
Comprehensive Comparison of Baculoviral and Plasmid Gene Delivery in Mammalian Cells
 , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
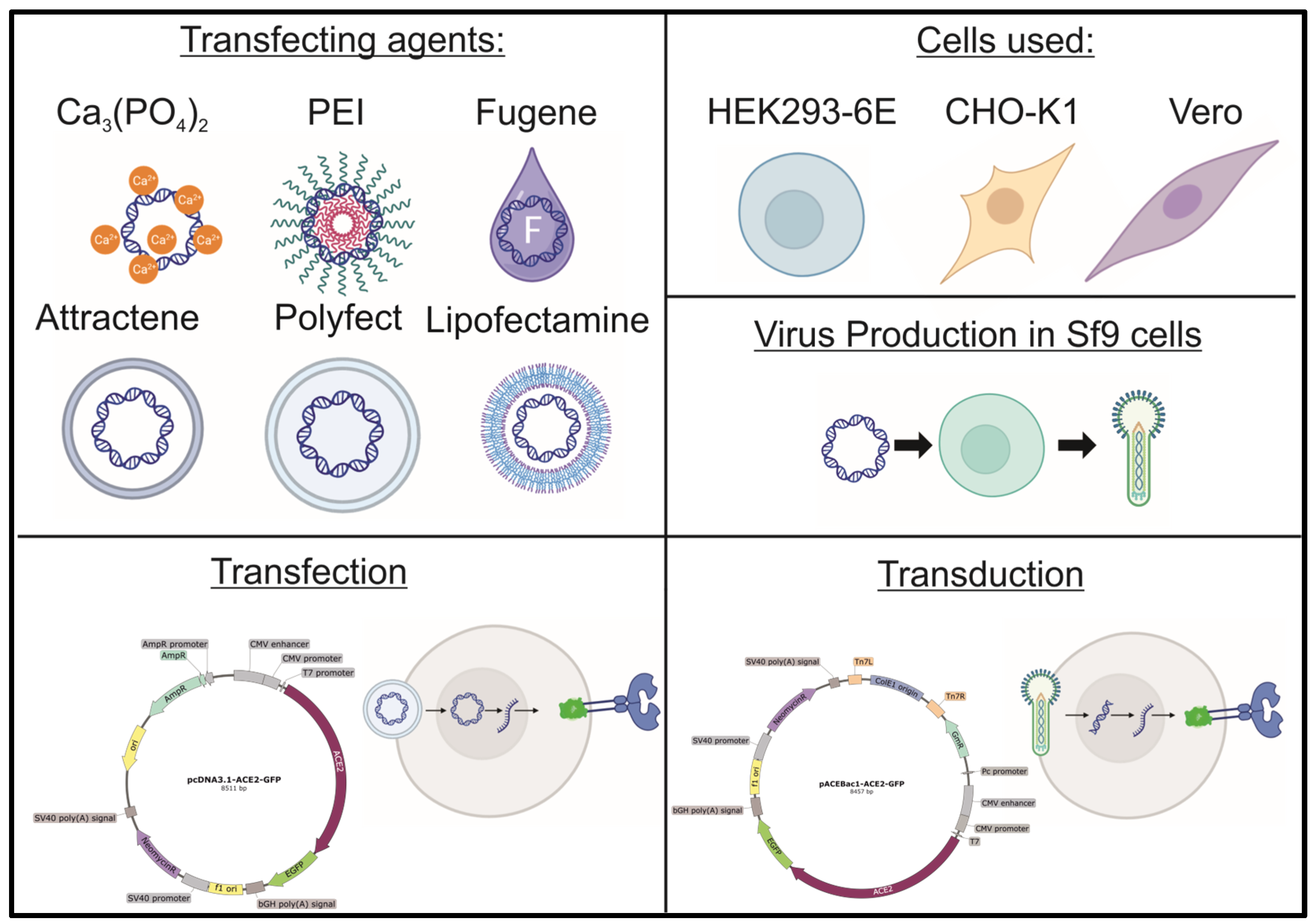
2. Materials and Methods
2.1. Cell Culture
2.2. Baculovirus Production
2.3. Ultracentrifugation of Baculorivus
2.4. Transfection
2.5. Transduction
2.6. Viability and Fluorescence Measurements
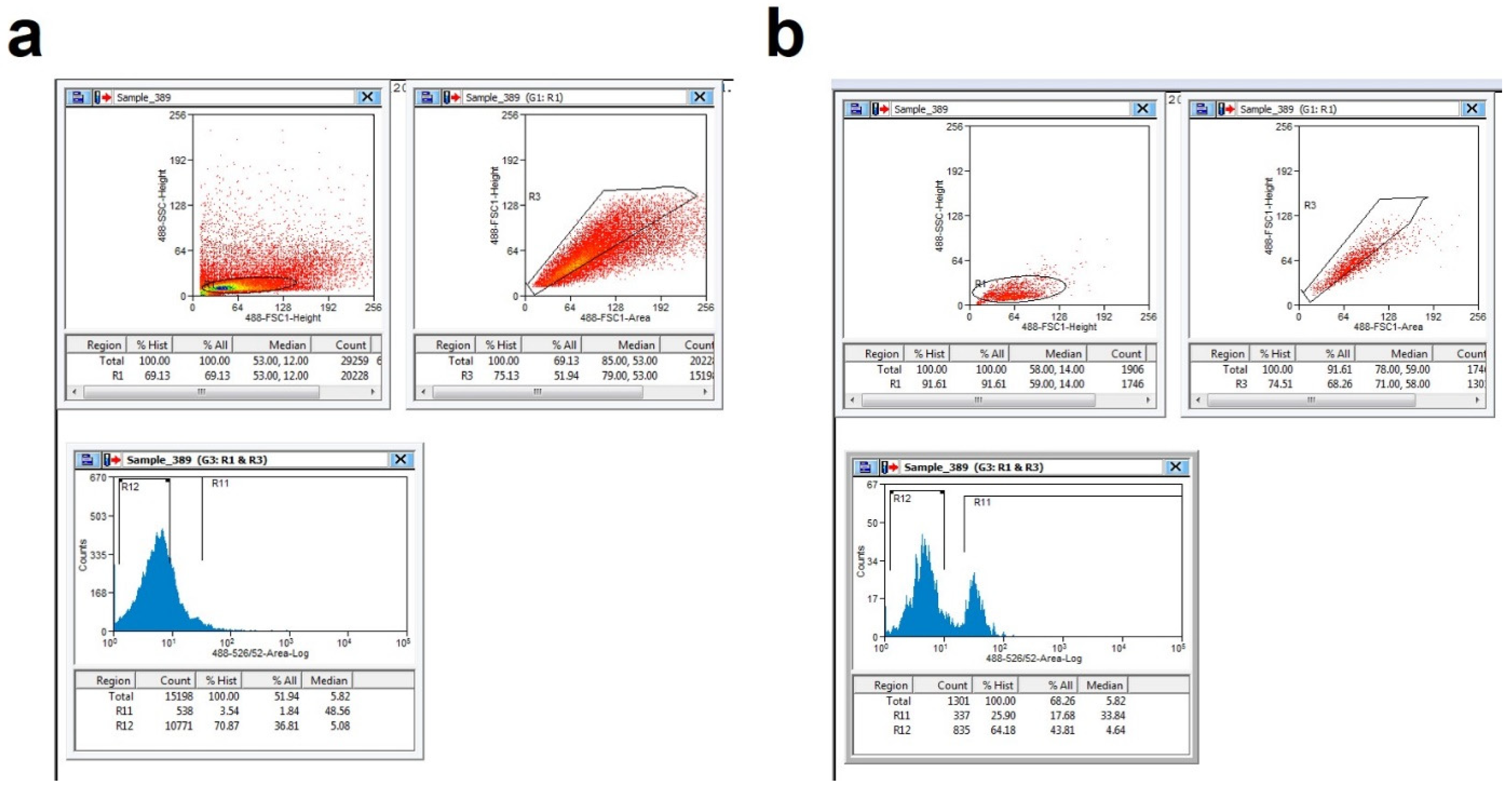
2.7. Cell Sorting
2.8. Data Analysis
3. Results
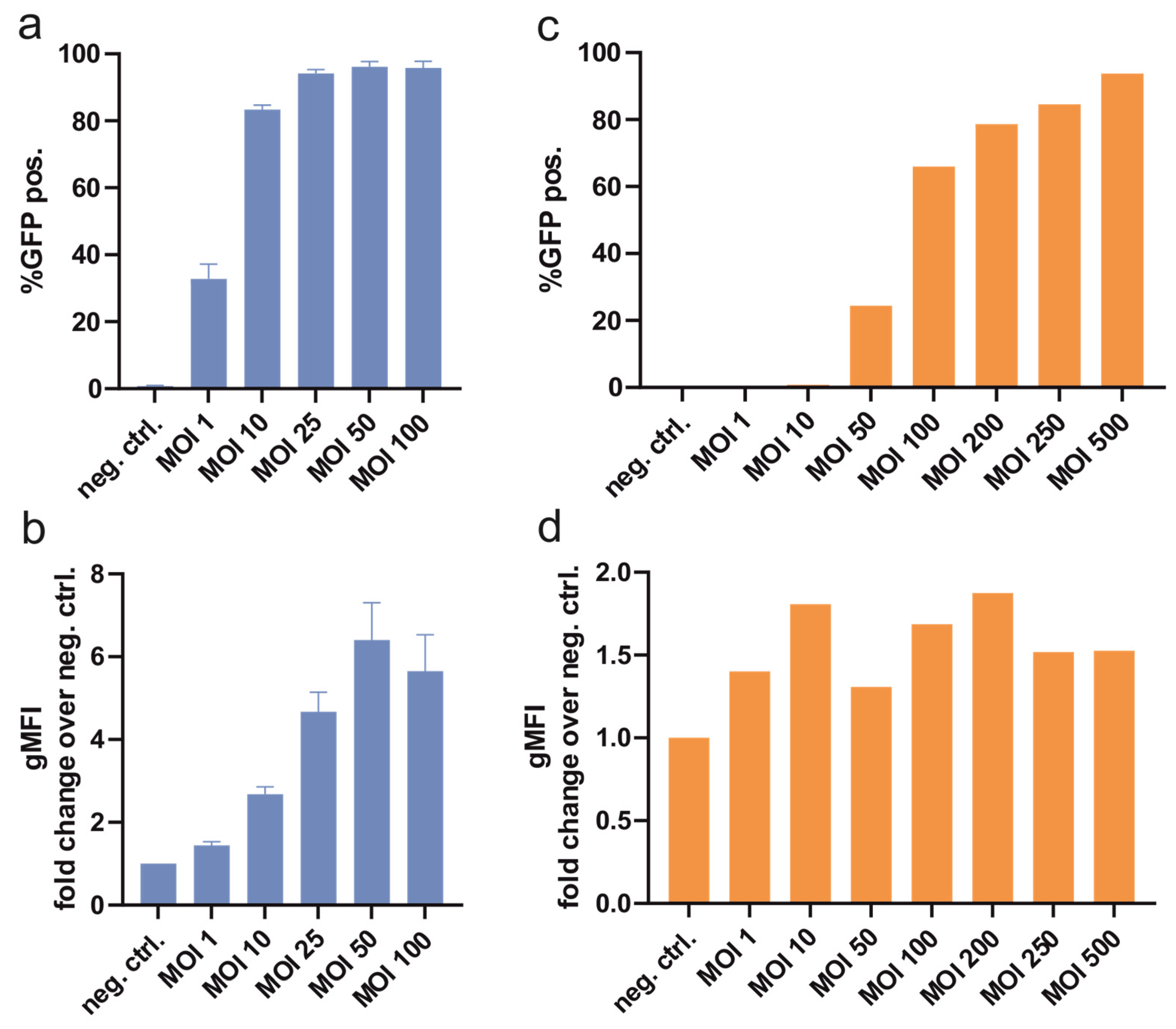
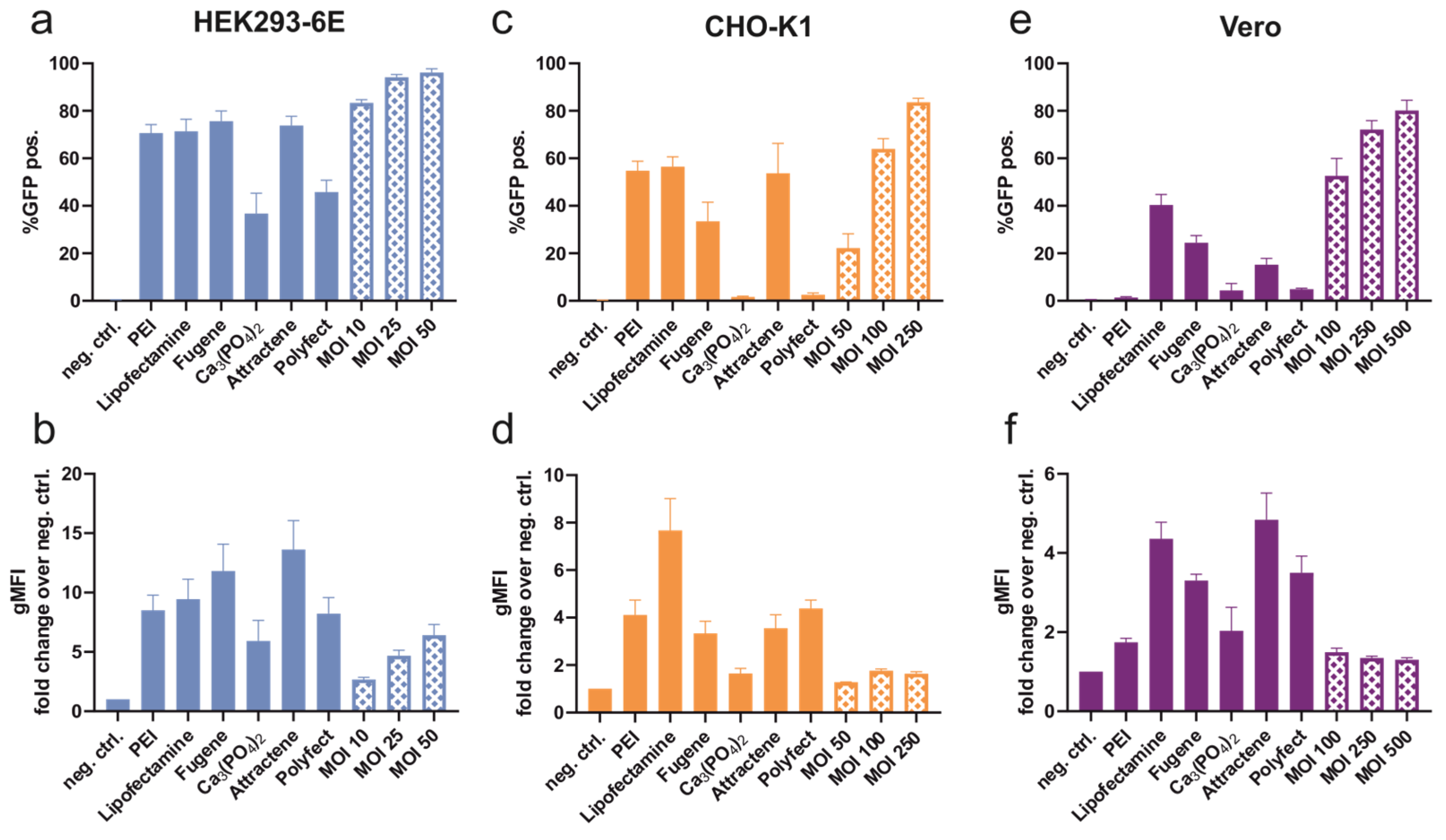
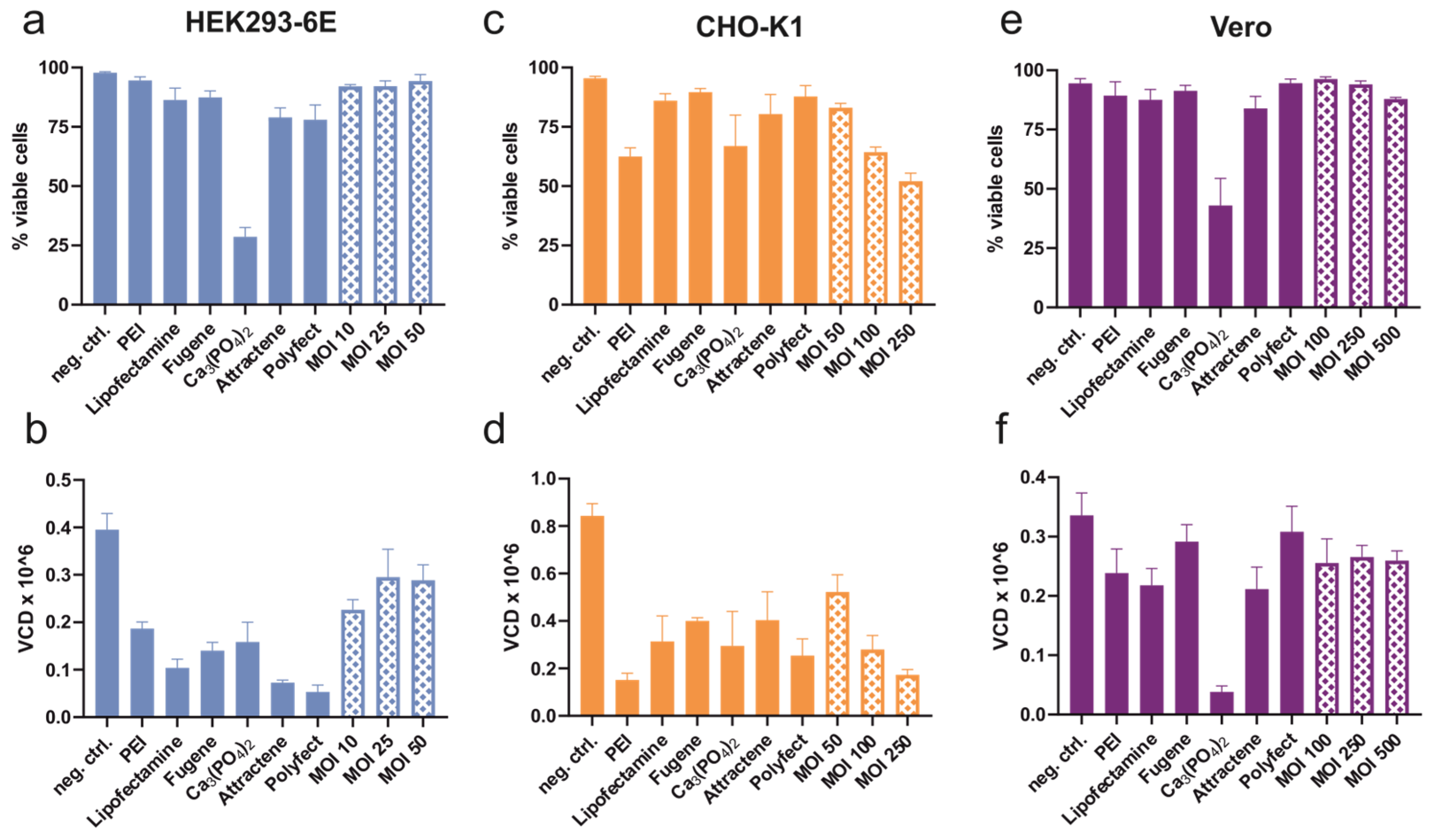
3.1. Transient Transfection and Transduction
3.1.1. MOI Titration
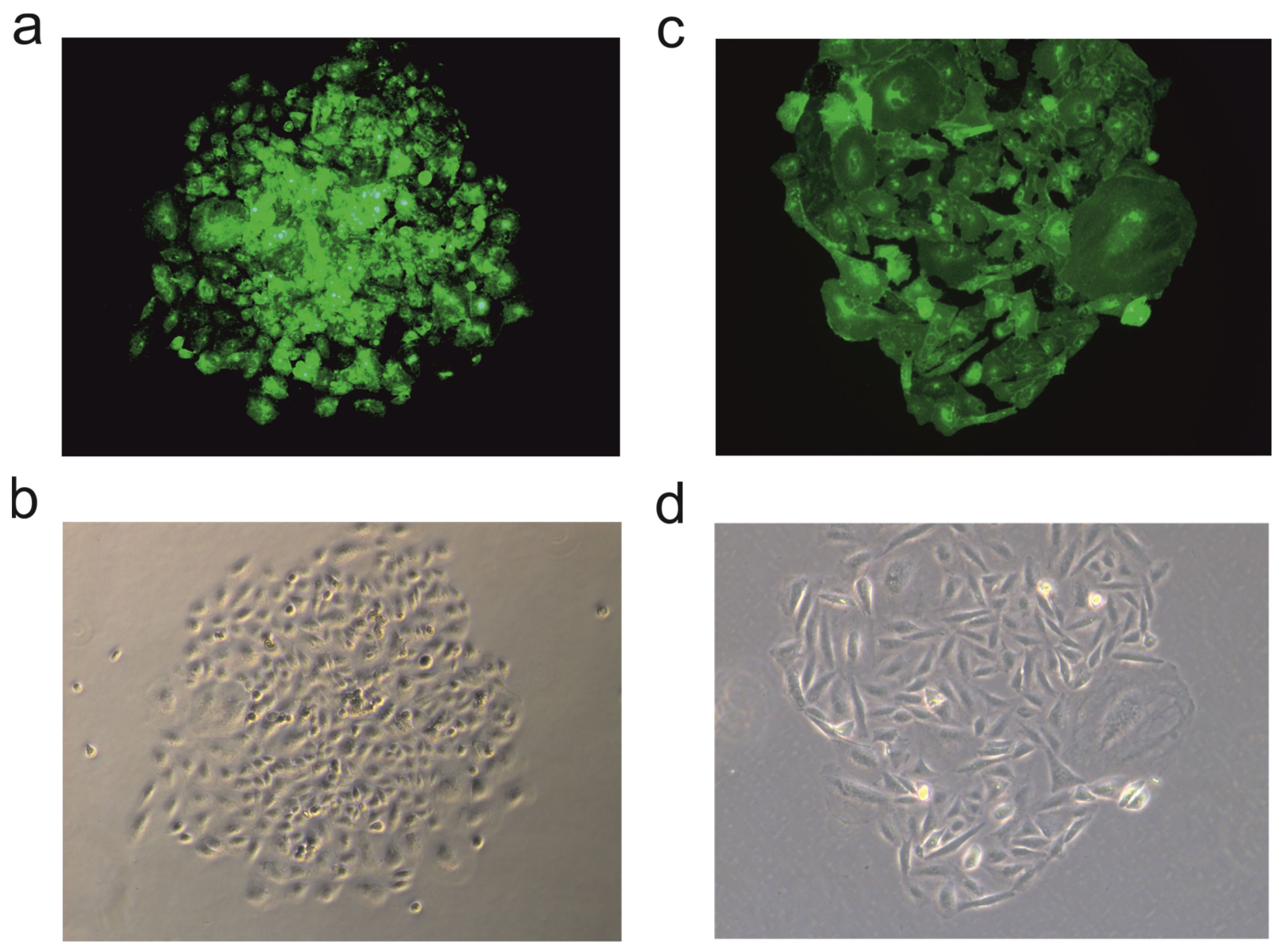
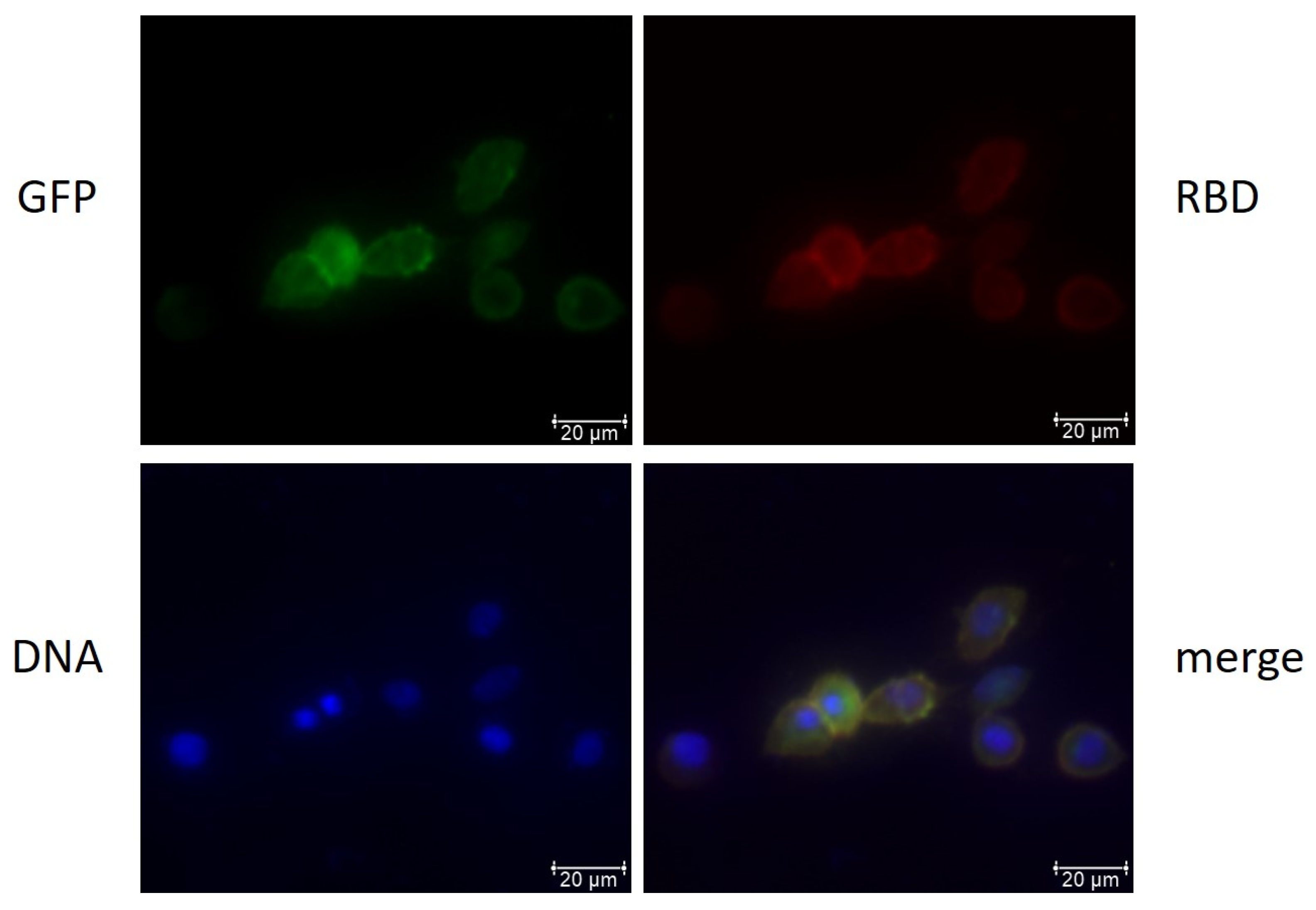
3.1.2. Recombinant Protein Expression after Transduction and Transfection
3.1.3. Cell Viability after Transduction and Transfection
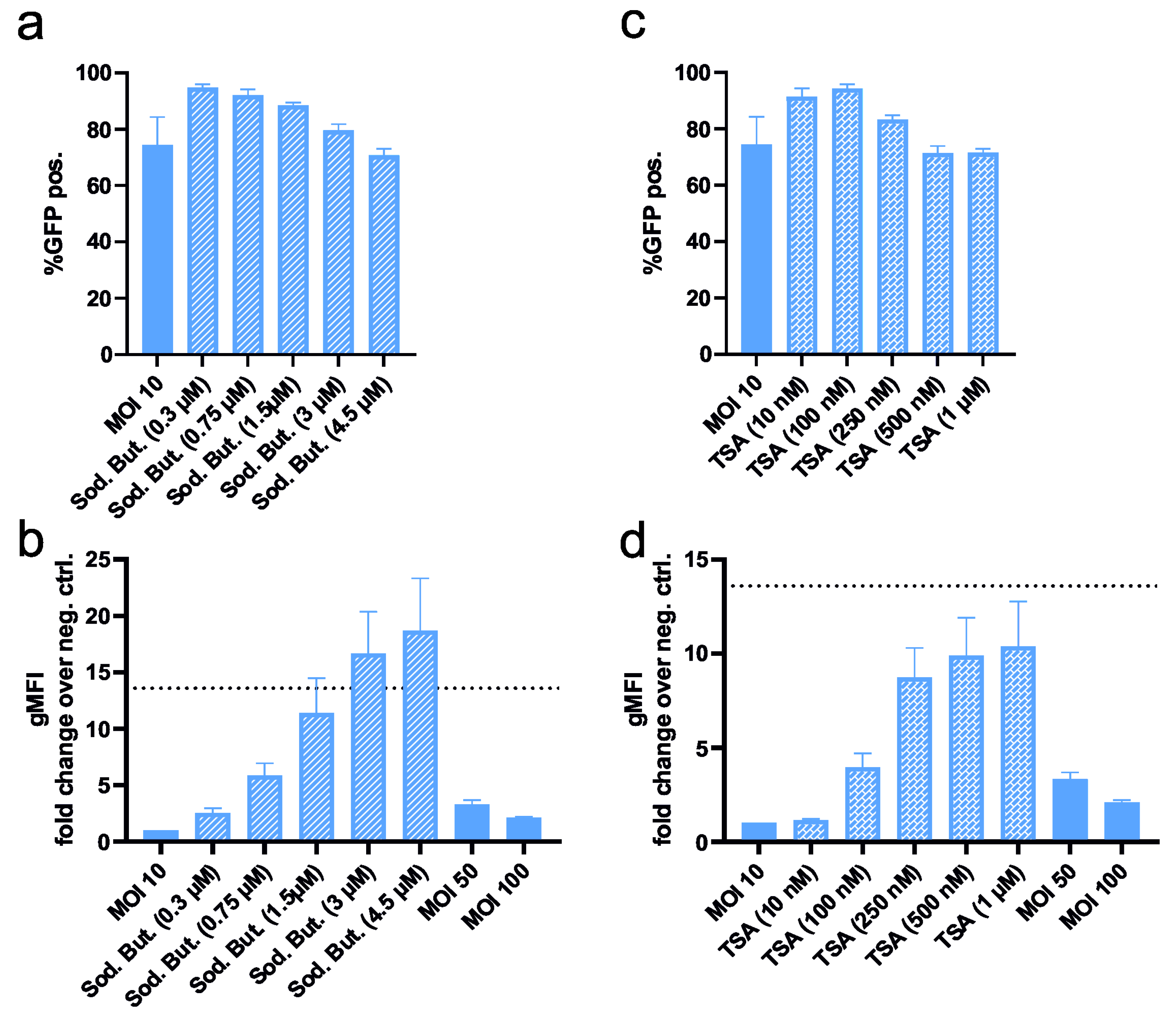
3.1.4. Enhancement of Transduced Protein Expression
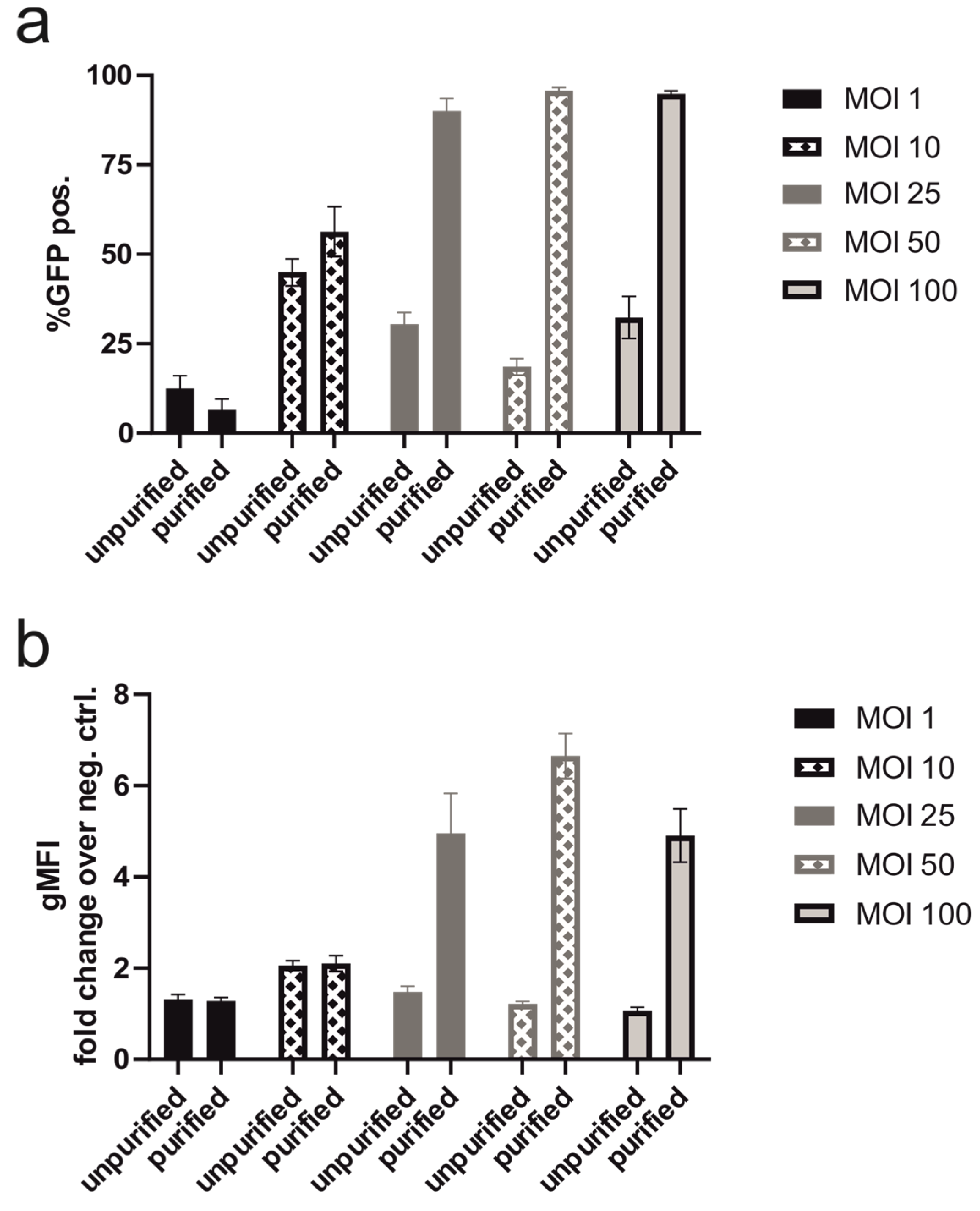
3.1.5. Transduction with Purified Baculovirus Produced on an Industrial Scale
3.2. Stable Transfection and Transduction
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A


References
- Recillas-Targa, F. Multiple Strategies for Gene Transfer, Expression, Knockdown, and Chromatin Influence in Mammalian Cell Lines and Transgenic Animals. Mol. Biotechnol. 2006, 34, 337–354. [Google Scholar] [CrossRef] [PubMed]
- Maurisse, R.; De Semir, D.; Emamekhoo, H.; Bedayat, B.; Abdolmohammadi, A.; Parsi, H.; Gruenert, D.C. Comparative Transfection of DNA into Primary and Transformed Mammalian Cells from Different Lineages. BMC Biotechnol. 2010, 10, 9. [Google Scholar] [CrossRef]
- Hashemi, A.; Roohvand, F.; Ghahremani, M.H.; Aghasadeghi, M.R.; Vahabpour, R.; Motevali, F.; Memarnejadian, A. Optimization of Transfection Methods for Huh-7 and Vero Cells: A Comparative Study. Cytol. Genet. 2012, 46, 347–353. [Google Scholar] [CrossRef]
- Graves, L.P.; Aksular, M.; Alakeely, R.A.; Buck, D.R.; Chambers, A.C.; Murguia-Meca, F.; Plata-Muñoz, J.J.; Hughes, S.; Johnson, P.R.V.; Possee, R.D.; et al. Improved Baculovirus Vectors for Transduction and Gene Expression in Human Pancreatic Islet Cells. Viruses 2018, 10, 574. [Google Scholar] [CrossRef] [PubMed]
- Puente-Massaguer, E.; Cajamarca-Berrezueta, B.; Volart, A.; González-Domínguez, I.; Gòdia, F. Transduction of HEK293 Cells with BacMam Baculovirus Is an Efficient System for the Production of HIV-1 Virus-like Particles. Viruses 2022, 14, 636. [Google Scholar] [CrossRef]
- Hu, Y.C. Baculovirus as a Highly Efficient Expression Vector in Insect and Mammalian Cells. Acta Pharmacol. Sin. 2005, 26, 405–416. [Google Scholar] [CrossRef]
- van Oers, M.M. Vaccines for Viral and Parasitic Diseases Produced with Baculovirus Vectors. Adv. Virus Res. 2006, 68, 193–253. [Google Scholar] [CrossRef]
- Felberbaum, R.S. The Baculovirus Expression Vector System: A Commercial Manufacturing Platform for Viral Vaccines and Gene Therapy Vectors. Biotechnol. J. 2015, 10, 702–714. [Google Scholar] [CrossRef]
- Gupta, K.; Tölzer, C.; Sari-Ak, D.; Fitzgerald, D.J.; Schaffitzel, C.; Berger, I. Multibac: Baculovirus-Mediated Multigene DNA Cargo Delivery in Insect and Mammalian Cells. Viruses 2019, 11, 198. [Google Scholar] [CrossRef] [PubMed]
- Hong, Q.; Liu, J.; Wei, Y.; Wei, X. Application of Baculovirus Expression Vector System (BEVS) in Vaccine Development. Vaccines 2023, 11, 1218. [Google Scholar] [CrossRef] [PubMed]
- Harrison, R.L.; Herniou, E.A.; Jehle, J.A.; Theilmann, D.A.; Burand, J.P.; Becnel, J.J.; Krell, P.J.; van Oers, M.M.; Mowery, J.D.; Bauchan, G.R. ICTV Virus Taxonomy Profile: Baculoviridae. J. Gen. Virol. 2018, 99, 1185–1186. [Google Scholar] [CrossRef]
- Volkman, L.E.; Goldsmith, P.A. In Vitro Survey of Autographa Californica Nuclear Polyhedrosis Virus Interaction with Nontarget Vertebrate Host Cells. Appl. Environ. Microbiol. 1983, 45, 1085–1093. [Google Scholar] [CrossRef]
- Kost, T.A.; Patrick, C.J.; Mickelson, C.A. Biosafety and Viral Gene Transfer Vectors. In Biological Safety; ASM Press: Washington, DC, USA, 2006; pp. 509–530. ISBN 9781555819248. [Google Scholar]
- Boyce, F.M.; Bucher, N.L. Baculovirus-Mediated Gene Transfer into Mammalian Cells. Proc. Natl. Acad. Sci. USA 1996, 93, 2348–2352. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, C.; Sandig, V.; Jennings, G.; Rudolph, M.; Schlag, P.; Strauss, M. Efficient Gene Transfer into Human Hepatocytes by Baculovirus Vectors. Proc. Natl. Acad. Sci. USA 1995, 92, 10099–10103. [Google Scholar] [CrossRef] [PubMed]
- Shoji, I.; Aizaki, H.; Tani, H.; Ishii, K.; Chiba, T.; Saito, I.; Miyamura, T.; Matsuura, Y. Efficient Gene Transfer into Various Mammalian Cells, Including Non-Hepatic Cells, by Baculovirus Vectors. J. Gen. Virol. 1997, 78, 2657–2664. [Google Scholar] [CrossRef] [PubMed]
- Berger, I.; Fitzgerald, D.J.; Richmond, T.J. Baculovirus Expression System for Heterologous Multiprotein Complexes. Nat. Biotechnol. 2004, 22, 1583–1587. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, D.J.; Berger, P.; Schaffitzel, C.; Yamada, K.; Richmond, T.J.; Berger, I. Protein Complex Expression by Using Multigene Baculoviral Vectors. Nat. Methods 2006, 3, 1021–1032. [Google Scholar] [CrossRef] [PubMed]
- Bieniossek, C.; Richmond, T.J.; Berger, I. MultiBac: Multigene Baculovirus-Based Eukaryotic Protein Complex Production. Curr. Protoc. Protein Sci. 2008, 51, 5.20.1–5.20.26. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.-Y.; Chen, G.-Y.; Hu, Y.-C. Recent Patents on the Baculovirus Systems. Recent Pat. Biotechnol. 2011, 5, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mansouri, M.; Berger, P. Baculovirus for Gene Delivery to Mammalian Cells: Past, Present and Future. Plasmid 2018, 98, 1–7. [Google Scholar] [CrossRef]
- Dressel, U.; Bailey, P.J.; Wang, S.-C.M.; Downes, M.; Evans, R.M.; Muscat, G.E.O. A Dynamic Role for HDAC7 in MEF2-Mediated Muscle Differentiation. J. Biol. Chem. 2001, 276, 17007–17013. [Google Scholar] [CrossRef]
- Hebbes, T.R.; Thorne, A.W.; Crane-Robinson, C. A Direct Link between Core Histone Acetylation and Transcriptionally Active Chromatin. EMBO J. 1988, 7, 1395–1402. [Google Scholar] [CrossRef] [PubMed]
- Goff, S.P. Silencing of Unintegrated Retroviral DNAs. Viruses 2021, 13, 2248. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, A.; Edelstein, H.I.; Glykofrydis, F.; Love, K.S.; Palacios, S.; Tycko, J.; Zhang, M.; Lensch, S.; Shields, C.E.; Livingston, M.; et al. The Sound of Silence: Transgene Silencing in Mammalian Cell Engineering. Cell Syst. 2022, 13, 950–973. [Google Scholar] [CrossRef] [PubMed]
- Spenger, A.; Ernst, W.; Condreay, J.P.; Kost, T.A.; Grabherr, R. Influence of Promoter Choice and Trichostatin a Treatment on Expression of Baculovirus Delivered Genes in Mammalian Cells. Protein Expr. Purif. 2004, 38, 17–23. [Google Scholar] [CrossRef]
- Condreay, J.P.; Witherspoon, S.M.; Clay, W.C.; Kost, T.A. Transient and Stable Gene Expression in Mammalian Cells Transduced with a Recombinant Baculovirus Vector. Proc. Natl. Acad. Sci. USA 1999, 96, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Dukkipati, A.; Park, H.H.; Waghray, D.; Fischer, S.; Garcia, K.C. BacMam System for High-Level Expression of Recombinant Soluble and Membrane Glycoproteins for Structural Studies. Protein Expr. Purif. 2008, 62, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Goehring, A.; Lee, C.H.; Wang, K.H.; Michel, J.C.; Claxton, D.P.; Baconguis, I.; Althoff, T.; Fischer, S.; Garcia, K.C.; Gouaux, E. Screening and Large-Scale Expression of Membrane Proteins in Mammalian Cells for Structural Studies. Nat. Protoc. 2014, 9, 2574–2585. [Google Scholar] [CrossRef]
- Bartolomé, A.; Liang, J.; Wang, P.; Ho, D.D.; Pajvani, U.B. Angiotensin Converting Enzyme 2 Is a Novel Target of the γ-Secretase Complex. Sci. Rep. 2021, 11, 9803. [Google Scholar] [CrossRef]
- Dutra, G.; Preinsperger, S.; Reithofer, M.; Grabherr, R.; Aguilar, P. Comparison between Methods for the Fast Purification of Baculovirus for Mammalian Cell Transduction. 2024; Manuscript in preparation. [Google Scholar]
- Liang, C.Y.; Chen, X.W. Baculoviruses as Vectors in Mammalian Cells. Virol. Sin. 2007, 22, 148–157. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′-3′) |
|---|---|
| 23.29 ACE2_GFP golden gate FWD | ggctaccacctgctgacattgattattgactagttattaatagtaatc |
| 23.30 ACE2_GFP golden gate REV | ggctaccacctgcgggctggataagatacattgatgagtttg |
| 23.31 pACEBac1 golden gate FWD | ggctaccacctgcgggctccactgcttgagcctagaag |
| 23.32 pACEBac1 golden gate REV | ggctaccacctgctgaccaatgtcatactagtgtttaaactcgctac |
| Cell Line | PEI | Lipofectamine | Fugene | Calcium Phosphate | Attractene | Polyfect | Baculovirus |
|---|---|---|---|---|---|---|---|
| HEK293-6E | 0/3 | 0/3 | 0/3 | 0/3 | 1/3 | 0/3 | 3/3 |
| CHO-K1 | 2/5 | 2/5 | 1/5 | 0/5 | 2/5 | 0/5 | 3/3 |
| Vero | 0/4 | 0/4 | 0/4 | 0/4 | 0/4 | 0/4 | n.d. 1 |
| Baculovirus Transduction | Conventional Transfection | |
|---|---|---|
| Time to obtain recombinant constructs | 1–2 weeks | 1–2 weeks |
| Time for gene transfer agent preparation | propagation of virus 3 weeks | big amount of plasmid up to 1 week |
| Storage conditions | 4 °C | −20 °C |
| Cost of cultivation of bacteria or insect cells | low to medium | low |
| BSL1 cell culture facility | yes | yes |
| Delivery efficiency (various cell lines) | high | low to high |
| Large cargo capacity | yes | no |
| Protein yield | comparable | |
| Stable cell line development | yes (higher probability) | yes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toth, M.; Reithofer, M.; Dutra, G.; Pereira Aguilar, P.; Dürauer, A.; Grabherr, R. Comprehensive Comparison of Baculoviral and Plasmid Gene Delivery in Mammalian Cells. Viruses 2024, 16, 426. https://doi.org/10.3390/v16030426
Toth M, Reithofer M, Dutra G, Pereira Aguilar P, Dürauer A, Grabherr R. Comprehensive Comparison of Baculoviral and Plasmid Gene Delivery in Mammalian Cells. Viruses. 2024; 16(3):426. https://doi.org/10.3390/v16030426
Chicago/Turabian StyleToth, Maria, Manuel Reithofer, Gregory Dutra, Patricia Pereira Aguilar, Astrid Dürauer, and Reingard Grabherr. 2024. "Comprehensive Comparison of Baculoviral and Plasmid Gene Delivery in Mammalian Cells" Viruses 16, no. 3: 426. https://doi.org/10.3390/v16030426