genomic Diversity in a Population of Spodoptera frugiperda nucleopolyhedrovirus. Infect. Genet. Evol. 2021, 90, 104749. [Google Scholar] [CrossRef]
- Baillie, V.L.; Bouwer, G. High Levels of Genetic Variation within Helicoverpa armigera nucleopolyhedrovirus Populations in Individual Host Insects. Arch. Virol. 2012, 157, 2281–2289. [Google Scholar] [CrossRef] [PubMed]
- Cory, J.S.; Green, B.M.; Paul, R.K.; Hunter-Fujita, F. Genotypic and Phenotypic Diversity of a Baculovirus Population within an Individual Insect Host. J. Invertebr. Pathol. 2005, 89, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.H.; Miller, L.K. Isolation of Genotypic Variants of Autographa californica nuclear polyhedrosis virus. J. Virol. 1978, 27, 754–767. [Google Scholar] [CrossRef] [Green Version]
- Arrizubieta, M.; Simón, O.; Caballero, P.; Williams, T. Nuevos Genotipos Del Nucleopoliedrovirus Simple de Helicoverpa Armigera (HearSNPV), Procedimiento Para Su Producción y Uso Como Agente de Control Biológico. Patent ES2555165, 29 December 2015. [Google Scholar]
- Caballero, P.; Bernal, A.; Simón, O.; Carnero, A.; Hernández-Suárez, E.; Williams, T.G. Nuevos Genotipos Del Nucleopoliedrovirus Simple de Chrysodeixis Chalcites (ChchSNPV), Procedimiento Para su Producción y Uso Como Agente de Control Biológico. Patent ES2504866, 8 October 2014. [Google Scholar]
- Caballero, P.; Murillo, R.; Muñoz, D.; Williams, T. El Nucleopoliedrovirus de Spodoptera exigua (Lepidoptera: Noctuidae) Como Bioplaguicida: Análisis de Avances Recientes En España. Rev. Colomb Entomol. 2009, 35, 105–115. [Google Scholar] [CrossRef]
- Oliveira, J.V.; Wolff, J.L.C.; Garcia-Maruniak, A.; Ribeiro, B.M.; de Castro, M.E.B.; de Souza, M.L.; Moscardi, F.; Maruniak, J.E.; de Andrade Zanotto, P.M. Genome of the Most Widely Used Viral Biopesticide: Anticarsia gemmatalis multiple nucleopolyhedrovirus. J. Gen. Virol. 2006, 87, 3233–3250. [Google Scholar] [CrossRef] [PubMed]
- Allen, G.E.; Knell, J.D. A Nuclear Polyhedrosis Virus of Anticarsia gemmatalis: I. Ultrastructure, Replication, and Pathogenicity. Fla. Entomol. 1977, 60, 233. [Google Scholar] [CrossRef]
- Maruniak, J.E.; Garcia-Maruniak, A.; Souza, M.L.; Zanotto, P.M.A.; Moscardi, F. Physical Maps and Virulence of Anticarsia gemmatalis nucleopolyhedrovirus Genomic Variants. Arch. Virol. 1999, 144, 1991–2006. [Google Scholar] [CrossRef]
- Fan, J.; Jehle, J.A.; Rucker, A.; Nielsen, A.L. First Evidence of CpGV Resistance of Codling Moth in the USA. Insects 2022, 13, 533. [Google Scholar] [CrossRef]
- Jehle, J.A.; Schulze-Bopp, S.; Undorf-Spahn, K.; Fritsch, E. Evidence for a Second Type of Resistance against Cydia pomonella granulovirus in Field Populations of Codling Moths. Appl. Environ. Microb. 2016, 83, e02330-16. [Google Scholar] [CrossRef] [Green Version]
- Hoover, K.; Washburn, J.O.; Volkman, L.E. Midgut-Based Resistance of Heliothis virescens to Baculovirus Infection Mediated by Phytochemicals in Cotton. J. Insect. Physiol. 2000, 46, 999–1007. [Google Scholar] [CrossRef]
- Eberle, K.E.; Asser-Kaiser, S.; Sayed, S.M.; Nguyen, H.T.; Jehle, J.A. Overcoming the Resistance of Codling Moth against Conventional Cydia pomonella granulovirus (CpGV-M) by a New Isolate CpGV-I12. J. Invertebr. Pathol. 2008, 98, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Gebhardt, M.M.; Eberle, K.E.; Radtke, P.; Jehle, J.A. Baculovirus Resistance in Codling Moth Is Virus Isolate-Dependent and the Consequence of a Mutation in Viral Gene Pe38. Proc. Natl. Acad. Sci. USA 2014, 111, 15711–15716. [Google Scholar] [CrossRef]
- Abot, A.R.; Moscardi, F.; Fuxa, J.R.; Sosa-Gómez, D.R.; Richter, A.R. Development of Resistance By Anticarsia gemmatalis from Brazil and the United States to a Nuclear Polyhedrosis Virus under Laboratory Selection Pressure. Biol. Control. 1996, 7, 126–130. [Google Scholar] [CrossRef]
- Levy, S.M.; Falleiros, A.M.F.; Moscardi, F.; Gregório, E.A. Susceptibility/Resistance of Anticarsia gemmatalis Larvae to Its Nucleopolyhedrovirus (AgMNPV): Structural Study of the Peritrophic Membrane. J. Invertebr. Pathol. 2007, 96, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Barrera, G.; Williams, T.; Villamizar, L.; Caballero, P.; Simón, O. Deletion Genotypes Reduce Occlusion Body Potency but Increase Occlusion Body Production in a Colombian Spodoptera frugiperda nucleopolyhedrovirus Population. PLoS ONE 2013, 8, e77271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greene, G.L.; Leppla, N.C.; Dickerson, W.A. Velvetbean Caterpillar: A Rearing Procedure and Artificial Medium123. J. Econ. Entomol. 1976, 69, 487–488. [Google Scholar] [CrossRef]
- King, L.A.; Possee, R.D. The Baculovirus Expression System. A Laboratory Guide; Chapman & Hall: London, UK, 1992; ISBN 0-412-37150-2. [Google Scholar]
- Hughes, P.R.; Wood, H.A. In Vivo and in Vitro Bioassay Methods for Baculoviruses. In The Biology of Baculoviruses; Granados, R.R., Federici, B.A., Eds.; CRC Press: Boca Raton, FL, USA, 1986; Volume 2, pp. 1–30. [Google Scholar]
- Polo Plus, version 1.0; LeOra Software: Parma, MO, USA, 2003.
- Therneau, T.M. A Package for Survival Analysis in R; 2023. Available online: https://CRAN.Rproject.org/package=survival (accessed on 5 April 2023).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- IBM SPSS Statistics. IBM SPSS Statistics for Windows, version 25.0; IBM Corp: Armonk, NY, USA, 2017. [Google Scholar]
- Ferreira, B.C.; Melo, F.L.; Silva, A.M.R.; Sanches, M.M.; Moscardi, F.; Ribeiro, B.M.; Souza, M.L. Biological and Molecular Characterization of Two Anticarsia gemmatalis multiple nucleopolyhedrovirus Clones Exhibiting Contrasting Virulence. J. Invertebr. Pathol. 2019, 164, 23–31. [Google Scholar] [CrossRef]
- da Silveira, E.B.; Cordeiro, B.A.; Ribeiro, B.M.; de Castro, M.E.B.; Soares, E.F.; Báo, S.N. An Anticarsia gemmatalis multiple nucleopolyhedrovirus Mutant, VApAg, Induces Hemocytes Apoptosis in Vivo and Displays Reduced Infectivity in Larvae of Anticarsia gemmatalis (Hübner) (Lepidoptera: Noctuidae). Virus Res. 2007, 130, 182–192. [Google Scholar] [CrossRef] [Green Version]
- Corsaro, B.G.; Fraser, M.J. Characterization of Genotypic and Phenotypic Variation in Plaque-Purified Strains of HzSNPV Elkar Isolate. Intervirology 1987, 28, 185–198. [Google Scholar] [CrossRef]
- Hodgson, D.J.; Vanbergen, A.J.; Watt, A.D.; Hails, R.S.; Cory, J.S. Phenotypic Variation between Naturally Co-Existing Genotypes of a Lepidopteran Baculovirus. Evol. Ecol. Res. 2001, 3, 687–701. [Google Scholar]
- Ribeiro, H.C.T.; Pavan, O.H.O.; Muotri, A.R. Comparative Susceptibility of Two Different Hosts to Genotypic Variants of the Anticarsia gemmatalis nuclear polyhedrosis virus. Entomol. Exp. Appl. 1997, 83, 233–237. [Google Scholar] [CrossRef]
- Ogembo, J.G.; Chaeychomsri, S.; Kamiya, K.; Ishikawa, H.; Katou, Y.; Ikeda, M.; Kobayashi, M. Cloning and Comparative Characterization of Nucleopolyhedroviruses Isolated from African Bollworm, Helicoverpa armigera, (Lepidoptera: Noctudiae) in Different Geographic Regions. J. Insect. Biotechnol. Sericol. 2007, 76, 1_39–1_49. [Google Scholar] [CrossRef]
- Bernal, A.; Simón, O.; Williams, T.; Muñoz, D.; Caballero, P. A Chrysodeixis chalcites single-nucleocapsid nucleopolyhedrovirus Population from the Canary Islands Is Genotypically Structured To Maximize Survival. Appl. Environ. Microb. 2013, 79, 7709–7718. [Google Scholar] [CrossRef] [Green Version]
- Simon, O.; Williams, T.; Lopez-Ferber, M.; Caballero, P. Genetic Structure of a Spodoptera frugiperda nucleopolyhedrovirus Population: High Prevalence of Deletion Genotypes. Appl. Environ. Microb. 2004, 70, 5579–5588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pijlman, G.P.; van den Born, E.; Martens, D.E.; Vlak, J.M. Autographa californica baculoviruses with Large Genomic Deletions Are Rapidly Generated in Infected Insect Cells. Virology 2001, 283, 132–138. [Google Scholar] [CrossRef] [Green Version]
- Kool, M.; Voncken, J.W.; van Lier, F.L.; Tramper, J.; Vlak, J.M. Detection and Analysis of Autographa californica nuclear polyhedrosis virus Mutants with Defective Interfering Properties. Virology 1991, 183, 739–746. [Google Scholar] [CrossRef]
- Smith, I.R.L.; Crook, N.E. In Vivo Isolation of Baculovirus Genotypes. Virology 1988, 166, 240–244. [Google Scholar] [CrossRef]
- Muñoz, D.; Castillejo, J.I.; Caballero, P. Naturally Occurring Deletion Mutants Are Parasitic Genotypes in a Wild-Type Nucleopolyhedrovirus Population of Spodoptera exigua. Appl. Environ. Microb. 1998, 64, 4372–4377. [Google Scholar] [CrossRef] [Green Version]
- Kumar, C.; Sathiah, N.; Rabindra, R. Optimizing the Time of Harvest of Nucleopolyhedrovirus Infected Spodoptera litura (Fabricius) Larvae under in Vivo Production Systems. Curr. Sci. India 2005, 88, 1682–1684. [Google Scholar]
- Cory, J.S.; Myers, J.H. Adaptation in an Insect Host–Plant Pathogen Interaction. Ecol. Lett. 2004, 7, 632–639. [Google Scholar] [CrossRef]
- Simón, O.; Williams, T.; López-Ferber, M.; Taulemesse, J.-M.; Caballero, P. Population Genetic Structure Determines Speed of Kill and Occlusion Body Production in Spodoptera frugiperda multiple nucleopolyhedrovirus. Biol. Control. 2008, 44, 321–330. [Google Scholar] [CrossRef]
- Williams, T.; López-Ferber, M.; Caballero, P. Nucleopolyhedrovirus Coocclusion Technology: A New Concept in the Development of Biological Insecticides. Front. Microbiol. 2022, 12, 810026. [Google Scholar] [CrossRef] [PubMed]
- Sanjuán, R. Collective Infectious Units in Viruses. Trends Microbiol. 2017, 25, 402–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leeks, A.; Sanjuán, R.; West, S.A. The Evolution of Collective Infectious Units in Viruses. Virus Res. 2019, 265, 94–101. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | 103 OBs/mL | 105 OBs/mL | 107 OBs/mL |
|---|---|---|---|
| Ag30WT | 8 ± 4 (78) | 58 ± 8 (73) | 98 ± 2 (83) |
| AgABB15 | 11 ± 5 (78) | 72 ± 8 (81) | 100 ± 0 (79) |
| AgABB51 | 10 ± 6 (82) | 87 ± 1 (77) | 100 ± 0 (74) |
| Virus | LC50 * (OBs/mL) | 95% Confidence Limits | MTD (h) | 95% Confidence Limits | OB Production (OBs/Larva) | 95% Confidence Limits | |||
|---|---|---|---|---|---|---|---|---|---|
| Low | High | Low | High | Low | High | ||||
| AgABB51 | 1.1 × 104 | 8.5 × 103 | 1.5 × 104 | 138.6 | 134.3 | 143.0 | 1.23 × 109 | 1.15 × 109 | 1.31 × 109 |
| Variant | LC50 (OBs/mL) | Relative Potency | 95% Confidence Limits | LC90 (OBs/mL) | Relative Potency | 95% Confidence Limits | Slope (±SE) | ||
|---|---|---|---|---|---|---|---|---|---|
| Low | High | Low | High | ||||||
| AgABB51 | 1.0 × 104 | 1 | - | - | 6.9 × 104 | 1 | - | - | 1.54 ± 0.128 |
| D | 8.8 × 103 | 1.15 | 0.84 | 1.56 | 5.3 × 104 | 1.30 | 0.77 | 2.11 | 1.64 ± 0.138 |
| E | 8.0 × 103 | 1.26 | 0.92 | 1.72 | 4.4 × 104 | 1.55 | 0.91 | 2.65 | 1.72 ± 0.147 |
| I | 7.6 × 103 | 1.33 | 0.97 | 1.81 | 4.1 × 104 | 1.68 | 1.00 | 2.9 | 1.75 ± 0.152 |
| L | 9.2 × 103 | 1.10 | 0.80 | 1.50 | 5.6 × 104 | 1.21 | 0.70 | 2.13 | 1.62 ± 0.141 |
| M | 9.2 × 103 | 1.10 | 0.74 | 1.36 | 4.9 × 104 | 1.40 | 0.83 | 2.35 | 1.65 ± 0.135 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parras-Jurado, A.; Muñoz, D.; Beperet, I.; Williams, T.; Caballero, P. Insecticidal Traits of Variants in a Genotypically Diverse Natural Isolate of Anticarsia Gemmatalis Multiple Nucleopolyhedrovirus (AgMNPV). Viruses 2023, 15, 1526. https://doi.org/10.3390/v15071526
Parras-Jurado A, Muñoz D, Beperet I, Williams T, Caballero P. Insecticidal Traits of Variants in a Genotypically Diverse Natural Isolate of Anticarsia Gemmatalis Multiple Nucleopolyhedrovirus (AgMNPV). Viruses. 2023; 15(7):1526. https://doi.org/10.3390/v15071526
Chicago/Turabian StyleParras-Jurado, Ana, Delia Muñoz, Inés Beperet, Trevor Williams, and Primitivo Caballero. 2023. "Insecticidal Traits of Variants in a Genotypically Diverse Natural Isolate of Anticarsia Gemmatalis Multiple Nucleopolyhedrovirus (AgMNPV)" Viruses 15, no. 7: 1526. https://doi.org/10.3390/v15071526