
The History of Baculovirology in Africa

1
Citrus Research International, P.O. Box 5095, Walmer, Gqeberha 6065, South Africa
2
Centre for Biological Control, Department of Zoology and Entomology, Rhodes University, P.O. Box 94, Makhanda 6140, South Africa
3
Department of Biochemistry and Microbiology, Rhodes University, P.O. Box 94, Makhanda 6140, South Africa
*
Author to whom correspondence should be addressed.
Viruses 2023, 15(7), 1519; https://doi.org/10.3390/v15071519
Submission received: 22 June 2023
/
Accepted: 4 July 2023
/
Published: 7 July 2023
(This article belongs to the Special Issue Applications of Baculoviruses: Expression Factories, Vaccines and VLPs, Gene Delivery Vectors, Biological Control and Virus Genetics Models)
Abstract
:Baculovirology has been studied on the African continent for the development of insect virus-based biopesticides and, to a much lesser extent, vaccine production and delivery, since the 1960s. In this review, we focus only on baculoviruses as biopesticides for agricultural pests in Africa. At least 11 species of baculovirus have been discovered or studied on the African continent, some with several distinct isolates, with the objective in most cases being the development of a biopesticide. These include the nucleopolyhedroviruses of Helicoverpa armigera, Cryptophlebia peltastica, Spodoptera exempta, Spodoptera frugiperda, Spodoptera littoralis, and Maruca vitrata, as well as the granuloviruses of Cydia pomonella, Plutella xylostella, Thaumatotibia (Cryptophlebia) leucotreta, Choristoneura occidentalis, and Phthorimaea operculella. Eleven different baculovirus-based biopesticides are recorded as being registered and commercially available on the African continent. Baculoviruses are recorded to have been isolated, researched, utilised in field trials, and/or commercially deployed as biopesticides in at least 13 different African countries. Baculovirus research is ongoing in Africa, and researchers are confident that further novel species and isolates will be discovered, to the benefit of environmentally responsible agricultural pest management, not only in Africa but also elsewhere.
1. Introduction
Baculoviridae is a family of double-stranded DNA (dsDNA) viruses with more than 90 species divided into four genera: the alpha-, beta-, delta-, and gammabaculoviruses [1,2]. Baculovirus species are classified according to a new binomial naming system, while the common names and abbreviations have not changed [3]. The genome comprises a single circular dsDNA molecule with a wide range of lengths. The smallest genomes are around 80 Kbp, as seen in Neodiprion lecontei nucleopolyhedrovirus (Gammabaculovirus nelecontei) at 81,755 bp and 89 open reading frames (ORFs) [2]. The largest genomes are around 180 Kbp, such as in Xestia c-nigrum granulovirus (Betabaculovirus xecnigri) at 178,733 bp containing 181 ORFs [4]. Baculoviridae viruses are characterised by the formation of occlusion bodies (OBs), with members falling into two morphological groupings. Nucleopolyhedroviruses (NPVs) form large OBs, referred to as polyhedra, which range from 0.6 to 2 µM. Granuloviruses (GVs) form smaller OBs ranging from 0.2 to 0.4 µM and are referred to as granules [5]. The formation of OBs enables baculoviruses to persist under normal environmental conditions for extended periods of time. The structural characteristics of baculoviruses have been extensively studied and are reviewed in detail in several publications (see, for example, [5,6]. Baculoviruses have a broad host range, infecting the larval stages of insects in the orders Hymenoptera, Diptera, and Lepidoptera [1].
The structural and biological characteristics of baculoviruses make them well suited for use as biological pesticides for the control of lepidopteran pests [7,8,9,10]. Over the past several decades, multiple baculovirus species have been tested, formulated, and sold as commercial biopesticides in various regions of the world. These regions primarily include South America, North America, Europe, and Asia [11,12,13,14]. As with elsewhere in the world, baculoviruses have been investigated as vaccine production and delivery systems in Africa. This specifically pertains to the development of a vaccine for the African horse sickness virus, using a baculovirus expression system [15,16,17,18]. However, challenges with production yields [19], the need for repeated boost inoculations [20], and the use of potent adjuvants to enhance immunogenicity [21] have limited the usefulness and application of this type of vaccine. Consequently, in this review, we chose to focus on the research and development of baculoviruses into biopesticides for the control of important pests across the African continent. At least 11 baculovirus species have been or are being investigated for use as biopesticides in African countries, as shown in Figure 1. These investigations are broad and include the isolation and identification of novel species, the evaluation of biological activity under laboratory conditions, the determination of efficacy under field conditions, and the formulation of baculoviruses as active ingredients in commercial products for large-scale agricultural applications.
2. Baculoviruses
2.1. Alphabaculovirus Helarmigerae, Helicoverpa armigera Nucleopolyhedrovirus (HearNPV)
Helicoverpa armigera (Hübner) (Noctuidae: Lepidoptera) has previously been ranked as the most important lepidopteran pest in South Africa [22,23]. Whether this is still so or not is questionable; nonetheless, it remains a very important and potentially destructive polyphagous pest on the continent. The first report of the occurrence of HearNPV in Africa was by Whitlock [24], who documented the symptomatology, not only of NPV but also a GV of the same insect, enabling the identification of the causative agent, based purely on symptoms of infection. However, the viruses were identified using electron microscopy. In laboratory bioassays, the LT50 with HearNPV was 5–8 days, whereas for HearGV, it was 17–25 days, depending on the concentration applied [25]. Furthermore, GV-infected larvae could continue feeding for up to 35 days, precluding any potential for use in a biopesticide [24]. Whitlock [26] also identified a degree of maturation immunity (from one instar to the next) to HearNPV, now a well-known phenomenon.
Roome [27] was the first to explore the efficacy of HearNPV in field trials. This he did against H. armigera on sorghum and cotton in Botswana. He found that a local isolate of HearNPV was as effective as the chemical insecticide, carbaryl, and more effective than commercial preparations of Helicoverpa zea NPV (HzNPV), Alphabaculovirus hezeae, and Bacillus thuringiensis var kurstaki (Btk), in reducing damage on both crops. The addition of molasses enabled a halving of the concentration of HearNPV applied, without any loss of efficacy. Contrary to expectations, spraying in the evening was no more effective than spraying in the day, possibly because the virus was protected against UV irradiation in the parts of plants where the larvae fed.
More recently, Moore et al. [28] conducted field trials against H. armigera on citrus in South Africa. Globally, Helicoverpa armigera is generally not recognised as a pest of citrus. However, in South Africa, the infestation of blossoms and setting fruitlets is an annual occurrence, often requiring spraying if the intervention threshold is surpassed [29]. Trials were initially conducted on potted tomato plants within a greenhouse, in order to obtain guidance on what concentrations to use in the field. In two field trials on Navel oranges, a concentration of 7.26 × 105 OBs/mL (and a couple of higher concentrations) resulted in a 100% reduction in H. armigera infestation within 14 days or longer. Damage to fruit was reduced by up to 84%, and rejection for export was reduced by up to 96%. These results were better than those achieved with Btk, Neem oil, and various organophosphates in the same trials. This work led to the development of the first commercially available HearNPV product in Africa, namely Helicovir (River Bioscience, Gqeberha, South Africa) [30]. Subsequently, three more products have been registered: two based on a different (non-African) isolate, Bolldex and Graboll (Andermatt Biocontrol, Grossdietwil, Switzerland), and one based on an isolate from Kenya, Helitec (Kenya Biologics, Runyenjes, Kenya) [31], initially registered in Ghana.
A few fundamental genetic and biological studies have been conducted on HearNPV in Africa. Through restriction endonuclease (REN) analysis, Ogembo [32] showed that two HearNPV isolates, one from South Africa, as originally reported by Whitlock [24], and one from Kenya, differed genetically. Ogembo et al. [33] went on to show in a dose–response study that both isolates had identical virulence to that of a commercial HzNPV product, Gemstar, against first- and second-instar H. armigera, but that the HearNPV isolates were more virulent than Gemstar against the third and fourth instars. Ultimately, the South African isolate was the most virulent, including having the shortest time to death in time–response assays. Ogembo et al. [34] expanded the study to include several additional isolates from Kenya and isolates from Zimbabwe and Thailand. Of the 162 clones isolated, 25 were shown to be unique, using REN. Finally, the virulence of several of the clones was compared, identifying the most promising candidate for biocontrol of H. armigera. Mtambanengwe [35] conducted full-genome sequencing with two novel isolates of HearNPV, collected from the Eastern Cape and KwaZulu-Natal provinces in South Africa, demonstrating their uniqueness in comparison with isolates described in studies from elsewhere.
Baillie and Bouwer [36] used next-generation sequencing (NGS) and denaturing gradient gel electrophoresis (DGGE) to identify the genetic differences between seven geographically distinct HearNPV isolates from South Africa. Depending on the gene, NGS identified between 31% and 35% of SNPs that were non-synonymous and may thus affect the biological function of the encoded proteins and therefore also the virulence of different virus isolates. Baillie and Bouwer [37] also found that inoculation dose with HearNPV against neonate H. armigera in bioassays significantly affected the genetic diversity of harvested virus thereafter.
Grant and Bouwer [38] demonstrated that not only differences in virulence occur between different HearNPV isolates but also different H. armigera populations respond differently to the same isolate. For example, although median lethal doses (LD50) did not differ significantly between two field-collected and one laboratory-reared population, the median lethal time (LT50) of the laboratory population was significantly shorter than the LT50 of the field populations. Bouwer et al. [39] also showed that the larvae surviving sublethal doses (LD25 an LD75) had significantly higher metabolic rates (respiration rates, measured as the rate of CO2 production) than that of untreated larvae. The authors postulated that this was due to a combination of viral replication, organ damage, and an energy-intensive induced cellular immune response.
2.2. Betabaculovirus Cypomonellae, Cydia pomonella Granulovirus (CpGV)
Cydia pomonella (Linnaeus) (Lepidoptera: Tortricidae), more commonly known as the codling moth, is a worldwide pest of apple, walnut, and pear. Biological control has long been implemented against C. pomonella, which includes entomopathogenic fungi, viruses, nematodes, bacteria, and microsporidia [40]. The betabaculovirus Cydia pomonella granulovirus (CpGV) is a highly efficacious and selective virus, which has been produced as a commercial product in Europe and North America and utilised globally for the control of C. pomonella since the early 1990s [41,42]. Two commercial products, Carpovirusine® (Arysta LifeScience, Noguères, France) and Madex® (Andermatt-Biocontrol AG, Grossdietwil, Switzerland), both of which are formulated with the Mexican CpGV isolate, are utilised in South Africa for the control of C. pomonella [43]. However, the continued use of CpGV in commercial products in Europe and the USA has led to the development of multiple types of resistance in C. pomonella populations in these regions [44,45,46,47,48,49,50]. In response, novel isolates have been continually formulated into new commercial products for use against these resistant populations.
While no resistance has yet been observed in South African populations of C. pomonella, a study was conducted by Motsoeneng et al. [51] to search for novel isolates that could be utilised should the need arise, leading to the isolation of CpGV-SA. The genetic analysis of CpGV-SA indicated this isolate to be unique from previously identified isolates, harbouring many unique single nucleotide polymorphisms (SNPs) [51]. Furthermore, the biological activity of CrleGV-SA and CpGV-M against C. pomonella was determined and compared. These isolates were found to have similar median lethal concentrations, LC50 and LC90 values at 1.6 × 103 and 1.2 × 105 OBs/mL for CpGV-SA and 3.1 × 103 and 2.8 × 105 OBs/mL for CpGV-M, respectively [52]. The LT50 for CpGV-SA and CpGV-M against C. pomonella were also determined to be 135 and 136 h, respectively, with each isolate applied at concentrations equal to their respective LC90 [52]. These results suggest that CpGV-SA could be developed into a commercial biopesticide and utilised as an alternative to or interchangeably with existing commercial biopesticides based on CpGV-M, both in South Africa and in other African countries where C. pomonella is problematic.
2.3. Betabaculovirus Pluxylostellae, Plutella xylostella Granulovirus (PlxyGV)
Plutella xylostella granulovirus (PlxyGV), a betabaculovirus that infects the diamondback moth, Plutella xylostella (L.) (Lepidoptera: Plutellidae), was first discovered in 1970 by Asayama and Osaki [53] from diseased larvae in Japan. Since then, additional isolates of PlxyGV were identified from several countries, including China, Taiwan, India, Malaysia, and of particular interest Kenya and South Africa. In field surveys conducted by Parnell et al. [54], 127 diseased P. xylostella cadavers collected from eight farms surrounding Nairobi, Kenya, were examined for the presence of PlxyGV. Of these, 95 cadavers were observed to be suffering from PlxyGV infections, from which 14 isolates with unique REN profile patterns were distinguished. Subsequently, laboratory and small-plot field trials were conducted by Grzywacz et al. [55] using several of these isolates, to evaluate their efficacy against P. xylostella. Dose–response bioassays in the laboratory demonstrated an LC50 ranging from 3.95 × 107 OBs/mL in the PlxyGV-Nya-40 isolate to 2.36 × 106 OBs/mL in the PlxyGV-Nya-01 isolate, compared with the LC50 of the Taiwanese reference isolate PlxyGV-Tw at 1.55 × 107 OBs/mL. Small-plot field trials were conducted using the PlxyGV-Nya-01 isolate on Kale, at both high (3.0 × 1014 OBs/ha) and low (3.0 × 1013 OBs/ha) application rates and compared against an unsprayed control and a chemical insecticide treatment [55]. Damage to crops by DBM was reduced below that seen in either the chemical insecticide treatment or untreated controls for both PlxyGV application rates.
Field surveys across 92 sites in Benin were conducted in 2001, in which more than 5000 larvae were sampled and screened for PlxyGV [56]. PlxyGV was not detected in any of these West African survey sites, resulting in the importation of the Kenyan isolate PlxyGV-Nya-01 into Benin for evaluation. Dose–response assays conducted using the Kenyan isolate against the Benin P. xylostella population resulted in an LC50 of 1.15 × 105 and 2.40 × 106 OBs/mL for the second and third instars, respectively. The LC50 was reported to be considerably lower in the Benin population than that measured against the Kenyan population, with Cherry et al. [56] hypothesising major differences in prior exposure to PlxyGV by these different geographic populations as a contributing factor, as evidenced by the complete lack of PlxyGV identified during these field surveys.
Abdulkadir et al. [57] isolated a South African isolate, PlxyGV-SA, from P. xylostella larvae collected from a field site in the Eastern Cape Province, South Africa. Surface dose bioassays of PlxyGV-SA against neonate P. xylostella larvae resulted in LC50 and LC90 values of 3.56 × 105 and 1.14 × 107 OBs/mL, respectively [58]. The LT50 for PlxyGV-SA was measured to be 5.3 days, which is comparable to the 4.9 days recorded for the PlxyGV-Tw isolate against neonate P. xylostella [58,59]. To date, the complete genome sequences of nine PlxyGV isolates have been determined and uploaded to GenBank (NCBI), of which PlxyGV-SA is the only African isolate [60]. The high degree of isolate variation identified in Kenya by Parnell et al. [54] led to the suggestion that there has been a long association between PlxyGV and DBM in Africa. With only a single African genome sequence currently available and a multitude of African isolates already identified, much of the genetic diversity among PlxyGV isolates in Africa remains untouched and unknown. Furthermore, PlxyGV is yet to be developed into a commercial biopesticide, likely due to the lack of an artificial diet, which is required for the mass rearing of P. xylostella larvae.
2.4. Betabaculovirus Cryleucotretae, Cryptophlebia leucotreta Granulovirus (CrleGV)
The false codling moth, Thaumatotibia leucotreta (Meyrick) (Lepidoptera: Tortricidae), is an important agricultural pest in Africa south of the Sahara [61,62]. Although it is capable of causing economic losses in citrus fruit (other than lemons and limes [63]), peaches, peppers, pomegranates, and some other crops, its main status is phytosanitary in nature, due to its endemism to Africa. A range of control options have been developed to manage the pest [64,65,66,67], one of which is the Cryptophlebia leucotreta granulovirus (CrleGV) [68,69,70]. The genus of the host was previously Cryptophlebia, hence the name of the virus, but the host name was changed to Thaumatotibia in the late 1990s [71].
The first discovery of CrleGV was from infected larvae found in the Ivory Coast in the 1960s [72]. It was found that if the CrleGV contamination of T. leucotreta larvae was not curtailed in a rearing facility, this could lead to the collapse of the culture [73]. Incidentally, the authors also noted a cypovirus (CPV) infection in the same laboratory culture. A second CrleGV isolate was reported in diseased larvae from Cape Verde [74]. The occurrence of a third isolate was reported from a laboratory culture of T. leucotreta in South Africa in 1980 [75]. Fritsch and Huber [76], Fritsch [77], and Jehle et al. [78] demonstrated, through REN analysis, that the three isolates were genetically distinct. Jehle et al. [78] constructed a restriction fragment map covering almost the entire genome of the Cape Verde isolate of CrleGV, identifying the position of the granulin gene through cross-hybridisation with granulin coding fragments of CpGV [79]. Moore [68] described the discovery and development of an additional novel CrleGV isolate, found infecting T. leucotreta larvae in a mass-rearing facility in the Western Cape Province of South Africa. This isolate was genetically characterised by Singh et al. [80]. Goble [81] further genetically characterised this same CrleGV isolate, comparing it with another South African isolate. Restriction analysis and partial amplifications of the granulin and ecdysteroid UDP-glucosyltransferase (egt) genes revealed 99% and 98% nucleotide identities, respectively, between the two isolates. Most recently, Opoku-Debrah et al. [82] succeeded in inducing outbreaks of CrleGV in five geographically distinct T. leucotreta laboratory cultures through the overcrowding of larvae. This led to the isolation and genetic characterisation of five novel South African CrleGV isolates. The single REN analysis of viral DNA and partial sequencing of granulin and egt genes and multiple alignments of nucleotide sequences were used to demonstrate these differences, leading to a proposal for two phylogenetic CrleGV-SA groups [82].
In addition to the genetic characterisation studies, extensive biological assays have been conducted. In dose–response assays with the Cape Verde CrleGV isolate (CrleGV-CV) and five CV clones, Fritsch [77] determined that the LC50 ranged from 9.38 × 103 to 1.86 × 107 OBs/mL. Moore et al. [70] reported surface inoculation dose–response, time–response, and detached fruit bioassays against T. leucotreta neonate larvae. LC50 and LC90 values were estimated as 4.09 × 103 and 1.18 × 105 OBs/mL, respectively. LT50 and LT90 values were estimated to be 4 days 22 h and 7 days 8 h, respectively. Opoku-Debrah et al. [83] tested the virulence of five geographically distinct and two commercial isolates of CrleGV against larvae from five T. leucotreta laboratory cultures of different regional origins, in laboratory bioassays. They demonstrated that virulence is a very specific relationship between host and pathogen, showing that certain isolates were significantly more or less virulent against certain regionally distinct host populations. Consequently, it could not be claimed that any particular isolate was more or less virulent, without contextualising it by stating against which population the comparison was conducted.
The first use of CrleGV for the control of T. leucotreta was in Cape Verde in the 1980s, where one small-scale field trial was reported on citrus and Spanish pepper [84]. Virus concentrations of 108 and 109 OBs/mL were used, applied with skimmed milk powder and a wetting agent. Damage by T. leucotreta was reduced by 77% in citrus and 65% in Spanish pepper. Moore [68], Moore et al. [69], Kirkman [85], and Moore et al. [86] reported more than 50 field trials with CrleGV against T. leucotreta on citrus in South Africa, spanning from 2000 to 2015, the earlier trials leading to the registration of the first commercially produced CrleGV product [69]. In a representative sample of 13 field trials, Moore et al. [86] reported reducing T. leucotreta infestation of citrus fruit by between 30% and 92%, in field trials with CrleGV. Efficacy was shown to persist at a level of 70% for up to 17 weeks after the application of CrleGV. The addition of molasses substantially and sometimes significantly enhanced efficacy. It was also established that CrleGV should not be applied at less than ~ 2 × 1013 OBs/ha in order to avoid compromised efficacy [86]. In 2003, the first CrleGV product was registered in South Africa for the control of T. leucotreta on citrus, avocadoes, grapes, and other crops, namely Cryptogran (River Bioscience, Gqeberha, South Africa) [69,87]. Subsequently, another two products have been registered and are commercially available for use on a range of susceptible crops, i.e., Cryptex and Gratham (both Andermatt Biocontrol, Grossdietwil, Switzerland) [88].
2.5. Alphabaculovirus Crypeltasicae, Cryptophlebia peltastica Nucleopolyhedrovirus (CrpeNPV)
Cryptophlebia peltastica (Meyrick) (Lepidoptera: Tortricidae), more commonly known as the litchi moth, is an important pest of litchi (Litchi chinensis Sonnerat) in several sub-Saharan and Indian Ocean countries such as Mauritius, South Africa, and Réunion Island [89,90,91,92]. Marsberg et al. [93] initiated a laboratory culture of litchi moth, maintained on T. leucotreta artificial diet [94]. Diseased larvae were collected and examined for the presence of baculovirus infection. This resulted in the discovery of a novel nucleopolyhedrovirus, which was subsequently genetically and biologically characterised. The virus, Cryptophlebia peltastica nucleopolyhedrovirus (CrpeNPV), was examined using transmission electron microscopy, which revealed the presence of polyhedral occlusion bodies with numerous singly enveloped nucleocapsids. The phylogenetic analysis of CrpeNPV based on late expression factor 8 (lef-8), lef-9, and polyhedrin sequence data confirmed the isolate to be a novel group two alphabaculovirus, with a genome length of 115,728 bp encompassing 126 ORFs (Marsberg et al. 2018). The biological activity of CrpeNPV in terms of median lethal concentration and time was evaluated against its homologous host C. peltastica. An LC50 and LC90 of 6.46 × 103 and 2.46 × 105 OBs/mL and LT50 and LT90 values of 76.32 and 93.49 h were measured, respectively [93].
As discussed above, C. pomonella is a serious pest worldwide. The development of several types of resistance toward existing biopesticides formulated with different CpGV isolates has created additional challenges for control strategies in Europe and the USA. A study by Wennmann et al. [95] examined the biological activity of CrpeNPV against several laboratory-reared strains of C. pomonella, each of which exhibited different types of resistance to CpGV infection. These included CpRR1, which exhibited type I resistance; CpR5M, which exhibited type II resistance; CpRGO, which exhibited type III resistance; and a susceptible strain CpS. Each resistance type reduces C. pomonella susceptibility to certain CpGV isolates, such as in CpRR1, which exhibits reduced susceptibility to CpGV-M, or CpRGO, which exhibits reduced susceptibility to CpGV-M and CpGV-S [44,49,50]. Each population was tested against CrpeNPV and two CpGV isolates, CpGV-M and CpGV-E2, the latter of which remains infectious to all these C. pomonella strains. CrpeNPV was shown to be highly infectious to both susceptible and CpGV-resistant C. pomonella strains, with biological activity similar to the resistance-breaking isolate CpGV-E2. CrpeNPV is currently undergoing registration as a biopesticide for use in Europe for the control of C. pomonella and Grapholita molesta (Busck) (Lepidoptera: Tortricidae) on pome fruit and stone fruit. It was also recently registered in South Africa for use against C. pomonella on pome fruit, T. leucotreta and T. batrachopa on macadamias, and T. leucotreta and C. peltastica on litchis (Sean Thackeray, Personal communication [96]).
2.6. Alphabaculovirus Spexemptae, Spodoptera exempta Nucleopolyhedrovirus (SpexNPV)
The larval stage of the African armyworm moth, Spodoptera exempta (Walker) (Lepidoptera: Noctuidae), is regarded as a devastating cyclical and migratory crop pest of maize, wheat, sorghum, and other staple crops in a large portion of sub-Saharan Africa [97]. The first finding of an NPV disease in the larvae of S. exempta was attributed to Graham [98] in Brown and Swaine [99]. The latter reported surveys and laboratory studies with the virus, concluding that its occurrence was widespread in field populations of S. exempta, particularly where there were pest outbreaks [99]. Odindo [100] concluded that epizootics in Kenya were highest in sites of high larval density, wide fluctuations in daily temperatures, and high humidity. In sites of high incidence, the epizootic led to larval population collapse within 3 weeks of the outbreak. Vilaplana et al. [101] examined the prevalence of covert infection of NPV in field populations of S. exempta, in Tanzania. They found that virtually all the insects collected in the field were positive for SpexNPV DNA, and 60% of these insects had transcriptionally active viruses. This was so, even in the absence of pest outbreaks, suggesting that SpexNPV is not only transmitted horizontally, but also vertically, at extremely high levels in field populations of S. exempta and can maintain a persistent infection without obvious symptoms. This was supported by the finding that similarly high levels of virus DNA and RNA were detected in an S. exempta culture that had been maintained in the laboratory for 5 years [101]. Subsequently, Graham et al. [102] developed a real-time quantitative polymerase chain reaction (qPCR) procedure for the specific detection and quantification of SpexNPV. They used this assay for the quantification of covert virus infection in asymptomatic larvae, concluding that viral load peaked in early instars (up to 6 days post-hatch), and decreased markedly with larval age, particularly during the final two instars.
Brown et al. [103] were the first to provide a physical map of SpexNPV, constructed using a range of restriction enzymes, and it showed a genome size of 131.89 Kbp. More recently, Redman et al. [104] cloned an isolate of SpexNPV in vivo, identifying at least 17 genetically distinct genotypes, which varied in size from approximately 115 to 153 Kbp. Subsequently, Escasa et al. [105] reported fully sequencing the genome of a SpexNPV isolate, collected in Tanzania in the 1970s, and thus very likely the same isolate as investigated by Redman et al. [104]. The genome was 129.5 Kbp.
Harrap et al. [106] established a laboratory culture of S. exempta, field-collected in Kenya, from which an NPV infection was isolated. Subsequently, Cherry et al. [107] developed and optimised an in vivo production system for SpexNPV. The larvae of S. exempta were reared in individual cells (17 × 17 × 40 mm) on an artificial diet that had been sprayed with a range of concentrations. The maximum yield obtained per larva was 3.29 × 109 OB per larva after 7 days of incubation. LT50 values fell with an increase in dose, being as short as 5.5 days when using 1 × 107 OB/cell. A virus production protocol was refined and prescribed in detail by Mushobozi et al. [108], including guidelines for the formulation and application of the virus preparation.
McKinley et al. [109] found that this virus was cross-infectious with the fall armyworm, S. frugiperda (Smith) (Lepidoptera: Noctuidae). However, rather than being directly pathogenic to the heterologous host, it appeared to trigger the homologous virus through stress induction. Odindo [110], through a series of dose–response and time–response bioassays with third-instar S. exempta and its virus, established the LD50 as 48 OBs per larva and the LT50 to range from 146 to 221 h, depending on the dosage applied.
A project managed by the Natural Resources Institute in the UK was launched in 1996 to develop and evaluate the use of SpexNPV as a biopesticide alternative to chemical control [111]. The project ran for at least a decade [112]. Field trials demonstrated that both the ground and aerial application of SpexNPV to armyworm outbreaks on pasture could initiate virus epizootics and population collapse [113,114]. SpexNPV was effective when applied at 1 x 1012 OB/ha if applied early during an outbreak and before larvae reached the fourth instar. Mass mortality was apparent 3–10 days after application. Efficacy in field trials in northern Tanzania, with doses as low as 1 x 1011 OB/ha, ranged from 22% to around 90% [115]. These differences in efficacy were attributed to the presence or absence of Wolbachia, which was related to an increase in the virulence of SpexNPV, contrary to most other findings with Wolbachia [112]. It was concluded that SpexNPV was sufficiently effective to replace a chemical pesticide strategy for controlling S. exempta [114]. Consequently, a large biopesticide production facility for SpexNPV was built in Arusha, Tanzania, from 2008 to 2011 [116]. Despite this, the field production of the virus was conducted instead, based on the Brazilian system for production of the Anticarsia gemmatalis NPV (AgNPV), Alphabaculovirus angemmatalis [117,118]. This was deemed technically feasible because of the large synchronous outbreaks of larvae at high densities that are a feature of this pest [113,114]. Unfortunately, the facility never became fully operational, despite the compelling arguments made in favour of the programme [119]. This was partly due to a subsequent decline in the pest status of S. exempta and the arrival of S. frugiperda [120], which superseded the latter in research and management efforts.
2.7. Alphabaculovirus Spofrugiperdae, Spodoptera frugiperda Nucleopolyhedrovirus (SfNPV)
Spodoptera frugiperda is native to tropical and subtropical regions of the American continents, stretching from Argentina in the south to southern Florida and Texas in the north [121,122]. It was first reported as an invader in Africa in 2016, appearing in Nigeria, Sao Tomé, Benin, and Togo [123]. It was most recently confirmed to be in more than 30 African countries, including as far south as South Africa [124]. Spodoptera frugiperda is highly polyphagous [125,126] but is mainly a pest of maize, followed by sorghum, cotton, sweet corn, and sugarcane [127]. It is considered a devastating pest throughout much of Africa, causing estimated yield losses in maize on the continent of USD 9.4 billion [128]. Despite this, surprisingly little research has been conducted on the Spodoptera frugiperda nucleopolyhedrovirus (SfNPV) on the continent, possibly because a notable body of research from elsewhere already exists (e.g., see [129,130,131,132,133,134,135]). Consequently, a commercial preparation of SfNPV, Fawligen (AgBiTech) was relatively rapidly registered in Africa, initially in Kenya [127]. An SfNPV isolate has also now been reported from Africa (Nigeria) and has been genetically characterised [136], showing a close relatedness to isolates described from the Americas, particularly from Brazil [137].
2.8. Alphabaculovirus Spolittoralis, Spodoptera littoralis Nucleopolyhedrovirus (SpliNPV)
Spodoptera littoralis (Boisd) (Lepidoptera: Noctuidae), the Egyptian cotton leafworm, is a polyphagous insect of important crops in Africa, Mediterranean Europe, and countries in the Middle East [138,139]. The baculovirus-based control of S. littoralis has a long history in Africa, with early field trials and evaluations dating back to the 1980s [140,141]. Field trials of Spodoptera littoralis nucleopolyhedrovirus (SpliNPV) were conducted by Topper et al. [140] against S. littoralis in Egypt, to compare formulations, spraying techniques, and virus efficacy. Although no difference was found between the spray technique and formulation, a significant reduction in leaf damage was measured in the SpliNPV treatments compared with an unsprayed control application. Large-scale field trials further showed decreased leaf damage for the SpliNPV treatment as compared to the control; however, no difference in cotton yield was observed. This lack of difference was attributed to low levels of damage, with virus treatments only applied to fields that exceeded a predetermined egg mass per ha threshold.
In field trials conducted in 1988 and 1990, Jones et al. [141] evaluated the use of a wettable powder formulation of SpliNPV for the control of S. littoralis. Three different virus concentrations were applied and compared against the traditional method of control, which involved the collection of egg masses by hand. The analysis of the 1988 field trial showed a significant reduction in mean leaf damage 14 days after treatment for both the egg collection and the two highest NPV concentrations as compared to the control. The NPV treatments further showed a significant reduction in mean leaf damage 14 days after spraying as compared to the control and egg collection treatments in the 1990 field trials. The highest NPV concentration maintained a significant reduction in leaf damage 21 days after application. In both the 1988 and 1990 field trials, a clear dose-dependent response was observed, with higher doses correlating with reduced leaf damage.
More recently, a study by Adel-Satter et al. [142] evaluated the resistance to different pyrethroids and biocides in several geographically distinct strains of S. littoralis, collected from isolated field populations in Egypt across three cotton seasons (2006, 2007, and 2008). Included among the biocides tested were several formulations of an NPV that infects S. littoralis (presumably SpliNPV). The results obtained showed that all field strains were highly susceptible to the NPV biocide, while high levels of resistance to pyrethroid insecticides were observed. The results from this study strongly support the use of NPV-based biocides for the control of S. littoralis in Egyptian cotton fields, either as a replacement to conventional insecticides or for strategic use in integrated pest management (IPM) programmes. Furthermore, research by Alfy et al. [143] investigated interactions between Bacillus thuringiensis subsp. mexicanensis (Btm) on SpliNPV, applied against S. littoralis. The virus was isolated from larvae collected at the Nubaria Agricultural Research Station, Egypt, with semi-field trials conducted in 2017 and 2018. The control of S. littoralis by SpliNPV alone and in combination with Btm was evaluated. Synergistic interactions were observed, with the combination of Btm and SpliNPV providing the highest measurement of S. littoralis larval mortality. Although a commercial bioinsecticide using SpliNPV has been formulated and sold under the name Littovir (Andermatt Biocontrol, Switzerland) in several European countries, the virus is yet to be developed into a product for use in African countries.
2.9. Betabaculovirus Choccidentalis, Choristoneura occidentalis Granulovirus (ChocGV)
The citrus leafroller, Choristoneura occidentalis (Walsingham) (Lepidoptera: Tortricidae), is native to South Africa but more widely distributed throughout sub-Saharan Africa, with records from Gambia, Kenya, Mozambique, Zimbabwe, Sierra Leone, Tanzania, and Zambia [144]. In South Africa, it is recognised as a pest of citrus, avocado, coffee, and various ornamentals [145,146]. It should not be confused with the North American species of the same name, C. occidentalis (Freeman). The citrus leafroller was moved from the genus Archips to Choristoneura by Razowski [147]. However, based on a phylogenetic study, Fagua et al. [148] recommended its transfer to the genus Cacoecimorpha or possibly even a reversion to Archips (Giovanny Fagua, Personal communication [149]).
Smith and Graça [150] reported infected C. occidentalis larvae from citrus in Eswatini (Swaziland). Upon electron microscopic inspection, the disease was determined to be caused by a granulovirus. However, the causative organism was only morphologically characterised, and it was therefore not determined whether this was a novel granulovirus and what its relatedness is to the more recently described Choristoneura occidentalis granulovirus (ChocGV), characterised and described by Escasa et al. [151] from the North American host species. Due to the disparity in host nomenclature, it is highly likely that these are two distinct baculovirus species and that the name for this African species should thus not be ChocGV. This must be resolved. No further work has been reported, and consequently, its potential for exploitation as a control agent against C. occidentalis must still be determined.
2.10. Alphabaculovirus Mavitratae, Maruca vitrata Multi-Nucleopolyhedrovirus (MaviMNPV)
The legume pod borer, Maruca vitrata (Fabricius) (Lepidoptera: Crambidae), is an important pest of cowpea (Vigna unguiculata (L.) Walp. (Fabales: Fabaceae) in West Africa. Srinivasan et al. [152] evaluated the efficacy of a Taiwanese isolate of Maruca vitrata multi-nucleopolyhedrovirus (MaviMNPV) against M. vitrata in semi-field caged experiments at IITA, in Benin. Maruca vitrata larval mortality increased by 78.8% following the application of MaviMNPV at a concentration of 2 × 1013 OBs/ha, 7 days after spraying, as compared to an untreated control application. The authors also noted the detection of M. vitrata larvae with apparent MaviMNPV symptomatology at a site 150 km from where the semi-field trials were conducted. MaviMNPV had not previously been detected or released into the open in Benin. Srinivasan et al. [152] evaluated whether the M. vitrata parasitoid, Apanteles taragamae Viereck (Hymenoptera: Braconidae), was involved in the transmission of MaviMNPV. It was found that the partial or complete body contamination of A. taragamae with MaviMNPV resulted in the transmission of the virus to M. vitrata, as compared to control treatments where the parasitoid was not contaminated.
Laboratory and field trials were further conducted by Sokame et al. [153] using the same Taiwanese isolate of MaviMNPV in Benin. Experiments evaluated the effect of combining the virus with different botanical oils against M. vitrata. The laboratory results showed a significant decrease in the mean survival time (MST) of M. vitrata when MaviMNPV was applied alone, decreasing from 14+ days in the control to 8.0 ± 2.1 and 5.0 ± 0.8 days at concentrations of 3 × 104 and 5 × 106 OBs/mL, respectively. The MST was further decreased to 2.0 ± 0.2 days when MaviMNPV (at a concentration of 5 × 106 OBs/mL) was combined with Azadirachta indica (neem) oil at 0.5%. A similar synergistic interaction was observed when evaluating the percentage mortality induced by MaviMNPV, increasing from 56.7% when applied alone at a concentration of 5 × 106 OBs/mL to 97.5% with the addition of 0.5% A. indica oil. A significant reduction in damage (30.7 ± 4.2% to 13.6 ± 1.0%) and loss (24.8 ± 3.5% to 7.2 ± 0.6%), and an increase in yield (523.0 ± 28.2 to 875.1 ± 47.8 Kg/ha) of M. vitrata grain was further demonstrated by Sokame et al. [153] during the first of two growing seasons in 2011 when using MaviMNPV as compared to control treatments in the field. Although MaviMNPV outperformed the botanical oils evaluated during this growing season, treatment with the chemical deltamethrin was observed to provide the greatest degree of M. vitrata control. The combination of MaviMNPV with A. indica oil did, however, significantly reduce grain loss to 1.7 ± 0.3%, which was comparable to the chemical control, indicating an additive or synergistic interaction. No statistically significant results were obtained from field trials conducted during the second growing season.
2.11. Betabaculovirus Phoperculellae, Phthorimaea operculella Granulovirus (PhopGV)
The baculovirus Phthorimaea operculella granulovirus (PhopGV) was discovered in Australia in 1964 [154], although the first report in Africa was by Broodryk and Pretorius [155] in South Africa. The virus is known to infect at least six species within the family Gelechiidae, some of which are important pests of solanaceous crops worldwide. These are the potato tuber moth, Phthorimaea operculella (Zeller); the Guatemala potato tuber moth, Tecia solanivora (Povolny); the tomato leaf miner, Phthorimaea absoluta (Meyrick); Eurysacca quinoa (Povolny); Symmetrischema tangolias (Gyen); and Paraschema detectendum (Povolny) [156,157,158,159,160].
Substantial genetic analyses have been conducted on PhopGV, with partial sequence data available for most isolates and complete genome sequence data available for at least 12 isolates, including two from Africa [156,159,161]. These studies have resulted in each PhopGV isolate being assigned to either one of five groups based on partial egt gene sequences or one of four groups based on whole-genome sequence analysis. This excludes PhopGV-SA, which did not cluster within the four groups. At least 10 African PhopGV isolates have been reported, all of which originated from infected P. operculella larvae [155,156,156,161,162,163,164].
The evaluation of the first known African PhopGV isolate by Broodryk and Pretorius [155] showed a significantly lower survival of P. operculella larvae when treated with PhopGV as compared to an untreated control application. Subsequently, field trials of PhopGV as spray and powdered (combined with magnesium silicate) formulations for the control of P. operculella were evaluated by Salah and Aalbu [163] in Tunisia. In their first experiment, PhopGV was applied to the soil surface as a spray at 50, 75, 88, and 94 days after the emergence of the plants, with tubers evaluated weekly from four weeks prior to harvest for P. operculella damage. The second experiment involved the utilisation of the powdered formulation of PhopGV, with the powder applied to elevated soil surfaces 50 and 75 days after the emergence of the plants. Damage by P. operculella was evaluated at harvest by assessing 100 tubers per treatment. A significant reduction in P. operculella tuber damage was recorded for the spray formulation at only 6% damage, as compared to a control (no spray) at 19.77% damage, and a water-only spray at 29.33% damage. The powdered formulation was not significantly different from the control (no spray), at 14.33% tuber damage recorded. The effect of these treatments on stored potatoes was further examined, with both the spray and powder formulations reducing P. operculella damage after 30 days of storage.
Recently, several new African PhopGV isolates were identified in diseased larvae originating from Tunisia, Kenya, and South Africa [156,161]. Ben Tiba et al. [156] characterised five isolates: three from Tunisia, one from Kenya, and one from Yemen. A discriminating dose of 1.8 × 105 OBs/mL was utilised to evaluate the biological activity of all five isolates against neonate larvae from two geographic P. operculella populations (Italy and Tunisia) and a P. absoluta population. The highest and lowest mortalities were observed against the P. absoluta population, with PhopGV-Tu1.11 having the highest measured mean mortality at 70.3% and PhopGV-Ym14 the lowest measured mean mortality at 11.7%. The mean mortality of these isolates ranged between 34% and 60% against neonate larvae from either of the P. operculella populations. These results suggest that PhopGV could be utilised as a biopesticide for the control of both P. operculella and P. absoluta in Africa. The recent development of PhopGV into the biopesticide Tutavir (Andermatt Biocontrol, Switzerland) in Europe suggests that this virus has commercial potential, which may prompt further research and development in African countries.
3. Conclusions
The exploration and investigation of baculoviruses for development as biopesticides for the management of agricultural pests have been pursued in Africa since the 1960s, when the occurrence of a CrleGV isolate was first reported in T. leucotreta larvae in the Ivory Coast [72]. Since then, the occurrence of at least a further series of 10 baculovirus species has been reported in agriculturally important pests, with numerous distinct isolates of some of these species (Table 1). To date, at least 11 baculovirus products have been registered and made commercially available for use in Africa, several of these being based on African isolates and produced in Africa. However, to date, there are only two known commercial baculovirus biopesticide production facilities in Africa: River Bioscience in South Africa and Kenya Biologics in Kenya.
Africa is a continent of high biological diversity, with the estimated plant and insect diversity for the Afrotropics second only to the Neotropics [165]. Thus, the small number of baculoviruses studied on the continent and exploited for the biological control of important lepidopteran pests must be considered the tip of the iceberg. The relatively recent discovery of the first NPV in the grapholitini tribe of tortricids [93] is considered a breakthrough of seismic proportions in the world of agricultural baculovirology. The grapholitini tribe consists of several globally important agricultural pests, such as the codling moth, C. pomonella; the false codling moth, T. leucotreta; the oriental fruit moth, G. molesta; the litchi moth, C. peltastica; and a couple of species of macadamia nut borer, T. batrachopa and C. ombrodelta. This NPV, CrpeNPV, has an excitingly broad host range, being virulent against most [93], if not all of these species, including C. pomonella resistant to its own granulovirus, CpGV [95].
Work on baculovirology in Africa is continuing, and if it retains a large exploratory component, it is likely that in the near future, a suite of novel species and isolates will emerge. Furthermore, production costs in Africa are much lower than in industrialised countries [116]. It should therefore be possible to produce baculovirus biopesticides at a much more affordable price than is the case with those produced outside of the continent and imported into Africa, creating an opportunity both for increased local uptake and for biopesticide exports. It is critical that this endeavour be encouraged and funded across the continent, in order to prepare for the growing global move towards more environmentally sustainable means of farming, with reduced chemical inputs, driven by initiatives such as the European Green Deal and Farm to Fork Strategy [67,166].
Author Contributions
Conceptualisation, S.M.; writing—original draft preparation, S.M. and M.J.; writing—review and editing, S.M. and M.J. All authors have read and agreed to the published version of the manuscript.
Funding
The APC was funded by Citrus Research International (CRI) and the Research council of Rhodes University.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the writing of the manuscript or in the decision to publish.
References
- Harrison, R.L.; Herniou, E.A.; Jehle, J.A.; Theilmann, D.A.; Burand, J.P.; Becnel, J.J.; Krell, P.J.; van Oers, M.M.; Mowery, J.D.; Bauchan, G.R. ICTV Virus Taxonomy Profile: Baculoviridae. J. Gen. Virol. 2018, 99, 1185–1186. [Google Scholar] [CrossRef] [PubMed]
- Lauzon, H.A.; Lucarotti, C.J.; Krell, P.J.; Feng, Q.; Retnakaran, A.; Arif, B.M. Sequence and Organization of the Neodiprion Lecontei Nucleopolyhedrovirus Genome. J. Virol. 2004, 78, 7023–7035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Oers, M.M.; Herniou, E.A.; Jehle, J.A.; Krell, P.J.; Abd-Alla, A.M.M.; Ribeiro, B.M.; Theilmann, D.A.; Hu, Z.; Harrison, R.L. Developments in the Classification and Nomenclature of Arthropod-Infecting Large DNA Viruses That Contain Pif Genes. Arch. Virol. 2023, 168, 182. [Google Scholar] [CrossRef]
- Hayakawa, T.; Ko, R.; Okano, K.; Seong, S.-I.; Goto, C.; Maeda, S. Sequence Analysis of the Xestia C-Nigrum Granulovirus Genome. Virology 1999, 262, 277–297. [Google Scholar] [CrossRef] [Green Version]
- Rohrmann, G.F. Baculovirus Molecular Biology. In Introduction to the Baculoviruses, Their Taxonomy, and Evolution; National Center for Biotechnology Information: Bethesda, MD, USA, 2019; pp. 11–24. [Google Scholar]
- Miller, L.K. The Baculoviruses; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Beas-Catena, A.; Sánchez-Mirón, A.; García-Camacho, F.; Contreras-Gómez, A.; Molina-Grima, E. Baculovirus Biopesticides: An Overview. JAPS J. Anim. Plant Sci. 2014, 24, 362–373. [Google Scholar]
- Lacey, L.A.; Grzywacz, D.; Shapiro-Ilan, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect Pathogens as Biological Control Agents: Back to the Future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, S.; Jukes, M. Advances in Microbial Control in IPM: Entomopathogenic Viruses. In Integrated Management of Insect Pests: Current and Future Developments; Burleigh Dodds Science Publishing: Cambridge, UK, 2019; pp. 593–648. [Google Scholar]
- Abd-Alla, A.M.; Meki, I.K.; Demirbas-Uzel, G. Insect Viruses as Biocontrol Agents: Challenges and Opportunities. In Challenges and Opportunities. Cottage Industry of Biocontrol Agents and Their Applications: Practical Aspects to Deal Biologically with Pests and Stresses Facing Strategic Crops; Springer: Berlin/Heidelberg, Germany, 2020; pp. 277–295. [Google Scholar]
- Haase, S.; Sciocco-Cap, A.; Romanowski, V. Baculovirus Insecticides in Latin America: Historical Overview, Current Status and Future Perspectives. Viruses 2015, 7, 2230–2267. [Google Scholar] [CrossRef]
- Lacey, L.A.; Frutos, R.; Kaya, H.K.; Vail, P. Insect Pathogens as Biological Control Agents: Do They Have a Future? Biol. Control 2001, 21, 230–248. [Google Scholar] [CrossRef] [Green Version]
- Szewczyk, B.; Hoyos-Carvajal, L.; Paluszek, M.; Skrzecz, I.; Souza, M.L. Baculoviruses—Re-emerging biopesticides. Biotechnol. Adv. 2006, 24, 143–160. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.L.; Peng, H.Y. Recent advances in biological control of pest insects by using viruses in China. Virol. Sin. 2007, 22, 158–162. [Google Scholar] [CrossRef]
- Dennis, S.J.; Meyers, A.E.; Hitzeroth, I.I.; Rybicki, E.P. African Horse Sickness: A Review of Current Understanding and Vaccine Development. Viruses 2019, 11, 844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, P.; Bishop, D.H.; Howard, S.; Aitchison, H.; Erasmus, B. Recombinant Baculovirus-Synthesized African Horsesickness Virus (AHSV) Outer-Capsid Protein VP2 Provides Protection against Virulent AHSV Challenge. J. Gen. Virol. 1996, 77, 2053–2057. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Torrecuadrada, J.L.; Diaz-Laviada, M.; Roy, P.; Sanchez, C.; Vela, C.; Sanchez-Vizcaino, J.M.; Casal, J.I. Full Protection against African Horsesickness (AHS) in Horses Induced by Baculovirus-Derived AHS Virus Serotype 4 VP2, VP5 and VP7. J. Gen. Virol. 1996, 77, 1211–1221. [Google Scholar] [CrossRef]
- Kanai, Y.; Rijn, P.A.; Maris-Veldhuis, M.; Kaname, Y.; Athmaram, T.N.; Roy, P. Immunogenicity of recombinant VP2 proteins of all nine serotypes of African horse sickness virus. Vaccine 2014, 32, 4932–4937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maree, S.; Paweska, J.T. Preparation of Recombinant African Horse Sickness Virus VP7 Antigen via a Simple Method and Validation of a VP7-Based Indirect ELISA for the Detection of Group-Specific IgG Antibodies in Horse Sera. J. Virol. Methods 2005, 125, 55–65. [Google Scholar] [CrossRef]
- Du Plessis, M.; Cloete, M.; Aitchison, H.; Van Dijk, A.A.; Verwoerd, D.W. Protein Aggregation Complicates the Development of Baculovirus-Expressed African Horsesickness Virus Serotype 5 VP2 Subunit Vaccines. OJVR 1998, 65, 321–329. [Google Scholar]
- Scanlen, M.; Paweska, J.T.; Verschoor, J.A.; Dijk, A.A. The Protective Efficacy of a Recombinant VP2-Based African Horsesickness Subunit Vaccine Candidate Is Determined by Adjuvant. Vaccine 2002, 20, 1079–1088. [Google Scholar] [CrossRef]
- Moran, V.C. The Phytophagous Insects and Mites of Cultivated Plants in South Africa: Patterns and Pest Status. J. Appl. Ecol. 1983, 20, 439–450. [Google Scholar] [CrossRef]
- Bell, M.A. An Evaluation of the Pest Status and Research Conducted on Phytophagous Lepidoptera on Cultivated Plants in South Africa. Afr. Entomol. 1996, 4, 161–170. [Google Scholar]
- Whitlock, V.H. Symptomatology of Two Viruses Infecting Heliothis Armigera. J. Invertebr. Pathol. 1974, 23, 70–75. [Google Scholar] [CrossRef]
- Whitlock, V.H. Dosage-Mortality Studies of a Granulosis and a Nuclear Polyhedrosis Virus of a Laboratory Strain of Heliothis Armigera. J. Invertebr. Pathol. 1978, 32, 386–387. [Google Scholar] [CrossRef]
- Whitlock, V.H. Effect of Larval Maturation on Mortality Induced by Nuclear Polyhedrosis and Granulosis Virus Infections of Heliothis Armigera. J. Invertebr. Pathol. 1977, 30, 80–86. [Google Scholar] [CrossRef]
- Roome, R.E. Field Trials with a Nuclear Polyhedrosis Virus and Bacillus Thuringiensis against Larvae of Heliothis Armigera (Hb.) (Lepidoptera, Noctuidae) on Sorghum and Cotton in Botswana. Bull. Entomol. Res. 1975, 65, 507–514. [Google Scholar] [CrossRef]
- Moore, S.D.; Pittaway, T.; Bouwer, G.; Fourie, J.G. Evaluation of Helicoverpa Armigera Nucleopolyhedrovirus, HaNPV, for Control of Helicoverpa Armigera Hübner (Noctuidae: Lepidoptera) on Citrus in South Africa. Biocontrol Sci. Technol. 2004, 14, 239–250. [Google Scholar] [CrossRef]
- Moore, S.D.; Grout, T.G.; Hattingh, V.; Hofmeyr, J.H. Thresholds and Guidelines for Intervention against Citrus Pests. SA Fruit J. 2008, 7, 77–81. [Google Scholar]
- Moore, S.D.; Kirkman, W. HELICOVIR: A Virus for the Biological Control of Bollworm. SA Fruit J. 2010, 9, 63–67. [Google Scholar]
- Anonymous. Kenya: Natural Solutions for Pest Control. Int. Pest Control 2015, 15, 132. [Google Scholar]
- Ogembo, J.G. Comparative Biological and Biochemical Studies of Two Isolates of Nuncleopolyhedrovirus Infecting Helicoverpa Armigera (Hübner). Noctuidae: Lepidoptera. Master’s Thesis, University of Zimbabwe, Harare, Zimbabwe, 2002. [Google Scholar]
- Ogembo, J.G.; Kunjeku, E.C.; Sithanantham, S. A Preliminary Study on the Pathogenicity of Two Isolates of Nucleopolyhedroviruses Infecting African Bollworm, Helicoverpa Armigera (Lepidoptera: Noctuidae). Int. J. Trop. Insect Sci. 2005, 25, 218–222. [Google Scholar] [CrossRef]
- Ogembo, J.G.; Chaeychomsri, S.; Kamiya, K.; Ishikawa, H.; Katou, Y.; Ikeda, M.; Kobayashi, M. Cloning and Comparative Characterization of Nucleopolyhedroviruses Isolated from African Bollworm, Helicoverpa Armigera (Lepidoptera: Noctuidae) in Different Geographic Regions. J. Insect Biotechnol. Sericol 2007, 76, 39–49. [Google Scholar]
- Mtanbanengwe, K. Genetic Characterisation of a Range of Geographically Distinct Helicoverpa Armigera Nucleopolyhedrovirus (HearNPV) Isolates and Evaluation of Biological Activity against South African Populations of the African Bollworm, Helicoverpa Armigera (Hubner). Ph.D. Thesis, Rhodes University, Grahamstown, South Africa, 2019. [Google Scholar]
- Baillie, V.L.; Bouwer, G. High Levels of Genetic Variation within Core Helicoverpa Armigera Nucleopolyhedrovirus Genes. Virus Genes 2012, 44, 149–162. [Google Scholar] [CrossRef]
- Baillie, V.L.; Bouwer, G. The Effect of Inoculum Dose on the Genetic Diversity Detected within Helicoverpa Armigera Nucleopolyhedrovirus Populations. J. Gen. Virol. 2013, 94, 2524–2529. [Google Scholar] [CrossRef] [PubMed]
- Grant, M.; Bouwer, G. Variation in the Time-Mortality Responses of Laboratory and Field Populations of Helicoverpa Armigera Infected with Nucleopolyhedrovirus. J. Appl. Entomol. 2009, 133, 647–650. [Google Scholar] [CrossRef]
- Bouwer, G.; Nardini, L.; Duncan, F.D. Helicoverpa Armigera (Lepidoptera: Noctuidae) Larvae That Survive Sublethal Doses of Nucleopolyhedrovirus Exhibit High Metabolic Rates. J. Insect Physiol. 2009, 55, 369–374. [Google Scholar] [CrossRef]
- Lacey, L.A.; Unruh, T.R. Biological control of codling moth (Cydia pomonella, Lepidoptera: Tortricidae) and its role in integrated pest management, with emphasis on entomopathogens. Vedalia 2005, 12, 33–60. [Google Scholar]
- Cross, J.V.; Solomon, M.G.; Chandler, D.; Jarrett, P.; Richardson, P.N.; Winstanley, D.; Bathon, H.; Huber, J.; Keller, B.; Langenbruch, G.A.; et al. Biocontrol of Pests of Apples and Pears in Northern and Central Europe: 1. Microbial Agents and Nematodes. Biocontrol Sci. Technol. 1999, 9, 125–149. [Google Scholar] [CrossRef]
- Lacey, L.A.; Thomson, D.; Vincent, C.; Arthurs, S.P. Codling Moth Granulovirus: A Comprehensive Review. Biocontrol Sci. Technol. 2008, 18, 639–663. [Google Scholar] [CrossRef]
- Knox, C.; Moore, S.D.; Luke, G.A.; Hill, M.P. Baculovirus-Based Strategies for the Management of Insect Pests: A Focus on Development and Application in South Africa. Biocontrol Sci. Technol. 2015, 25, 1–20. [Google Scholar] [CrossRef]
- Asser-Kaiser, S.; Fritsch, E.; Undorf-Spahn, K.; Kienzle, J.; Eberle, K.E.; Gund, N.A.; Reineke, A.; Zebitz, C.P.W.; Heckel, D.G.; Huber, J. Rapid Emergence of Baculovirus Resistance in Codling Moth Due to Dominant, Sex-Linked Inheritance. Science 2007, 317, 1916–1918. [Google Scholar] [CrossRef] [PubMed]
- Berling, M.; Blachere-Lopez, C.; Soubabere, O.; Lery, X.; Bonhomme, A.; Sauphanor, B.; Lopez-Ferber, M. Cydia Pomonella Granulovirus Genotypes Overcome Virus Resistance in the Codling Moth and Improve Virus Efficiency by Selection against Resistant Hosts. Appl. Environ. Microbiol. 2009, 75, 925–930. [Google Scholar] [CrossRef] [Green Version]
- Eberle, K.E.; Asser-Kaiser, S.; Sayed, S.M.; Nguyen, H.T.; Jehle, J.A. Overcoming the Resistance of Codling Moth against Conventional Cydia Pomonella Granulovirus (CpGV-M) by a New Isolate CpGV-I12. J. Invertebr. Pathol. 2008, 98, 293–298. [Google Scholar] [CrossRef]
- Fan, J.; Jehle, J.A.; Rucker, A.; Nielsen, A.L. First Evidence of CpGV Resistance of Codling Moth in the USA. Insects 2022, 13, 533. [Google Scholar] [CrossRef] [PubMed]
- Gebhardt, M.M.; Eberle, K.E.; Radtke, P.; Jehle, J.A. Baculovirus Resistance in Codling Moth Is Virus Isolate-Dependent and the Consequence of a Mutation in Viral Gene Pe38. Proc. Natl. Acad. Sci. USA 2014, 111, 15711–15716. [Google Scholar] [CrossRef] [PubMed]
- Jehle, J.A.; Schulze-Bopp, S.; Undorf-Spahn, K.; Fritsch, E. Evidence for a Second Type of Resistance against Cydia Pomonella Granulovirus in Field Populations of Codling Moths. Appl. Environ. Microbiol. 2017, 83, 02330-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauer, A.J.; Schulze-Bopp, S.; Fritsch, E.; Undorf-Spahn, K.; Jehle, J.A. A Third Type of Resistance to Cydia Pomonella Granulovirus in Codling Moths Shows a Mixed Z-Linked and Autosomal Inheritance Pattern. Appl. Environ. Microbiol. 2017, 83, 01036-17. [Google Scholar] [CrossRef] [Green Version]
- Motsoeneng, B.; Jukes, M.D.; Knox, C.M.; Hill, M.P.; Moore, S.D. Genome Analysis of a Novel South African Cydia Pomonella Granulovirus (CpGV-SA) with Resistance-Breaking Potential. Viruses 2019, 11, 658. [Google Scholar] [CrossRef] [Green Version]
- Motsoeneng, B.M. Genetic and Biological Characterisation of a Novel South African Cydia Pomonella Granulovirus (CpGV-SA) Isolate. Master’s Thesis, Rhodes University, Grahamstown, South Africa, 2016. [Google Scholar]
- Asayama, T.; Osaki, N. A Granulosis of the Diamondback Moth, Plutella Xylostella. J. Invertebr. Pathol. 1970, 15, 284–286. [Google Scholar] [CrossRef]
- Parnell, M.; Grzywacz, D.; Jones, K.A.; Brown, M.; Oduor, G.; Ong’Aro, J. The Strain Variation and Virulence of Granulovirus of Diamondback Moth (Plutella Xylostella Linnaeus, Lep., Yponomeutidae) Isolated in Kenya. J. Invertebr. Pathol. 2002, 79, 192–196. [Google Scholar] [CrossRef]
- Grzywacz, D.; Parnell, M.; Kibata, G.; Oduor, G.; Poole, J.; Ogutu, W. The Granulovirus of Plutella xylostella (Diamondback Moth, DBM) and Its Potential for Control of DBM in Kenya. Improving Biocontrol of Plutella xylostella. In Proceedings of the International Symposium, Montpellier, France, 21–24 October 2004; pp. 179–184. [Google Scholar]
- Cherry, A.; Osae, M.; Djegui, D. Relative Potency, Yield and Transmission of a Kenyan Isolate of Plutella xylostella Granulovirus in a Population of Diamondback Moth from Benin, West Africa. Improving Biocontrol of Plutella xylostella. In Proceedings of the International Symposium, Montpellier, France, 21–24 October 2004; pp. 158–162. [Google Scholar]
- Abdulkadir, F.; Marsberg, T.; Knox, C.M.; Hill, M.P.; Moore, S.D. Morphological and Genetic Characterization of a South African Plutella Xylostella Granulovirus (Plxy GV) Isolate. Afr. Entomol. 2013, 21, 168–171. [Google Scholar] [CrossRef]
- Abdulkadir, F.; Knox, C.; Marsberg, T.; Hill, M.P.; Moore, S.D. Genetic and Biological Characterisation of a Novel Plutella Xylostella Granulovirus, PlxyGV-SA. BioControl 2015, 60, 507–515. [Google Scholar] [CrossRef]
- Dezianian, A.; Sajap, A.S.; Lau, W.H.; Omar, D.; Kadir, H.A.; Mohamed, R.; Yusoh, M.R.M. Morphological Characteristics of P. Xylostella Granulovirus and Effects on Its Larval Host Diamondback Moth Plutella xylostella L. (Lepidoptera, Plutellidae). Am. J. Agric. Biol. Sci. 2010, 5, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Jukes, M.D.; Motsoeneng, B.M.; Knox, C.M.; Hill, M.P.; Moore, S.D. The Comparative Analysis of Complete Genome Sequences from Two South African Betabaculoviruses: Phthorimaea Operculella Granulovirus and Plutella Xylostella Granulovirus. Arch. Virol. 2016, 161, 2917–2920. [Google Scholar] [CrossRef] [PubMed]
- Newton, P.J. False Codling Moth Cryptophlebia leucotreta (Meyrick). In Citrus Pests in the Republic of South Africa; Bedford, E.C.G., Van den Berg, M.A., De Villiers, E.A., Eds.; Institute for Tropical and Subtropical Crops: Nelspruit, South Africa, 1998; pp. 192–200. [Google Scholar]
- Grout, T.G.; Moore, S.D. Citrus. In Insects of Cultivated Plants in Southern Africa; Prinsloo, G.L., Uys, V., Eds.; Entomological Society of Southern Africa: Pretoria, South Africa, 2015; pp. 447–501. [Google Scholar]
- Moore, S.D.; Kirkman, W.; Hattingh, V. The Host Status of Lemons for the False Codling Moth, Thaumatotibia leucotreta (Meyrick) (Lepidoptera: Tortricidae) with Particular Reference to Export Protocols. Afr. Entomol. 2015, 23, 519–525. [Google Scholar] [CrossRef]
- Moore, S.D.; Hattingh, V. A Review of Current Pre-Harvest Control Options for False Codling Moth in Citrus in Southern Africa. South Afr. Fruit J. 2012, 11, 82–85. [Google Scholar]
- Moore, S.D.; Hattingh, V. A Review of Control Measures and Risk Mitigation Options for False Codling Moth (FCM), Thaumatotibia leucotreta, and Their Efficacy. Citrus Res. Int. 2016, 1–8. Available online: https://www.citrusres.com/wp-content/uploads/2021/08/FCM-risk-mitigation-and-control-review-Dec-2016.pdf (accessed on 21 June 2023).
- Malan, A.P.; Diest, J.I.; Moore, S.D.; Addison, P. Control Options for False Codling Moth, Thaumatotibia leucotreta (Lepidoptera: Tortricidae), in South Africa, with Emphasis on the Potential Use of Entomopathogenic Nematodes and Fungi. Afr. Entomol. 2018, 26, 14–29. [Google Scholar] [CrossRef]
- Moore, S.D. Biological Control of a Phytosanitary Pest (Thaumatotibia leucotreta): A Case Study. Int. J. Environ. Res. Public Health 2021, 18, 1198. [Google Scholar] [CrossRef]
- Moore, S.D. The Development and Evaluation of Cryptophlebia leucotreta Granulovirus (CrleGV) as a Biological Control Agent for the Management of False Codling Moth, Cryptophlebia leucotreta, on Citrus. Ph.D. Thesis, Rhodes University, Grahamstown, South Africa, 2002. [Google Scholar]
- Moore, S.D.; Kirkman, W.; Stephen, P. CRYPTOGRAN: A Virus for the Biological Control of False Codling Moth. SA Fruit J. 2004, 3, 35–39. [Google Scholar]
- Moore, S.D.; Hendry, D.A.; Richards, G.I. Virulence of a South African Isolate of the Cryptophlebia leucotreta Granulovirus (CrleGV-SA) to Thaumatotibia Leucotreta Neonate Larvae. BioControl 2011, 56, 341–352. [Google Scholar] [CrossRef]
- Komai, F. A Taxonomic Review of the Genus Grapholita and Allied Genera (Lepidoptera tortricidae) in the Palaerctic Region. Entomol. Scand. 1999, 55, 1–219. [Google Scholar]
- Angélini, A.; Amargier, A.; Vandamme, P.; Duthoit, J.L. Une Virose á Granules Chez le Lepidoptére Argyroploce Leucotreta; Laboratoire de Cytopathologie: Paris, France, 1965. [Google Scholar]
- Angélini, A.; Le Rumeur, C. Sur Une Maladie à Virus d’Argyroploce Leucotreta Découverte en Côte d’Ivoire; Laboratoire de Cytopathologie: Paris, France, 1962. [Google Scholar]
- Mück, O. Biologie, verhalten und wirtshcaftliche bedeutaung von parasiten schädlicher Lepidopteren auf den Kapverden. Neue Entomol. Nachr. 1985, 18, 168. [Google Scholar]
- Whitlock, V.H. Long Virus-like Rods Associated with a Granulosis Virus of the False Codling Moth, Cryptophlebia leucotreta (Meyr). Phytophylactica 1980, 12, 181–186. [Google Scholar]
- Fritsch, E.; Huber, J. The Granulosis Virus of the False Codling Moth, Cryptophlebia leucotreta (Meyr). In Proceedings of the 4th International Colloquium for Invertebrate Pathology, Veldhoven, The Netherlands, 18–22 August 1986; p. 112. [Google Scholar]
- Fritsch, E. Das Granulosevirus des Falschen Apfelwicklers, Cryptophlebia leucotreta (Meyrick). Ph.D. Thesis, Technische Universität, Darmstadt, Germany, 1989. [Google Scholar]
- Jehle, A.J.; Fritsch, E.; Backhaus, H.; Huber, J. Risk Assessment on Genetic Engineering of Baculoviruses: In Vivo Integration of Host DNA into the Genome of Granulosis Viruses. In Proceedings of the 2nd International Symposium on the Biosafety Results of Field Tests of Genetically Modified Plants and Microorganisms, Goslar, Germany, 11–14 May 1992; Casper, R., Landsmann, J., Eds.; Biologische Bundesanstalt für Land-und Forstwirtschaft: Goslar, Germany, 1992; pp. 250–256. [Google Scholar]
- Jehle, J.A.; Backhaus, H. The Granulin Gene Region of Cryptophlebia Leucotreta Granulosis Virus: Sequence Analysis and Phylogenetic Considerations. J. Gen. Virol. 1994, 75, 3667–3671. [Google Scholar] [CrossRef]
- Singh, S.; Moore, S.D.; Spillings, B.; Hendry, D.A. South African Isolate of Cryptophlebia leucotreta Granulovirus. J. Invertebr. Pathol. 2003, 83, 249–252. [Google Scholar] [CrossRef]
- Goble, T. The Genetic Characterisation and Biological Activity of the South African Cryptophlebia leucotreta Granulovirus (CrleGV-SA) in Two Biopesticides, Cryptogran and Cryptex. BSc Honours, Rhodes University, Grahamstown, South Africa, 2007. [Google Scholar]
- Opoku-Debrah, J.K.; Hill, M.P.; Knox, C.; Moore, S.D. Overcrowding of False Codling Moth, Thaumatotibia leucotreta (Meyrick) Leads to the Isolation of Five New Cryptophlebia Leucotreta Granulovirus (CrleGV-SA) Isolates. J. Invertebr. Pathol. 2013, 112, 219–228. [Google Scholar] [CrossRef] [Green Version]
- Opoku-Debrah, J.K.; Hill, M.P.; Knox, C.; Moore, S.D. Heterogeneity in Virulence Relationships between Cryptophlebia Leucotreta Granulovirus Isolates and Geographically Distinct Host Populations: Lessons from Codling Moth Resistance to CpGV-M. BioControl 2016, 61, 449–459. [Google Scholar] [CrossRef]
- Fritsch, E. Biologische bekämpfung des falschen apfelwicklers, Cryptophlebia leucotreta (Meyrick) (Lep., Tortricidae), mit granuloseviren. Mitt. Dtsch. Ges. Allg. Angew. Ent. 1988, 6, 280–283. [Google Scholar]
- Kirkman, W. Understanding and Improving the Residual Efficacy of the Cryptophlebia Leucotreta Granulovirus (Cryptogran). Master’s Thesis, Rhodes University, Grahamstown, South Africa, 2007. [Google Scholar]
- Moore, S.D.; Kirkman, W.; Richards, G.I.; Stephen, P. The Cryptophlebia Leucotreta Granulovirus—10 Years of Commercial Field Use. Viruses 2015, 7, 1284–1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grové, T.; De Beer, M.S.; Joubert, P.H. Developing a Systems Approach for Thaumatotibia Leucotreta (Lepidoptera: Tortricidae) on ‘Hass’ Avocado in South Africa. J. Econ. Entomol. 2010, 103, 1112–1128. [Google Scholar] [CrossRef] [PubMed]
- Kessler, P.; Zingg, D. New Baculovirus Products Offer Solutions for the Biological Control of Cydia pomonella and Cryptophlebia leucotreta. In Proceedings of the 23rd International Congress of Entomology, Durban, South Africa, 6–12 July 2008. [Google Scholar]
- Quilici, S.; Verbizier, B.; Trahais, B.; Manikom, R. Note sur les ravageurs du litchi à la Réunion. Fruits 1988, 43, 459–464. [Google Scholar]
- Mamet, J.R.; Williams, J.R. The Recorded Food Plants of Mauritian Insects; Sugar Industry Research Institute: Réduit, Mauritius, 1993. [Google Scholar]
- Timm, A.E.; Geertsema, H.; Warnich, L. Population Genetic Structure of Economically Important Tortricidae (Lepidoptera) in South Africa: A Comparative Analysis. Bull. Entomol. Res. 2010, 100, 421–431. [Google Scholar] [CrossRef]
- Manrakhan, A.; Abeeluck, D.; Gokool, A. Assessment of Damage by Cryptophlebia peltastica (Meyrick) (Lepidoptera: Tortricidae) in Litchi Orchards in Mauritius. Afr. Entomol. 2008, 16, 203–208. [Google Scholar] [CrossRef]
- Marsberg, T.; Jukes, M.D.; Krejmer-Rabalska, M.; Rabalski, L.; Knox, C.M.; Moore, S.D.; Hill, M.P.; Szewczyk, B. Morphological, Genetic and Biological Characterisation of a Novel Alphabaculovirus Isolated from Cryptophlebia peltastica (Lepidoptera: Tortricidae). J. Invertebr. Pathol. 2018, 157, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.D.; Richards, G.I.; Chambers, C.; Hendry, D. An Improved Larval Diet for Commercial Mass Rearing of the False Codling Moth, Thaumatotibia leucotreta (Meyrick) (Lepidoptera: Tortricidae). Afr. Entomol. 2014, 22, 216–219. [Google Scholar] [CrossRef]
- Wennmann, J.T.; Eigenbrod, M.; Marsberg, T.; Moore, S.D.; Knox, C.A.; Hill, M.P.; Jehle, J.A. Cryptophlebia peltastica Nucleopolyhedrovirus Is Highly Infectious to Codling Moth Larvae and Cells. Appl. Environ. Microbiol. 2019, 85, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Thackeray, S.; (River Bioscience, Gqeberha, South Africa). Personal Communication, 2023.
- Rose, D.J.W.; Dewhurst, C.F.; Page, W.W. The African Armyworm Handbook; Natural Resources Institute, University of Greenwich: Chatham, ON, Canada, 2000. [Google Scholar]
- Graham, J.F. Annual Report of the Senior Entomologist; Report for the Department of Agriculture of Kenya; Cabi Direct: Glasgow, UK, 1961; Volume 2. [Google Scholar]
- Brown, E.S.; Swaine, G. Virus Disease of the African Armyworm, Spodoptera exempta (Wlk.). Bull. Entomol. Res. 1965, 56, 95–116. [Google Scholar] [CrossRef]
- Odindo, M.O. Epizootiological Observations on a Nuclear Polyhedrosis of the African Armyworm Spodoptera exempta (Walk). Insect Sci. Its Appl. 1983, 4, 291–298. [Google Scholar] [CrossRef]
- Vilaplana, L.; Wilson, K.; Redman, E.M.; Corey, J.S. Pathogen Persistence in Migratory Insects: High Levels of Vertically-Transmitted Virus Infection in Field Populations of the African Armyworm. Evol. Ecol. 2010, 24, 147–160. [Google Scholar] [CrossRef] [Green Version]
- Graham, R.I.; Tummala, Y.; Rhodes, G.; Cory, J.S.; Shirras, A.; Grzywacz, D.; Wilson, K. Development of a Real-Time QPCR Assay for Quantification of Covert Baculovirus Infections in a Major African Crop Pest. Insects 2015, 6, 746–759. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.E.; Maruniak, J.E.; Knudson, D.L. Physical Map of SeMNPV Baculovirus DNA: An AcMNPV Genomic Variant. Virology 1984, 136, 235–240. [Google Scholar] [CrossRef]
- Redman, E.M.; Wilson, K.; Grzywacz, D.; Cory, J.S. High Levels of Genetic Diversity in Spodoptera Exempta NPV from Tanzania. J. Invertebr. Pathol. 2010, 105, 190–193. [Google Scholar] [CrossRef]
- Escasa, S.R.; Harrison, R.L.; Mowery, J.D.; Bauchan, G.R.; Cory, J.S. The Complete Genome Sequence of an Alphabaculovirus from Spodoptera Exempta, an Agricultural Pest of Major Economic Significance in Africa. PLoS ONE 2019, 14, 0209937. [Google Scholar] [CrossRef]
- Harrap, K.A.; Payne, C.C.; Robertson, J.S. The Properties of Three Baculoviruses from Closely Related Hosts. Virology 1977, 79, 14–31. [Google Scholar] [CrossRef] [PubMed]
- Cherry, A.J.; Parnell, M.; Grzywacz, D.; Brown, M.; Jones, K.A. The Optimization of in Vivo Nuclear Polyhedrosis Virus Production of Spodoptera Exempta (Walker) and Spodoptera Exigua (Hubner). J. Invertebr. Pathol. 1997, 70, 50–58. [Google Scholar] [CrossRef]
- Mushobozi, W.; Grzywacz, D.; Moscardi, F.; Wilson, K. The African Armyworm Spodoptera exempta Nucleopolyhedrovirus (NPV) Production and Application Manual. Unpublished Work; 2006; pp. 1–71. Available online: https://assets.publishing.service.gov.uk/media/57a08c4be5274a27b20010e5/R8408_FTR_anx4.pdf (accessed on 21 June 2023).
- McKinley, D.J.; Brown, D.A.; Payne, C.C.; Harrap, K.A. Cross Infectivity and Activation Studies with Four Baculovirusesd. Entomophaga 1981, 26, 79–90. [Google Scholar] [CrossRef]
- Odindo, M.O. Dosage-Mortality and Time-Mortality Responses of the Armyworm Spodoptera Exempta to a Nuclear Polyhedrosis Virus. J. Invertebr. Pathol. 1981, 38, 251–255. [Google Scholar] [CrossRef]
- Grzywacz, D. Entomopathogenic Viruses for Control of African Armyworm Spodoptera Exempta in Tanzania; Natural Resources Institute, University of Greenwich: Chatham Maritime, UK, 2000. [Google Scholar]
- Graham, R.I.; Grzywacz, D.; Mushobozi, W.L.; Wilson, K. Wolbachia in a Major African Crop Pest Increases Susceptibility to Viral Disease Rather than Protects. Ecol. Lett. 2012, 15, 993–1000. [Google Scholar] [CrossRef] [Green Version]
- Mushobozi, W.L.; Grzywacz, D.; Musebe, R.; Kimani, M.; Wilson, K. New Approaches to Improve the Livelihoods of Poor Farmers and Pastoralists in Tanzania through Monitoring and Control of African Armyworm, Spodoptera exempta. Asp. Appl. Biol. 2005, 75, 35–37. [Google Scholar]
- Grzywacz, D.; Mushobozi, W.; Parnell, M.; Jolliffe, F.; Wilson, K. The Evaluation of Spodoptera exempta Nucleopolyhedrovirus (SpexNPV) for the Field Control of African Armyworm (Spodoptera exempta) in Tanzania. Crop Prot. 2008, 27, 17–24. [Google Scholar] [CrossRef]
- Parnell, M.; Dewhurst, C. Entomopathogenic Baculoviruses for Control of East African Armyworm Spodoptera exempta. In Tanzania: Field Trials of Spodoptera exempta Nucleopolyhedrovirus Carried Out against Armyworm Outbreaks on Rangeland in Tanzania; NRI Report; Natural Resources Institute: Chatham, UK, 1999; p. 32. [Google Scholar]
- Grzywacz, D.; Moore, S.; Luke, B.; Subramanian, S.; Moore, D.; Rabindra, R.J. Mass Production of Entomopathogens in Less Industrialized Countries. In Mass Production of Beneficial Organisms; Elsevier: Amsterdam, The Netherlands, 2023; pp. 431–462. [Google Scholar]
- Moscardi, F. Assessment of the Application of Baculoviruses for the Control of Lepidoptera. Annu. Rev. Entomol. 1999, 44, 257–289. [Google Scholar] [CrossRef]
- Moscardi, F. Development and Use of the Nucleopolyhedrovirus of the Velvet Bean Caterpillar in Soybeans. In Biological Control: A Global Perspective; Vincet, C., Goettel, M., Lazarovits, M.S., Eds.; CAB Publishing: Wallingford, UK, 2007; pp. 344–353. [Google Scholar]
- Grzywacz, D.; Stevenson, P.C.; Mushobozi, W.L.; Belmain, S.; Wilson, K. The Use of Indigenous Ecological Resources for Pest Control in Africa. Food Secur. 2014, 6, 86. [Google Scholar] [CrossRef] [Green Version]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J.; et al. Fall Armyworm: Impacts and Implications for Africa. Outlooks Pest Manag. 2017, 28, 196–201. [Google Scholar] [CrossRef] [Green Version]
- Nagoshi, R.N.; Meagher, R.L.; Hay-Roe, M. Inferring the Annual Migration Patterns of Fall Armyworm (Lepidoptera: Noctuidae) in the United States from Mitochondrial Haplotypes. Ecol. Evol. 2012, 2, 1458–1467. [Google Scholar] [CrossRef]
- Early, R.; González-Moreno, P.; Murphy, S.T.; Day, R. Forecasting the Global Extent of Invasion of the Cereal Pest Spodoptera Frugiperda, the Fall Armyworm. NeoBiota 2018, 40, 25–50. [Google Scholar] [CrossRef] [Green Version]
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First Report of Outbreaks of the Fall Armyworm Spodoptera frugiperda (JE Smith) (Lepidoptera, Noctuidae), a New Alien Invasive Pest in West and Central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Food and Agriculture Organization. Integrated Management of the Fall Armyworm on Maize: A Guide for Farmer Field Schools in Africa; Food and Agriculture Organization: Rome, Italy, 2018. [Google Scholar]
- Casmuz, A.; Juárez, M.L.; Socías, M.G.; Murúa, M.G.; Prieto, S.; Medina, S.; Willink, E.; Gastaminza, G. Revisión de los hospederos del gusano cogollero del maíz, Spodoptera frugiperda (Lepidoptera: Noctuidae). Rev. Soc. Entomol. Argent. 2010, 69, 209–231. [Google Scholar]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.D.; Peterson, J.A.; Hunt, T.E. Host Plants of Spodoptera Frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef] [Green Version]
- Kenis, M.; Benelli, G.; Biondi, A.; Calatayud, P.; Day, R.; Desneux, N.; Harrison, R.; Kriticos, D.; Rwomushana, I.; Berg, J. Invasiveness, Biology, Ecology, and Management of the Fall Armyworm, Spodoptera frugiperda. Entomol. Gen. 2022, 42, 1–55. [Google Scholar] [CrossRef]
- Eschen, R.; Beale, T.; Bonnin, J.M.; Constantine, K.L.; Duah, S.; Finch, E.A.; Makale, F.; Nunda, W.; Ogunmodede, A.; Pratt, C.F.; et al. Towards Estimating the Economic Cost of Invasive Alien Species to African Crop and Livestock Production. CABI Agric. Biosci. 2021, 2, 18. [Google Scholar] [CrossRef]
- Gardner, W.A.; Fuxa, J.R. Pathogens for the Suppression of the Fall Armyworm. Fla. Entomol. 1980, 63, 439–447. [Google Scholar] [CrossRef]
- Williams, T.; Goulson, D.; Caballero, P.; Cisneros, J.; Martínez, A.M.; Chapman, J.W.; Roman, D.X.; Cave, R.D. Evaluation of a Baculovirus Bioinsecticide for Small Scale Maize Growers in Latin America. Biol. Control 1999, 14, 67–75. [Google Scholar] [CrossRef] [Green Version]
- García-Banderas, D.; Tamayo-Mejía, F.; Pineda, S.; Rosa, J.I.F.; Lasa, R.; Chavarrieta-Yáñez, J.M.; Gervasio-Rosas, E.; Zamora-Avilés, N.; Morales, S.I.; Ramos-Ortiz, S.; et al. Biological Characterization of Two Spodoptera Frugiperda Nucleopolyhedrovirus Isolates from Mexico and Evaluation of One Isolate in a Small-Scale Field Trial. Biol. Control 2020, 149, 104316. [Google Scholar] [CrossRef]
- Raghunandan, B.L.; Patel, N.M.; Dave, H.J.; Mehta, D.M. Natural Occurrence of Nucleopolyhedrovirus Infecting Fall Armyworm, Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae) in Gujarat, India. J. Entomol. Zool. Stud. 2019, 7, 1040–1043. [Google Scholar]
- Firake, D.M.; Behere, G.T. Natural Mortality of Invasive Fall Armyworm, Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae) in Maize Agroecosystems of Northeast India. Biol. Control 2020, 148, 104303. [Google Scholar] [CrossRef]
- Firake, D.M.; Sharma, S.K.; Behere, G.T. Occurrence of Nuclear Polyhedrosis Virus of Invasive Fall Armyworm, Spodoptera frugiperda (JE Smith) Meghalaya North East India. Curr. Sci. 2020, 118, 1876–1877. [Google Scholar]
- Lei, C.; Yang, J.; Wang, J.; Hu, J.; Sun, X. Molecular and Biological Characterization of Spodoptera Frugiperda Multiple Nucleopolyhedrovirus Field Isolate and Genotypes from China. Insects 2020, 11, 777. [Google Scholar] [CrossRef] [PubMed]
- Wennmann, J.T.; Tepa-Yotto, G.T.; Jehle, J.A.; Goergen, G. Genome Sequence of a Spodoptera Frugiperda Multiple Nucleopolyhedrovirus Isolated from Fall Armyworm (Spodoptera frugiperda) in Nigeria, West Africa. Microbiol. Resour. Announc. 2021, 10, e00565-21. [Google Scholar] [CrossRef]
- Wolff, J.L.C.; Valicente, F.H.; Martins, R.; Oliveira, J.V.C.; Zanotto, P.M.A. Analysis of the Genome of Spodoptera Frugiperda Nucleopolyhedrovirus (SfMNPV-19) and of the High Genomic Heterogeneity in Group II Nucleopolyhedroviruses. J. Gen. Virol. 2008, 89, 1202–1211. [Google Scholar] [CrossRef]
- Lopez-Vaamonde, C. Spodoptera littoralis (Boisduval), African Cotton Leaf Worm (Noctuidae, Lepidoptera). Chapter 13: Species Accounts of 100 of the Most Invasive Alien Species in Europe; Hal Science: Lyon, France, 2009. [Google Scholar]
- Salama, H.S.; Dimetry, N.Z.; Salem, S.A. On the host preference and biology of the cotton leaf worm Spodoptera littoralis Bois. Z. Angew. Entomol. 1971, 67, 261–266. [Google Scholar] [CrossRef]
- Topper, C.; Moawad, G.; McKinley, D.; Hosny, M.; Jones, K.; Cooper, J.; El-Nagar, S.; El-Sheik, M. Field Trials with a Nuclear Polyhedrosis Virus against Spodoptera Littoralis on Cotton in Egypt. Int. J. Pest Manag. 1984, 30, 372–378. [Google Scholar]
- Jones, K.A.; Irving, N.S.; Grzywacz, D.; Moawad, G.M.; Hussein, A.H.; Fargahly, A. Application Rate Trials with a Nuclear Polyhedrosis Virus to Control Spodoptera littoralis (Boisd.) on Cotton in Egypt. Crop Prot. 1994, 13, 337–340. [Google Scholar] [CrossRef]
- Adel-Sattar, M.M.; El-Malla, M.A.; Ghoneim, Y.F.; Singab, M. Pyrethroids and Biocides Resistance in Field Strains of the Cotton Leaf Worm, Spodoptera littoralis (Boisd.) during 2006–2008 Cotton Seasons. Aust. J. Basic Appl. Sci. 2012, 6, 305–308. [Google Scholar]
- Alfy, H.; Ghareeb, R.Y.; Aziz, W.Z. Semi-Field Application of a New Formulation Based on Spodoptera Litura Nuclear Polyhedrosis Virus and Bacillus Thuringiensis Subsp. Mexicanensis Against Cotton Leaf Worm, Spodoptera Littoralis and Root Knot Nematode, Meloidogyne Incognita in Egypt. Curr. Appl. Sci. Technol. 2020, 20, 512–527. [Google Scholar]
- Gilligan, T.M.; Epstein, M.E. Tortricids of agricultural importance: Choristoneura occidentalis. In Interactive Keys Developed in Lucid 3; CDFA: Riverside, CA, USA, 2014. [Google Scholar]
- Erichsen, C.; Schoeman, A. Economic Losses Due to Insect Pests on Avocado Fruit in the Nelspruit/Hazyview Region of South Africa during 1991. South Afr. Avocado Grow. Assoc. Yearb. 1992, 15, 49–54. [Google Scholar]
- Newton, P.J.; Catling, H.D. Citrus Leafroller: Archips (=Cacoecia) Occidentalis (Walsingham). In Citrus Pests in the Republic of South Africa; Bedford, E.C.G., Van den Berg, M.A., De Villiers, E.A., Eds.; Institute for Tropical and Subtropical Crops: Nelspruit, South Africa, 1998. [Google Scholar]
- Razowski, J. Tortricidae (Lepidoptera) from South Africa. 6: Choristoneura Hübner and Procrica Diakonoff. Pol. Pismo Entomol. 2008, 77, 245–254. [Google Scholar]
- Fagua, G.; Condamine, F.L.; Dombroskie, J.J.; Byun, B.K.; Deprins, J.; Simonsen, T.J.; Baez, M.; Brunet, B.M.T.; Sperling, F.A.H. Genus Delimitation, Biogeography and Diversification of Choristoneura lederer (Lepidoptera: Tortricidae) Based on Molecular Evidence. Syst. Entomol. 2018, 44, 19–38. [Google Scholar] [CrossRef] [Green Version]
- Fagua, F.; (Department of Biological Sciences, CW 405 Biosciences Centre, University of Alberta, Edmonton, AL, Canada). Personal Communication, 2023.
- Smith, D.H.; Graça, J.V. Granulosis Virus from Cacoecia Occidentalis: Isolation and Morphological Description of a Granulosis Virus of the Citrus Leafroller, Cacoecia Occidentalis. J. Invertebr. Pathol. 1990, 55, 319–324. [Google Scholar] [CrossRef]
- Escasa, S.R.; Lauzon, H.A.M.; Mathur, A.C.; Krell, P.J.; Arif, B.M. Sequence analysis of the Choristoneura occidentalis granulovirus genome. J. Gen. Virol. 2006, 87, 1917–1933. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, R.; Tamò, M.; Lee, S.T.; Lin, M.Y.; Huang, C.C.; Hsu, Y.C. Towards Developing a Biological Control Program for Legume Pod Borer, Maruca Vitrata. In Proceedings of the International Conference on Grain Legumes, Kanpur, India, 14–16 February 2009; pp. 14–16. [Google Scholar]
- Sokame, B.M.; Tounou, A.K.; Datinon, B.; Dannon, E.A.; Agboton, C.; Srinivasan, R.; Pittendrigh, B.R.; Tamo, M. Combined Activity of Maruca Vitrata Multi-Nucleopolyhedrovirus, MaviMNPV, and Oil from Neem, Azadirachta indica Juss and Jatropha Curcas L., for the Control of Cowpea Pests. Crop Prot. 2015, 72, 150–157. [Google Scholar] [CrossRef]
- Reed, E.M. Granulosis Virus of Potato Moth. Aust. J. Sci. 1969, 31, 300–301. [Google Scholar]
- Broodryk, S.W.; Pretorius, L.M. Occurrence in South Africa of a Granulosis Virus Attacking Potato Tuber Moth, Phthorimaea operculella (Zeller) (Lepidoptera: Gelechiidae). J. Entomol. Soc. South. Afr. 1974, 37, 125–128. [Google Scholar]
- Ben Tiba, S.; Larem, A.; Laarif, A.; Fritsch, E.; Undorf-Spahn, K.; Abuelgasim Mohamed, S.; Ekesi, S.; Wennmann, J.T.; Kroschel, J.; Fattouch, S. The Potential of Novel African Isolates of Phthorimaea Operculella Granulovirus for the Control of Tuta Absoluta. J. Appl. Entomol. 2019, 143, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Carpio, C.; Dangles, O.; Dupas, S.; Léry, X.; López-Ferber, M.; Orbe, K.; Páez, D.; Rebaudo, F.; Santillán, A.; Yangari, B.; et al. Development of a viral biopesticide for the control of the Guatemala potato tuber moth Tecia solanivora. J. Invertebr. Pathol. 2013, 112, 184–191. [Google Scholar] [CrossRef]
- Mascarin, G.M.; Alves, S.B.; Rampelotti-Ferreira, F.T.; Urbano, M.R.; Demétrio, C.G.B.; Delalibera, I. Potential of a granulovirus isolate to control Phthorimaea operculella (Lepidoptera: Gelechiidae. BioControl 2010, 55, 657–671. [Google Scholar] [CrossRef]
- Larem, A.; Ben-Tiba, S.; Wennmann, J.T.; Gueli Alletti, G.; Jehle, J.A. Elucidating the Genetic Diversity of Phthorimaea operculella Granulovirus (PhopGV). J. Gen. Virol. 2019, 100, 679–690. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.E.C.; Metz, M.A. Classification of Tuta Absoluta (Meyrick, 1917) (Lepidoptera: Gelechiidae: Gelechiinae: Gnorimoschemini) Based on Cladistic Analysis of Morphology. Proc. Entomol. Soc. Wash. 2021, 123, 41–54. [Google Scholar]
- Jukes, M.D.; Knox, C.M.; Hill, M.P.; Moore, S.D. The Isolation and Genetic Characterisation of a South African Strain of Phthorimaea operculella Granulovirus, PhopGV-SA. Virus Res. 2014, 183, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Visser, D. The Potato Tuber Moth, Phthorimaea operculella (Zeller), in South Africa: Potential Control in Non-Refrigerated Store Environments. Ph.D. Thesis, University of Pretoria, Pretoria, South Africa, 2005. [Google Scholar]
- Salah, H.B.; Aalbu, R. Field Use of Granulosis Virus to Reduce Initial Storage Infestation of the Potato Tuber Moth, Phthorimaea operculella (Zeller), in North Africa. Agric. Ecosyst. Environ. 1992, 38, 119–126. [Google Scholar] [CrossRef]
- Smith, L.; Bellotti, A.C. Successful Biocontrol Projects with Emphasis on the Neotropics; Cornell University: Ithaca, NY, USA, 1996. [Google Scholar]
- Stork, N.E. How Many Species of Insects and Other Terrestrial Arthropods Are There on Earth? Annu. Rev. Entomol. 2018, 63, 31–45. [Google Scholar] [CrossRef] [Green Version]
- European Commission. European Commission the Green Deal: Communication from the Commission to the European Parliament, the European Council, the Council, the European Economic and Social Committee and the Committee of the Region; European Commission: Brussels, Belgium, 2019. [Google Scholar]
Figure 1.
Overview of African countries where baculoviruses have been isolated, researched, utilised in field trials, and/or commercially deployed as biopesticides.
Figure 1.
Overview of African countries where baculoviruses have been isolated, researched, utilised in field trials, and/or commercially deployed as biopesticides.


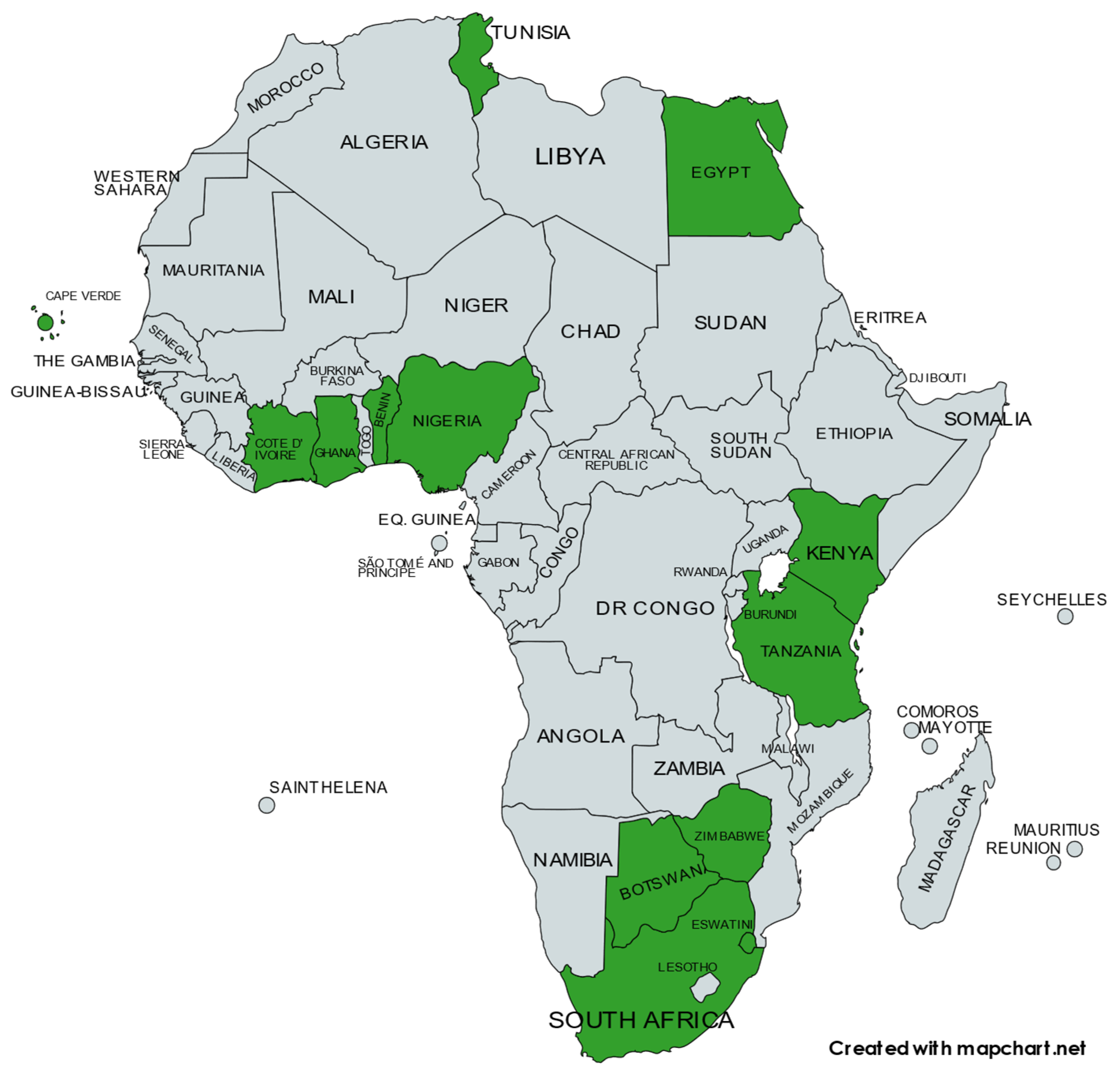{kind=link}
Table 1.
Overview of baculovirus species researched and/or utilised in Africa for the control of important agricultural pests.
Table 1.
Overview of baculovirus species researched and/or utilised in Africa for the control of important agricultural pests.
| Virus | Country of Origin | Targeted Host | Activities | Field Trial Crops | Number of Isolates 2 | Isolate References |
|---|---|---|---|---|---|---|
| ChocGV | Eswatini | C. occidentalis | LT | - | 1 | [150] |
| CpGV | South Africa | C. pomonella | LT, BA, BD 1 | - | 2 | [43,51] |
| CrleGV | Cape Verde, Ivory Coast, South Africa | T. leucotreta | LT, BA, FT, BD | Citrus, Spanish pepper, Avocado, Grape | 9 | [72,74,75,80,82] |
| CrpeNPV | South Africa | C. peltastica, C. pomonella, G. molesta, T. batrachopa, T. leucotreta | LT, BA, FT, BD | Citrus, Pome fruit, Stone fruit, Macadamia, Litchi | 1 | [93] |
| HearNPV | Botswana, Kenya, Ghana, South Africa, Zimbabwe | H. armigera | LT, BA, FT, BD | Sorghum, Cotton, Tomato, Citrus | 39 | [24,27,28,32,34,35,36] |
| MaviMNPV | Benin | M. vitrata | BA, FT | Legume | 1 | [152] |
| PhopGV | Kenya, South Africa, Tunisia | P. operculella, P. absoluta | LT, BA, FT | Potato, Tomato | 10 | [155,156,159,161,162,163,164] |
| PlxyGV | Benin, Kenya, South Africa | P. xylostella | LT, BA, FT | Kale | 15 | [54,57] |
| SfNPV | Kenya, Nigeria | S. frugiperda | LT, BD 1 | - | 2 | [127,136] |
| SpexNPV | Tanzania, Kenya | S. exempta, S. frugiperda | LT, BA, FT, BD | Pasture | 2 | [105,106] |
| SpliNPV | Egypt | S. littoralis | FT | Cotton | 2 | [140,142] |
LT = laboratory testing, BA = biological assay, FT = field trials, BD = biopesticide development. 1 Biopesticides developed outside of Africa with non-African isolate. 2 Estimated isolate number based on individual reports.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Moore, S.; Jukes, M. The History of Baculovirology in Africa. Viruses 2023, 15, 1519. https://doi.org/10.3390/v15071519
AMA Style
Moore S, Jukes M. The History of Baculovirology in Africa. Viruses. 2023; 15(7):1519. https://doi.org/10.3390/v15071519
Chicago/Turabian StyleMoore, Sean, and Michael Jukes. 2023. "The History of Baculovirology in Africa" Viruses 15, no. 7: 1519. https://doi.org/10.3390/v15071519
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.