
Identification of a Conserved, Linear Epitope on VP3 of Enterovirus A Species Recognized by a Broad-Spectrum Monoclonal Antibody

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Cells, Viruses and Anti-Serum
2.3. Generation of Monoclonal Antibody (mAb)
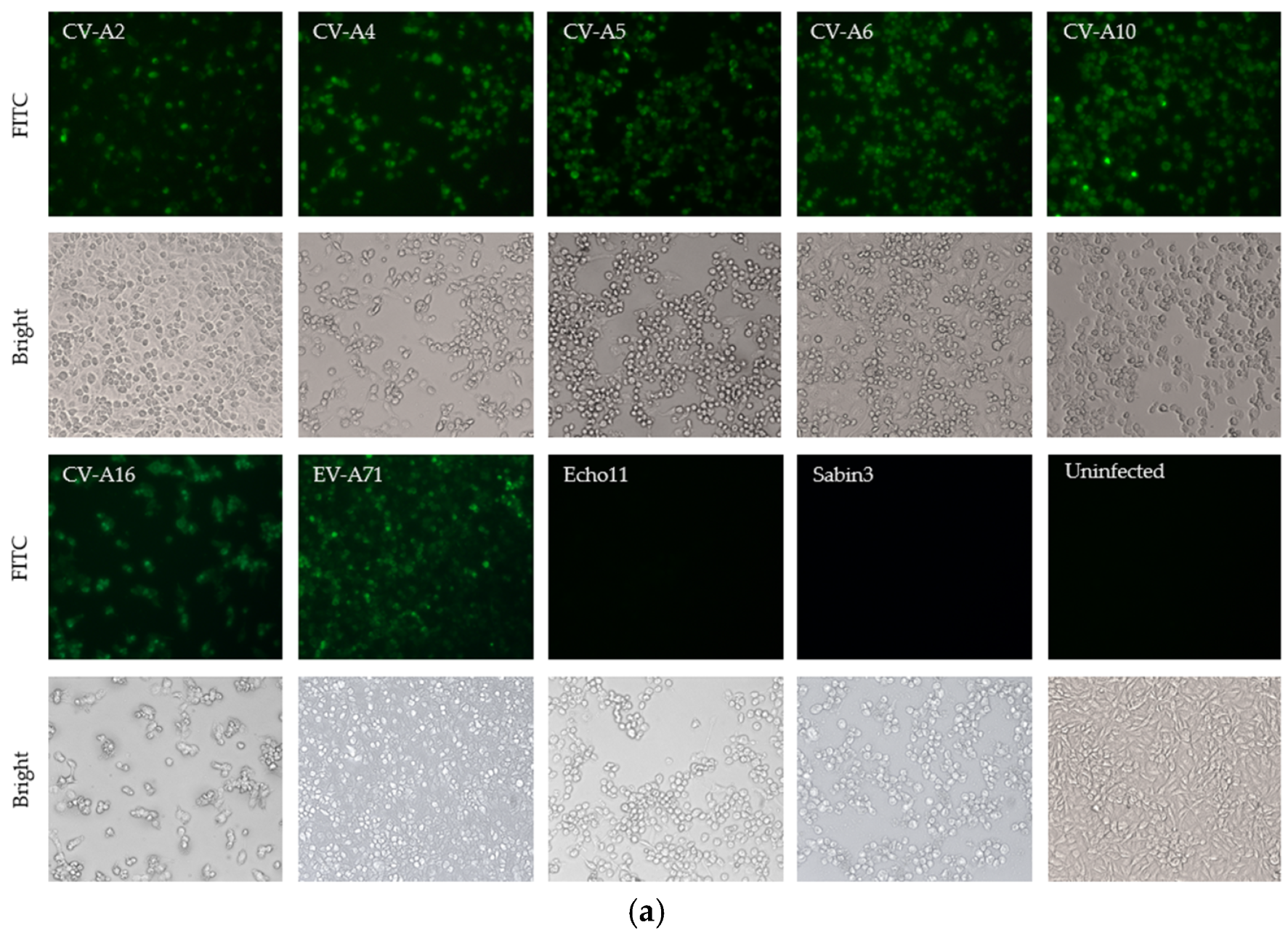
2.4. Indirect Immunofluorescence Assay (IFA)
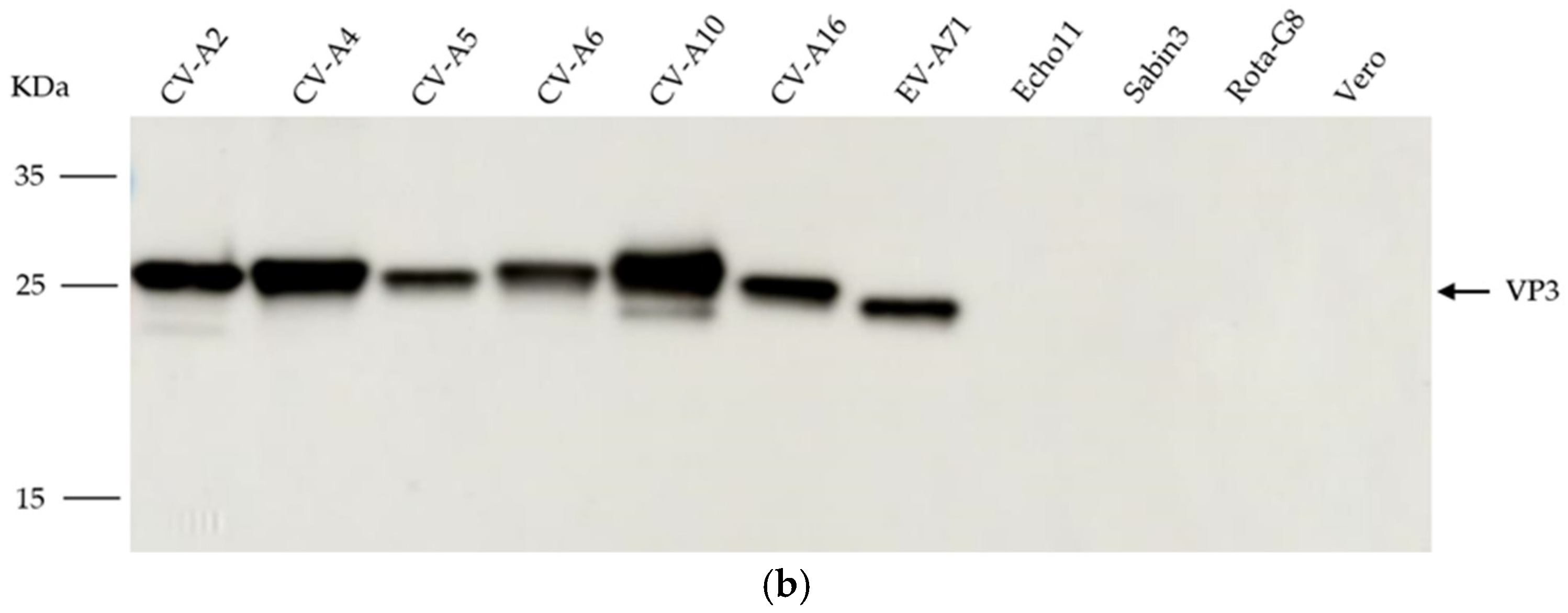
2.5. Western Blotting
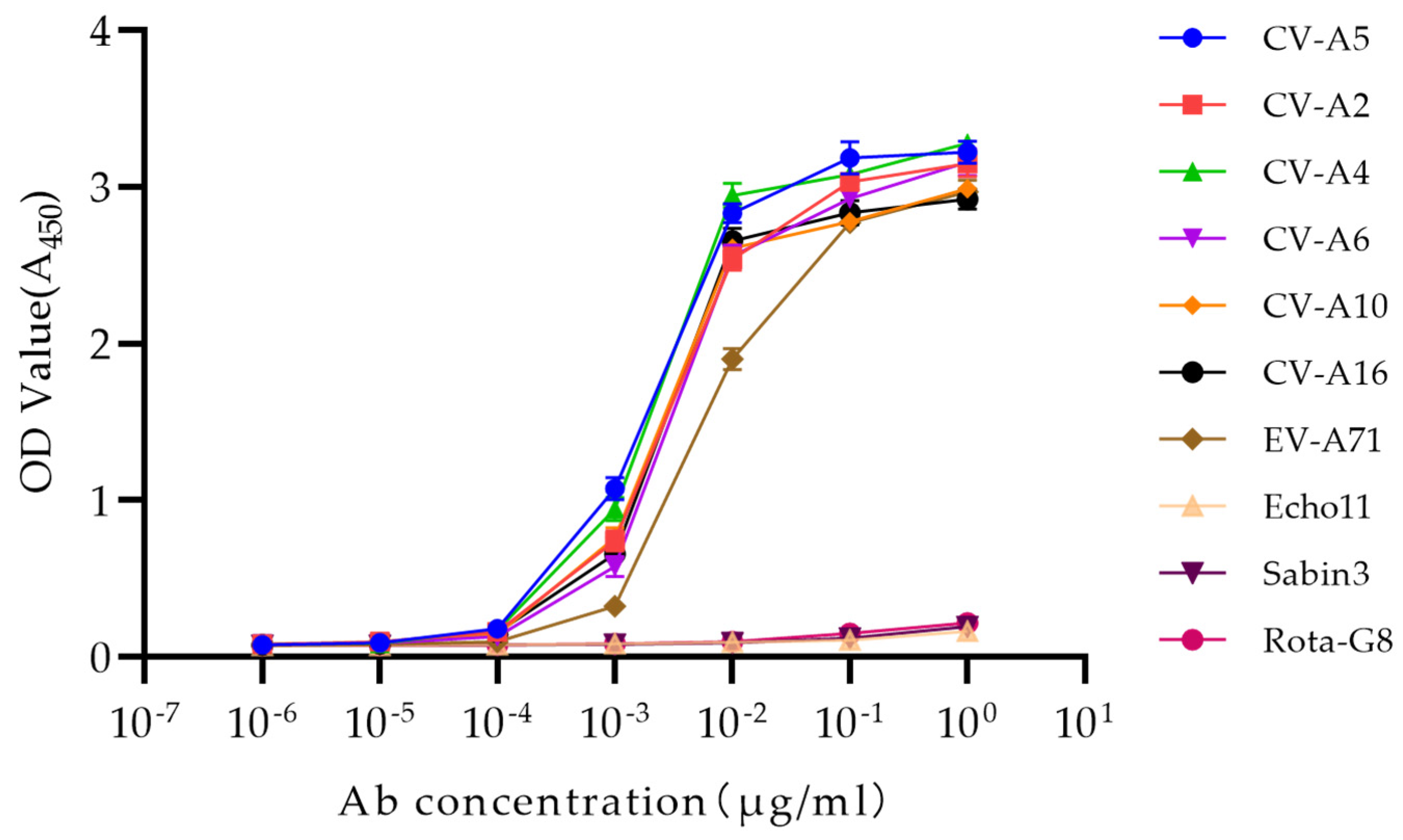
2.6. Indirect ELISA
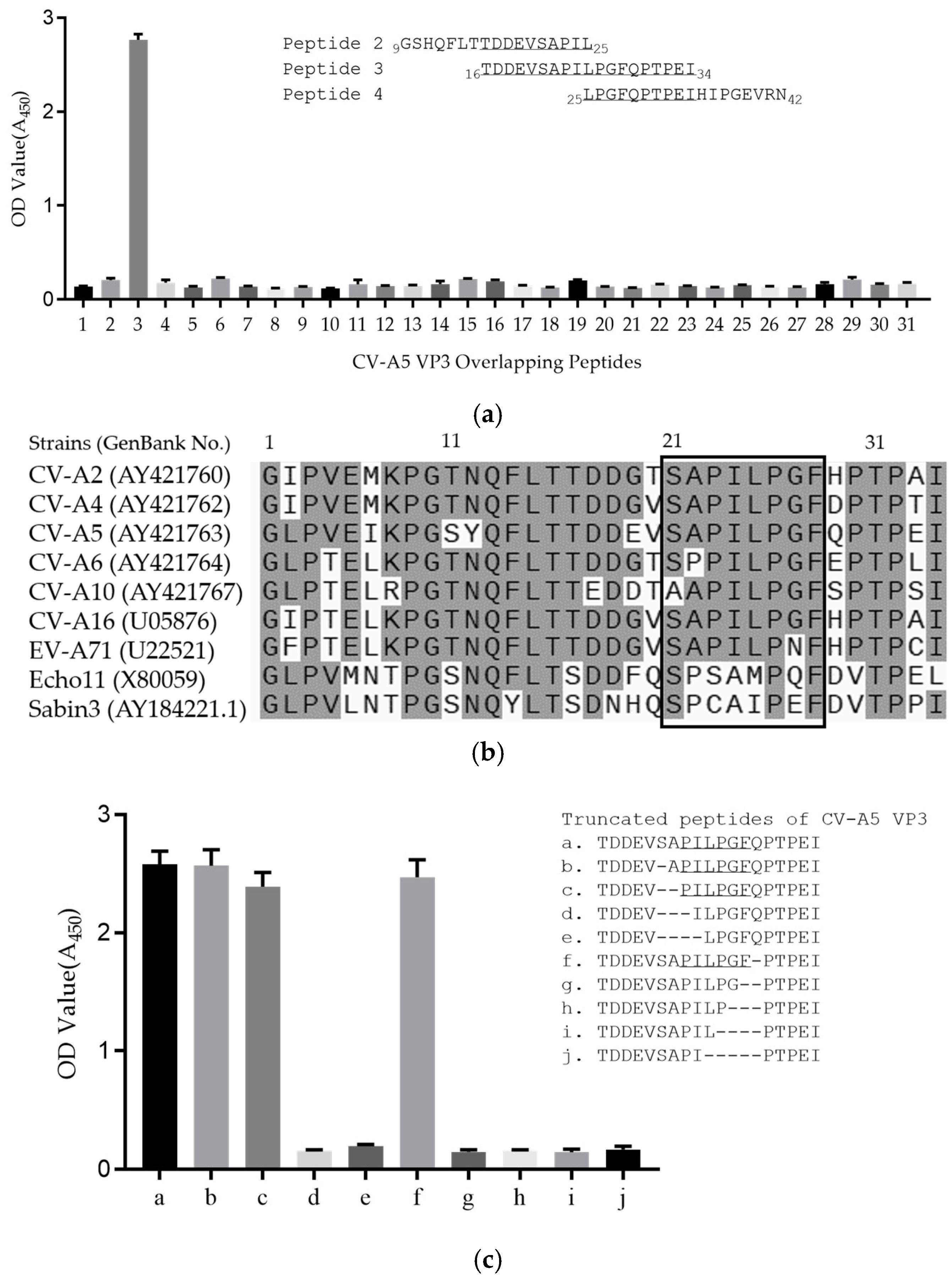
2.7. Epitope Mapping of mAb
2.8. BLAST of mAb Epitope
3. Results
3.1. Generation and Characterization of mAb 1A11
3.2. Binding Specificity of the Antibody to the Enterovirus A Species
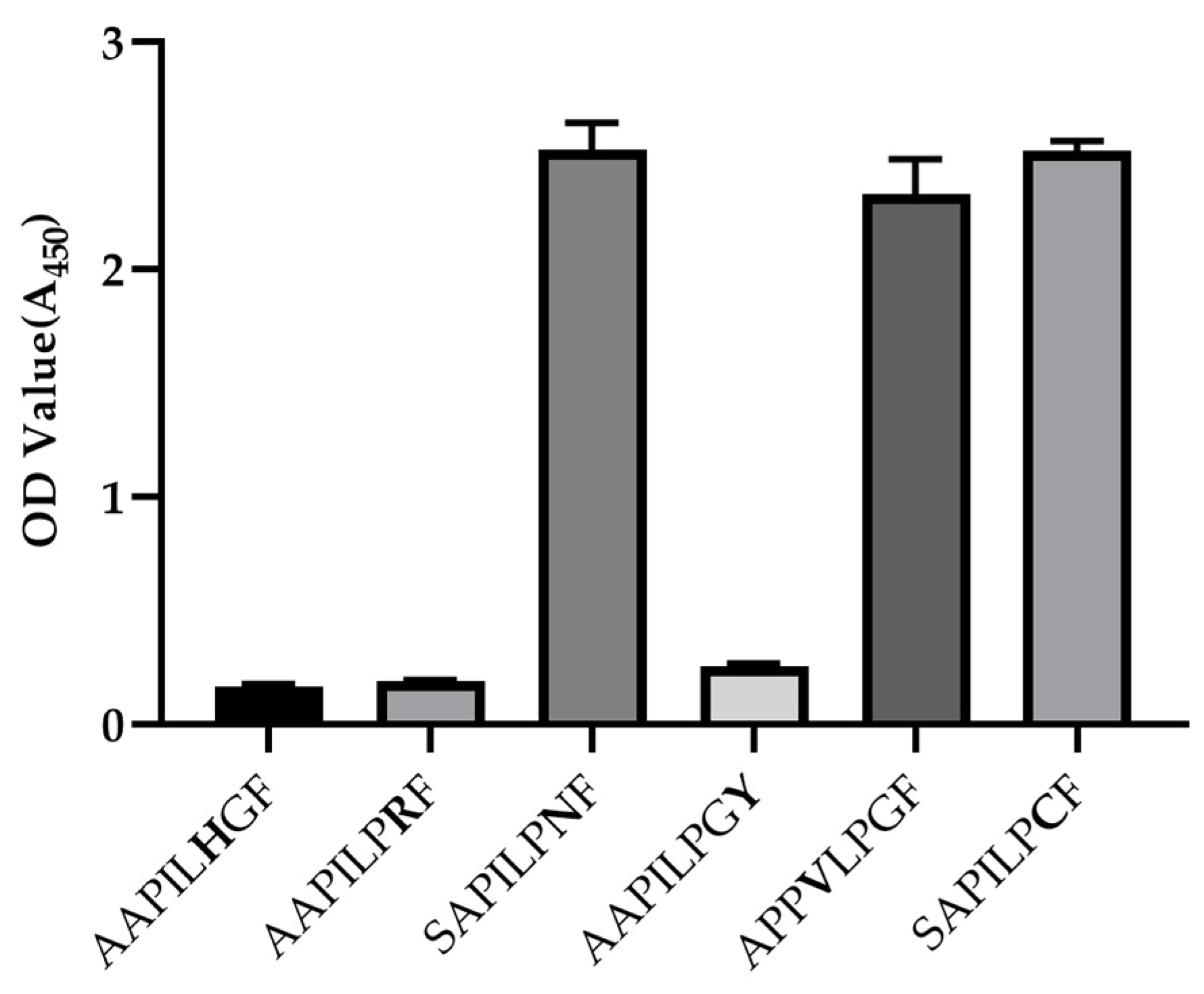
3.3. Epitope Mapping of mAb 1A11
3.4. BLAST Analysis of 1A11 Epitope
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Meng, X.D.; Tong, Y.; Wei, Z.N.; Wang, L.; Mai, J.Y.; Wu, Y.; Luo, Z.Y.; Li, S.; Li, M.; Wang, S.; et al. Epidemical and etiological study on hand, foot and mouth disease following EV-A71 vaccination in Xiangyang, China. Sci. Rep. 2020, 10, 20909. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Ren, M.; Chen, S.; Nie, T.; Cui, J.; Ran, L.; Li, Z.; Chang, Z. Pathogen spectrum of hand, foot, and mouth disease based on laboratory surveillance—China, 2018. China CDC Wkly. 2020, 2, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Wan, Z.; Li, Y.; Hu, Y.; Jin, X.; Zhang, C. National epidemiology and evolutionary history of four hand, foot and mouth disease-related enteroviruses in China from 2008 to 2016. Virol. Sin. 2020, 35, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Chen, T.; Peng, J.; Luo, Y.; Du, L.; Lu, Z.; He, J.; Liu, C.; Gan, Q.; Ma, W.; et al. The spatial-temporal distribution and etiological characteristics of hand-foot-and-mouth disease before and after EV-A71 vaccination in Kunming, China, 2017–2020. Sci. Rep. 2022, 12, 17028. [Google Scholar] [CrossRef] [PubMed]
- Nikonov, O.S.; Chernykh, E.S.; Garber, M.B.; Nikonova, E.Y. Enteroviruses: Classification, diseases they cause, and approaches to development of antiviral drugs. Biochemistry 2017, 82, 1615–1631. [Google Scholar] [CrossRef]
- Lo, C.W.; Wu, K.G.; Lin, M.C.; Chen, C.J.; Ho, D.M.; Tang, R.B.; Chan, Y.J. Application of a molecular method for the classification of human enteroviruses and its correlation with clinical manifestations. J. Microbiol. Immunol. Infect. 2010, 43, 354–359. [Google Scholar] [CrossRef]
- Adams, M.J.; Lefkowitz, E.J.; King, A.M.; Bamford, D.H.; Breitbart, M.; Davison, A.J.; Ghabrial, S.A.; Gorbalenya, A.E.; Knowles, N.J.; Krell, P.; et al. Ratification vote on taxonomic proposals to the International Committee on Taxonomy of Viruses (2015). Arch. Virol. 2015, 160, 1837–1850. [Google Scholar] [CrossRef]
- Adams, M.J.; Lefkowitz, E.J.; King, A.M.; Harrach, B.; Harrison, R.L.; Knowles, N.J.; Kropinski, A.M.; Krupovic, M.; Kuhn, J.H.; Mushegian, A.R.; et al. Ratification vote on taxonomic proposals to the International Committee on Taxonomy of Viruses (2016). Arch. Virol. 2016, 161, 2921–2949. [Google Scholar] [CrossRef]
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Adriaenssens, E.M.; Alfenas-Zerbini, P.; Dempsey, D.M.; Dutilh, B.E.; García, M.L.; Curtis Hendrickson, R.; et al. Recent changes to virus taxonomy ratified by the International Committee on Taxonomy of Viruses (2022). Arch. Virol. 2022, 167, 2429–2440. [Google Scholar] [CrossRef]
- Brouwer, L.; Moreni, G.; Wolthers, K.C.; Pajkrt, D. World-wide prevalence and genotype distribution of enteroviruses. Viruses 2021, 13, 434. [Google Scholar] [CrossRef]
- Shah, P.N.M.; Filman, D.J.; Karunatilaka, K.S.; Hesketh, E.L.; Groppelli, E.; Strauss, M.; Hogle, J.M. Cryo-EM structures reveal two distinct conformational states in a picornavirus cell entry intermediate. PLoS Pathog. 2020, 16, e1008920. [Google Scholar] [CrossRef]
- Zhu, L.; Sun, Y.; Fan, J.; Zhu, B.; Cao, L.; Gao, Q.; Zhang, Y.; Liu, H.; Rao, Z.; Wang, X. Structures of Coxsackievirus A10 unveil the molecular mechanisms of receptor binding and viral uncoating. Nat. Commun. 2018, 9, 4985. [Google Scholar] [CrossRef]
- Muir, P.; Kämmerer, U.; Korn, K.; Mulders, M.N.; Pöyry, T.; Weissbrich, B.; Kandolf, R.; Cleator, G.M.; van Loon, A.M. Molecular typing of enteroviruses: Current status and future requirements. The european union concerted action on virus meningitis and encephalitis. Clin. Microbiol. Rev. 1998, 11, 202–227. [Google Scholar] [CrossRef]
- Kitamura, N.; Semler, B.L.; Rothberg, P.G.; Larsen, G.R.; Adler, C.J.; Dorner, A.J.; Emini, E.A.; Hanecak, R.; Lee, J.J.; van der Werf, S.; et al. Primary structure, gene organization and polypeptide expression of poliovirus RNA. Nature 1981, 291, 547–553. [Google Scholar] [CrossRef]
- Minor, P.D. The molecular biology of poliovaccines. J. Gen. Virol. 1992, 73, 3065–3077. [Google Scholar] [CrossRef]
- Francisco-Velilla, R.; Embarc-Buh, A.; Abellan, S.; Martinez-Salas, E. Picornavirus translation strategies. FEBS Openbio 2022, 12, 1125–1141. [Google Scholar] [CrossRef]
- Mateu, M.G. Antibody recognition of picornaviruses and escape from neutralization: A structural view. Virus Res. 1995, 38, 1–24. [Google Scholar] [CrossRef]
- Minor, P.D.; Ferguson, M.; Evans, D.M.; Almond, J.W.; Icenogle, J.P. Antigenic structure of polioviruses of serotypes 1, 2 and 3. J. Gen. Virol. 1986, 67, 1283–1291. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, Y.; Li, H.; Liu, L. Hand-Foot-and-Mouth Disease-associated enterovirus and the development of multivalent HFMD Vaccines. Int. J. Mol. Sci. 2022, 24, 169. [Google Scholar] [CrossRef]
- Esposito, S.; Principi, N. Hand, foot and mouth disease: Current knowledge on clinical manifestations, epidemiology, aetiology and prevention. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 391–398. [Google Scholar] [CrossRef]
- Légeret, C.; Furlano, R. Oral ulcers in children- a clinical narrative overview. Ital. J. Pediatr. 2021, 47, 144. [Google Scholar] [CrossRef] [PubMed]
- Nassef, C.; Ziemer, C.; Morrell, D.S. Hand-foot-and-mouth disease: A new look at a classic viral rash. Curr. Opin. Pediatr. 2015, 27, 486–4891. [Google Scholar] [CrossRef] [PubMed]
- Yi, E.J.; Shin, Y.J.; Kim, J.H.; Kim, T.G.; Chang, S.Y. Enterovirus 71 infection and vaccines. Clin. Exp. Vaccine Res. 2017, 6, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Vuorinen, T.; Vainionpää, R.; Hyypiä, T. Five years’ experience of reverse-transcriptase polymerase chain reaction in daily diagnosis of enterovirus and rhinovirus infections. Clin. Infect. Dis. 2003, 37, 452–455. [Google Scholar] [CrossRef]
- Aw-Yong, K.L.; Tan, C.W.; Koh, M.T.; Sam, I.C.; Chan, Y.F. Diagnosis of human enterovirus A71 infection in Malaysia using a commercial IgM-capture enzyme-linked immunosorbent assay and an IgM-colloidal gold immunochromatographic assay. Trop. Biomed. 2016, 33, 238–245. [Google Scholar]
- Andréoletti, L.; Blassel-Damman, N.; Dewilde, A.; Vallée, L.; Cremer, R.; Hober, D.; Wattré, P. Comparison of use of cerebrospinal fluid, serum, and throat swab specimens in diagnosis of enteroviral acute neurological infection by a rapid RNA detection PCR assay. J. Clin. Microbiol. 1998, 36, 589–591. [Google Scholar] [CrossRef]
- Rittichier, K.R.; Bryan, P.A.; Bassett, K.E.; Taggart, E.W.; Enriquez, F.R.; Hillyard, D.R.; Byington, C.L. Diagnosis and outcomes of enterovirus infections in young infants. Pediatr. Infect. Dis. J. 2005, 24, 546–550. [Google Scholar] [CrossRef]
- Yerly, S.; Gervaix, A.; Simonet, V.; Caflisch, M.; Perrin, L.; Wunderli, W. Rapid and sensitive detection of enteroviruses in specimens from patients with aseptic meningitis. J. Clin. Microbiol. 1996, 34, 199–201. [Google Scholar] [CrossRef]
- Thanh, T.T.; Anh, N.T.; Tham, N.T.; Van, H.M.; Sabanathan, S.; Qui, P.T.; Ngan, T.T.; Van, T.T.; Nguyet, L.A.; Ny, N.T.; et al. Validation and utilization of an internally controlled multiplex Real-time RT-PCR assay for simultaneous detection of enteroviruses and enterovirus A71 associated with hand foot and mouth disease. Virol. J. 2015, 12, 85. [Google Scholar] [CrossRef]
- Tapparel, C.; Cordey, S.; Van Belle, S.; Turin, L.; Lee, W.M.; Regamey, N.; Meylan, P.; Mühlemann, K.; Gobbini, F.; Kaiser, L. New molecular detection tools adapted to emerging rhinoviruses and enteroviruses. J. Clin. Microbiol. 2009, 47, 1742–1749. [Google Scholar] [CrossRef]
- Samuelson, A.; Forsgren, M.; Sällberg, M. Characterization of the recognition site and diagnostic potential of an enterovirus group-reactive monoclonal antibody. Clin. Diagn. Lab. Immunol. 1995, 2, 385–386. [Google Scholar] [CrossRef]
- Yousef, G.E.; Mann, G.F.; Brown, I.N.; Mowbray, J.F. Clinical and research application of an enterovirus group-reactive monoclonal antibody. Intervirology 1987, 28, 199–205. [Google Scholar] [CrossRef]
- Man-Li, T.; Szyporta, M.; Fang, L.X.; Kwang, J. Identification and characterization of a monoclonal antibody recognizing the linear epitope RVADVI on VP1 protein of enterovirus 71. J. Med. Virol. 2012, 84, 1620–1627. [Google Scholar] [CrossRef]
- Shi, J.; Huang, X.; Liu, Q.; Huang, Z. Identification of conserved neutralizing linear epitopes within the VP1 protein of coxsackievirus A16. Vaccine 2013, 31, 2130–2136. [Google Scholar] [CrossRef]
- Ku, Z.; Ye, X.; Shi, J.; Wang, X.; Liu, Q.; Huang, Z. Single neutralizing monoclonal antibodies targeting the VP1 GH loop of enterovirus 71 inhibit both virus attachment and internalization during viral entry. J. Virol. 2015, 89, 12084–12095. [Google Scholar] [CrossRef]
- Lim, X.F.; Jia, Q.; Khong, W.X.; Yan, B.; Premanand, B.; Alonso, S.; Chow, V.T.; Kwang, J. Characterization of an isotype-dependent monoclonal antibody against linear neutralizing epitope effective for prophylaxis of enterovirus 71 infection. PLoS ONE 2012, 7, e29751. [Google Scholar] [CrossRef]
- Kiener, T.K.; Jia, Q.; Lim, X.F.; He, F.; Meng, T.; Chow, V.T.; Kwang, J. Characterization and specificity of the linear epitope of the enterovirus 71 VP2 protein. Virol. J. 2012, 9, 55. [Google Scholar] [CrossRef]
- Standardization Administration of China. Laboratory Animal-Guideline for Ethical Review of Animal Welfare. Standardization Administration of China. 2018. Available online: https://openstd.samr.gov.cn/bzgk/gb/newGbInfo?hcno=9BA619057D5C13103622A10FF4BA5D14 (accessed on 13 September 2022).
- Jin, W.P.; Lu, J.; Zhang, X.Y.; Wu, J.; Wei, Z.N.; Mai, J.Y.; Qian, S.S.; Yu, Y.T.; Meng, S.L.; Wang, Z.J.; et al. Efficacy of coxsackievirus A5 vaccine candidates in an actively immunized mouse model. J. Virol. 2021, 95, e01743-20. [Google Scholar] [CrossRef]
- Tian, Y.X.; Jin, W.P.; Wei, Z.N.; Lv, S.Y.; Wang, M.J.; Meng, S.L.; Guo, J.; Wang, Z.J.; Shen, S. Identification of specific and shared epitopes at the extreme N-terminal VP1 of Coxsackievirus A4, A2 and A5 by monoclonal antibodies. Virus Res. 2023, 328, 199074. [Google Scholar] [CrossRef]
- Yousef, G.E.; Brownm, I.N.; Mowbray, J.F. Derivation and biochemical characterization of an enterovirus group-specific monoclonal antibody. Intervirology 1987, 28, 163–170. [Google Scholar] [CrossRef]
- Saarinen, N.V.V.; Laiho, J.E.; Richardson, S.J.; Zeissler, M.; Stone, V.M.; Marjomäki, V.; Kantoluoto, T.; Horwitz, M.S.; Sioofy-Khojine, A.; Honkimaa, A.; et al. A novel rat CVB1-VP1 monoclonal antibody 3A6 detects a broad range of enteroviruses. Sci. Rep. 2018, 8, 33. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.Y.; Pierce, C.; Gray-Johnson, J.; DeLotell, J.; Shaw, C.; Chapman, N.; Yeh, E.; Schnurr, D.; Huang, Y.T. Monoclonal antibodies to VP1 recognize a broad range of enteroviruses. J. Clin. Microbiol. 2009, 47, 3108–3113. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Jin, W.P.; Wang, W.H.; Wu, J.; Lu, J.; Meng, S.L.; Wang, Z.J.; Shen, S. Generation of anti-CV-A5 polyclonal and monoclonal antibodies and establishment of quantitative antigen ELISA. Chin. J. Immunol. 2021, 37, 1346–1351. (In Chinese) [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enterovirus Species | Serotypes | Total Sequence No. | Identical Sequence No. | Sequence with a Single Mutation (No.) 1 | Percentage of Identical Sequence (%) |
|---|---|---|---|---|---|
| Enterovirus A (taxid: 138948) | CV-A2 | 63 | 63 | 0 | 100 |
| CV-A4 | 107 | 107 | 0 | 100 | |
| CV-A5 | 62 | 62 | 0 | 100 | |
| CV-A6 | 977 | 977 | 0 | 100 | |
| CV-A10 | 247 | 243 | PILHGF (2) PILPRF (2) | 98.4 | |
| CV-A16 | 523 | 522 | PILPNF (1) | 99.8 | |
| EV-A71 | 1491 | 3 | PILPNF (1487) | 0.2 | |
| EV-A91 | 2 | 0 | PILPGY (2) | 0 | |
| EV-A121 | 1 | 0 | PILPGY (1) | 0 | |
| EV-A125 | 1 | 0 | PVLPGF (1) | 0 | |
| SV19 | 9 | 8 | PILPGY (1) | 88.9 | |
| Other serotypes | 171 | 171 | 0 | 100 | |
| Enterovirus D (taxid: 138951) | EV-D68 | 1052 | 0 | PILPCF (3) | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, L.; Zhang, X.-Y.; Jin, W.-P.; Wang, C.; Qian, S.-S.; Wang, M.-J.; Wang, W.-H.; Meng, S.-L.; Guo, J.; Wang, Z.-J.; et al. Identification of a Conserved, Linear Epitope on VP3 of Enterovirus A Species Recognized by a Broad-Spectrum Monoclonal Antibody. Viruses 2023, 15, 1028. https://doi.org/10.3390/v15041028
Fu L, Zhang X-Y, Jin W-P, Wang C, Qian S-S, Wang M-J, Wang W-H, Meng S-L, Guo J, Wang Z-J, et al. Identification of a Conserved, Linear Epitope on VP3 of Enterovirus A Species Recognized by a Broad-Spectrum Monoclonal Antibody. Viruses. 2023; 15(4):1028. https://doi.org/10.3390/v15041028
Chicago/Turabian StyleFu, Lie, Xiao-Yu Zhang, Wei-Ping Jin, Chen Wang, Sha-Sha Qian, Meng-Jun Wang, Wen-Hui Wang, Sheng-Li Meng, Jing Guo, Ze-Jun Wang, and et al. 2023. "Identification of a Conserved, Linear Epitope on VP3 of Enterovirus A Species Recognized by a Broad-Spectrum Monoclonal Antibody" Viruses 15, no. 4: 1028. https://doi.org/10.3390/v15041028