
Identification and Molecular Characterization of a Novel Carlavirus Infecting Chrysanthemum morifolium in China


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Sample Collection
2.2. NGS and Sequence Assembly
2.3. Amplification of the Full-Length Genome Sequence of CiCV1-CN
2.4. Nucleic Acid and Protein Sequence Alignments
2.5. Phylogenetic and Recombination Analyses
2.6. Plant Material and Growth Conditions
2.7. PVX Construct and Viral Inoculation
2.8. RNA Extraction and Quantitative PCR (qPCR) Analysis
2.9. Statistical Analysis
3. Results
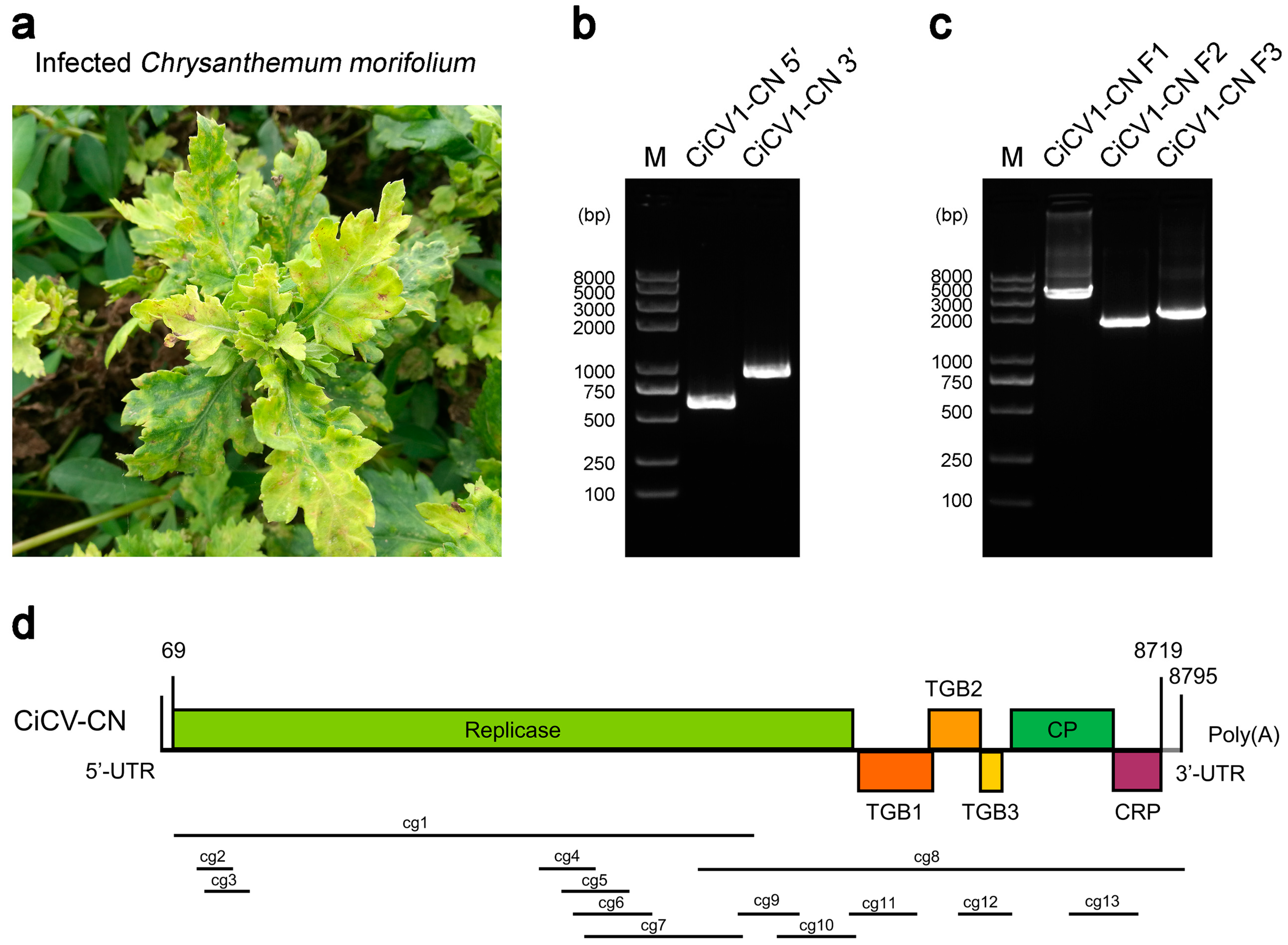
3.1. NGS and Genomic Organization of CiCV1-CN
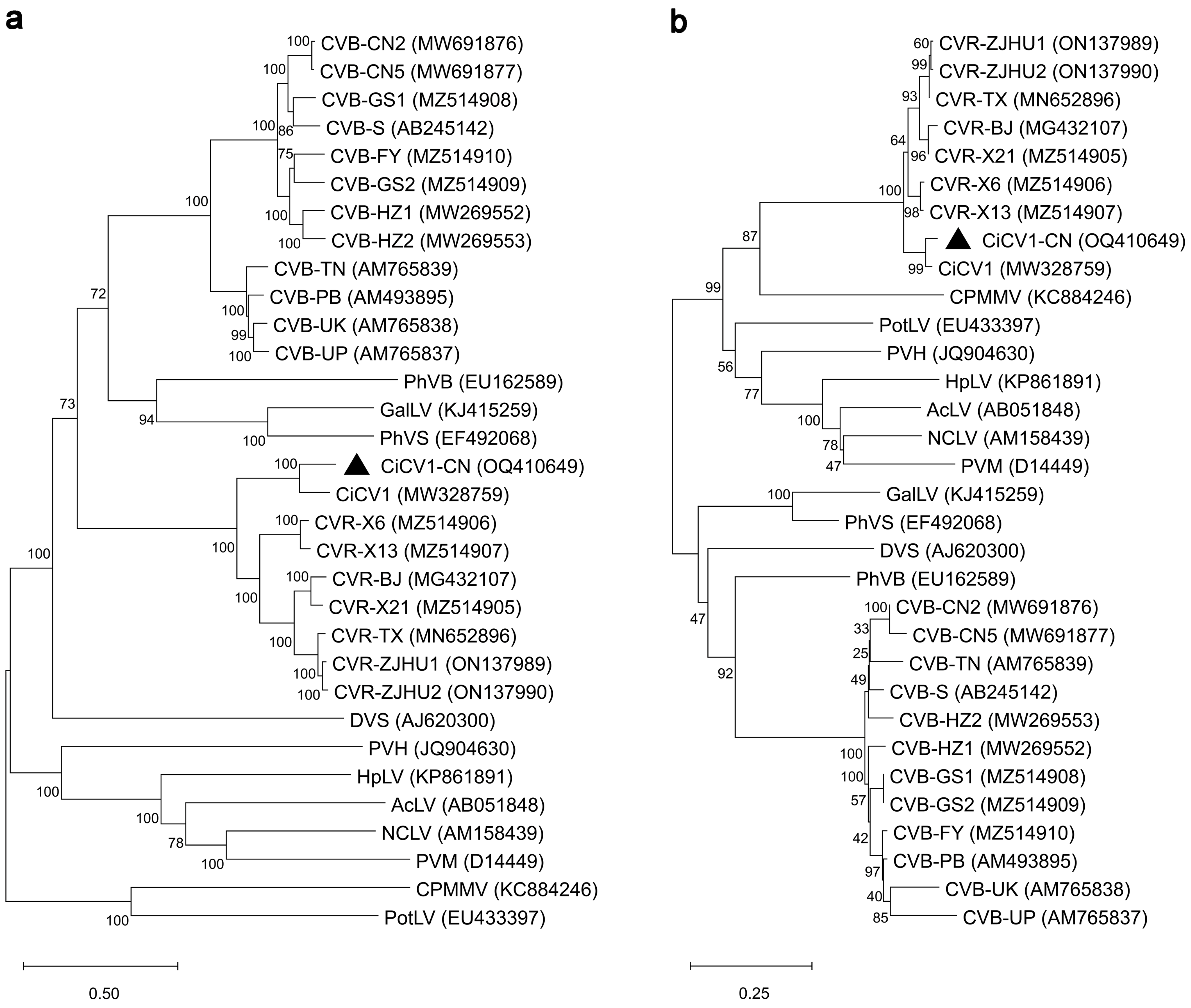
3.2. Phylogenetic and Recombination Analyses of CiCV1-CN
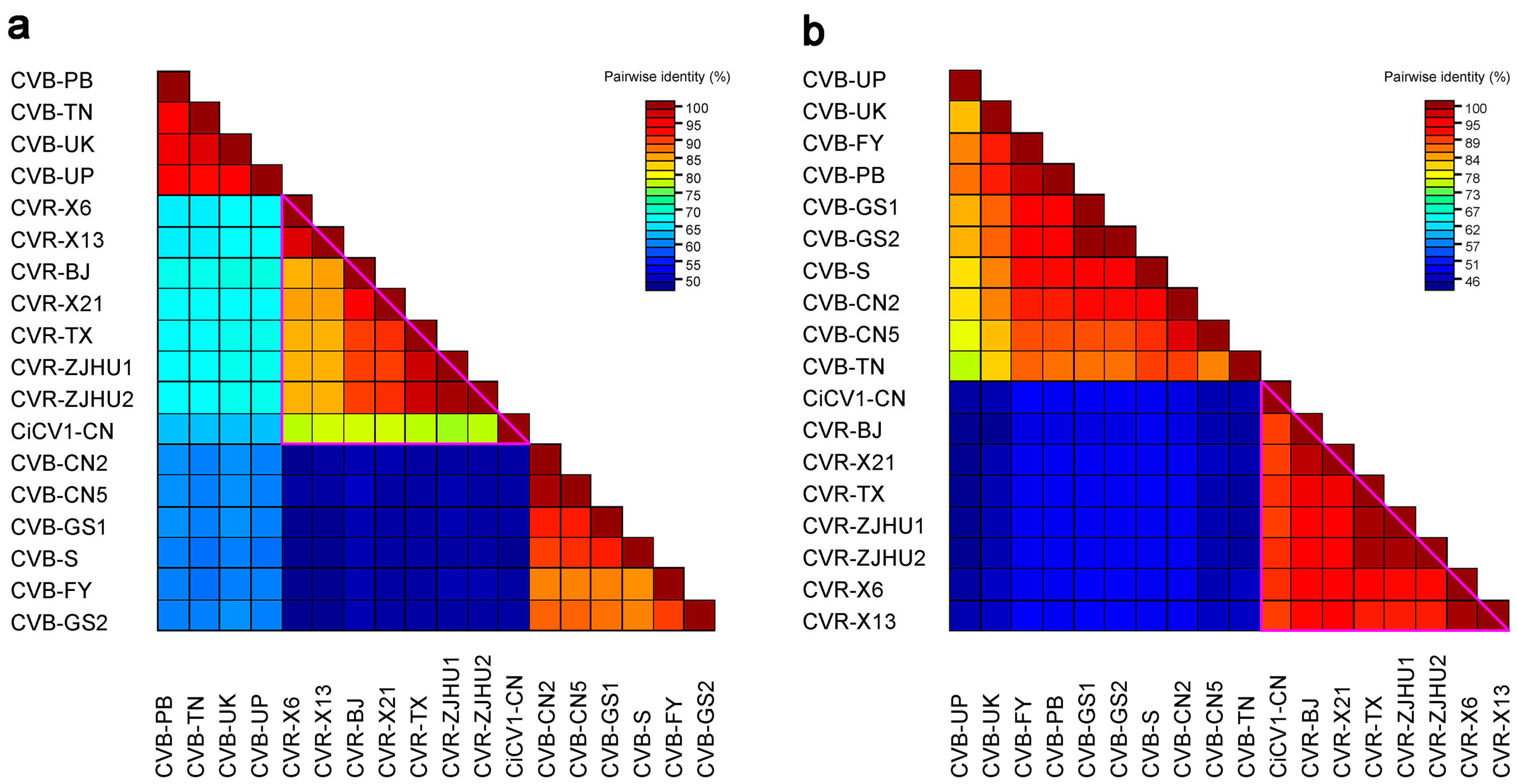
3.3. Sequence Identity Analysis of Carlaviruses from C. morifolium
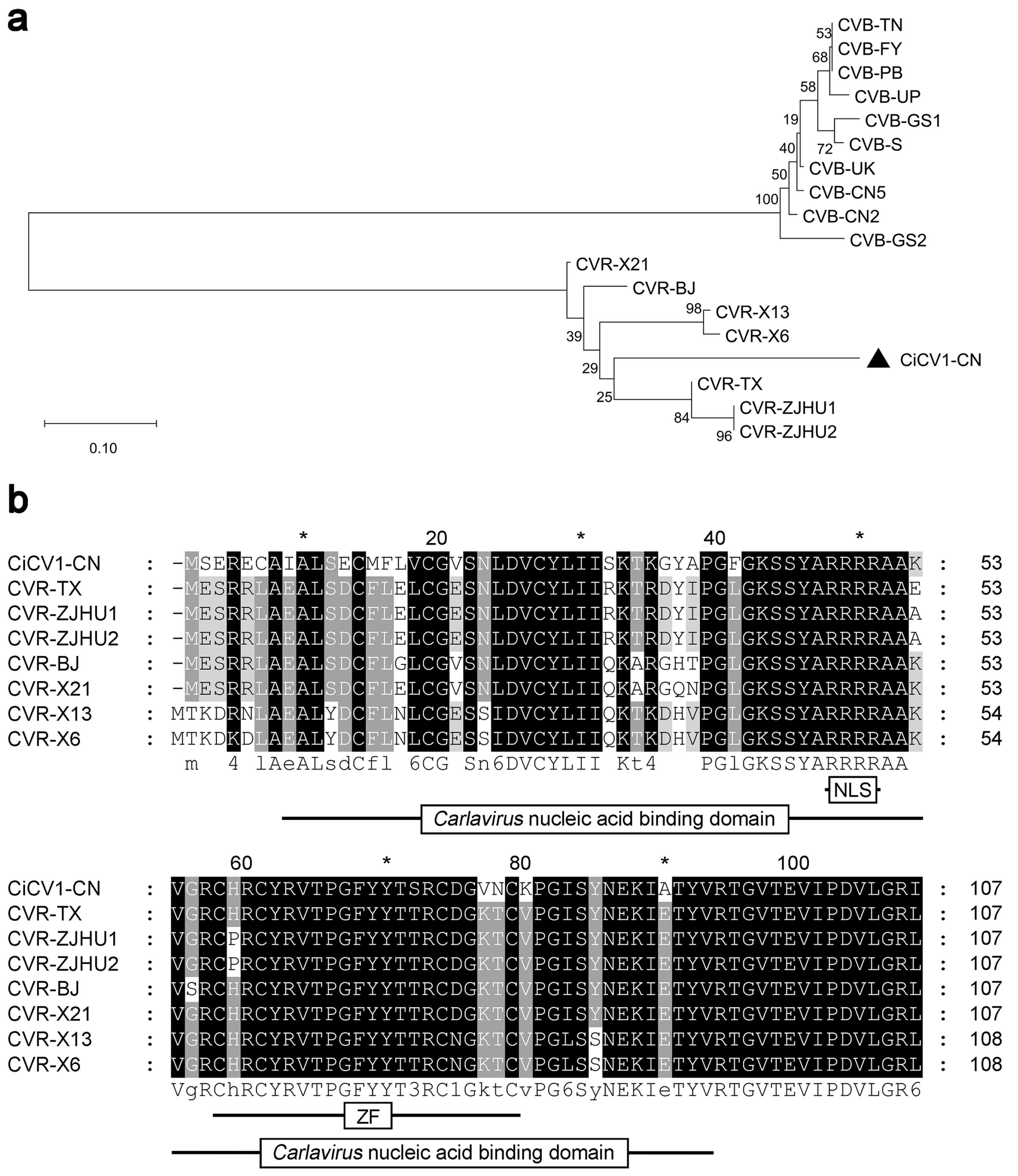
3.4. Phylogenetic and Sequence Analyses of CRP Proteins of Carlaviruses from C. morifolium
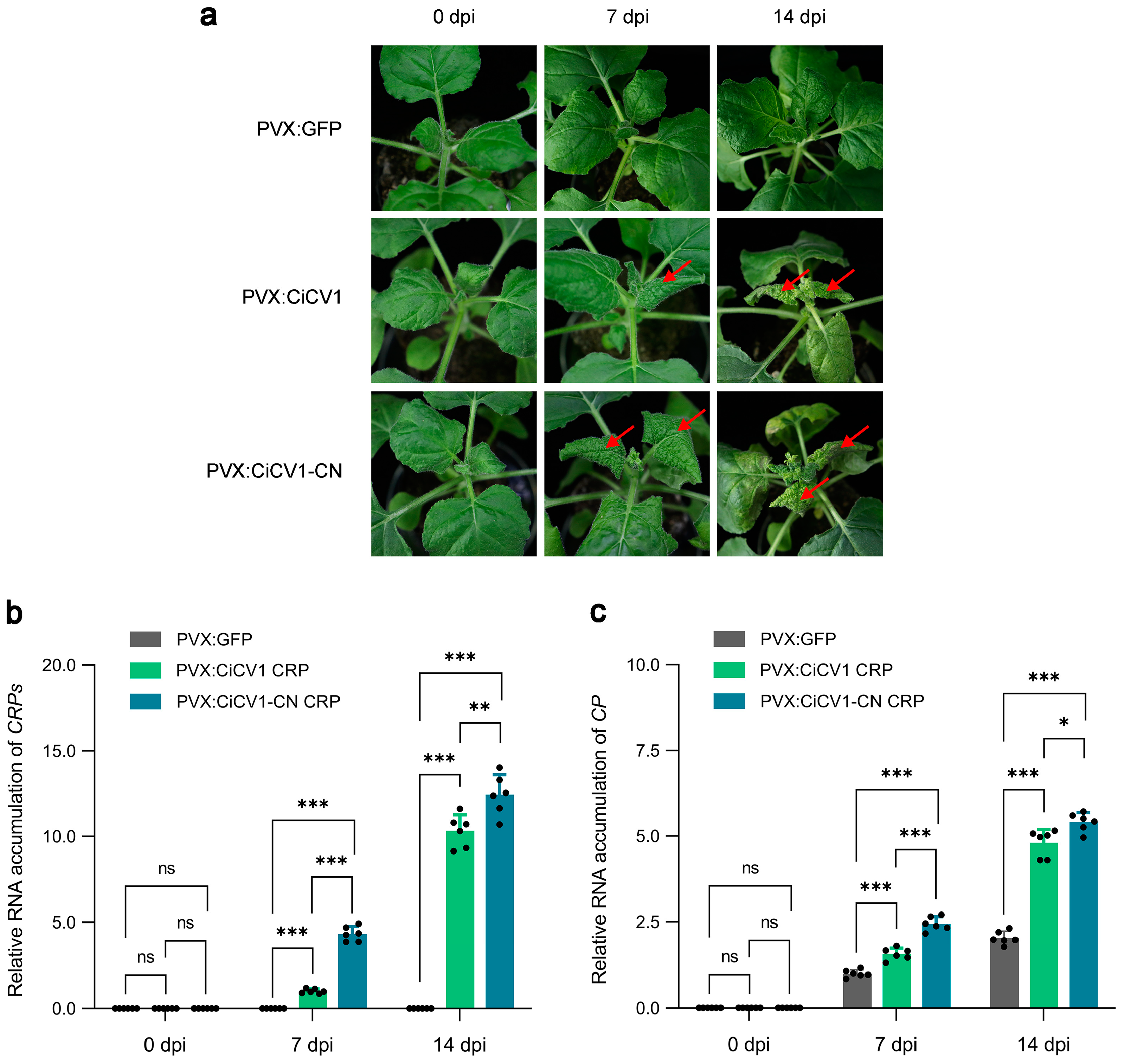
3.5. CiCV1-CN CRP Is a Pathogenicity Factor of CiCV1-CN
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Su, J.; Jiang, J.; Zhang, F.; Liu, Y.; Ding, L.; Chen, S.; Chen, F. Current achievements and future prospects in the genetic breeding of chrysanthemum: A review. Hortic. Res. 2019, 6, 109. [Google Scholar] [CrossRef]
- Zou, Q.; Wang, T.; Guo, Q.; Yang, F.; Chen, J.; Zhang, W. Combined metabolomic and transcriptomic analysis reveals redirection of the phenylpropanoid metabolic flux in different colored medicinal Chrysanthemum morifolium. Ind. Crops Prod. 2021, 164, 113343. [Google Scholar] [CrossRef]
- Zhou, L.J.; Wang, Y.; Wang, Y.; Song, A.; Jiang, J.; Chen, S.; Ding, B.; Guan, Z.; Chen, F. Transcription factor CmbHLH16 regulates petal anthocyanin homeostasis under different lights in chrysanthemum. Plant Physiol. 2022, 190, 1134–1152. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Jiang, S.; Liu, Y.; Daniyal, M.; Jian, Y.; Peng, C.; Shen, J.; Liu, S.; Wang, W. The flower head of Chrysanthemum morifolium Ramat. (Juhua): A paradigm of flowers serving as Chinese dietary herbal medicine. J. Ethnopharmacol. 2020, 261, 113043. [Google Scholar] [CrossRef] [PubMed]
- Hadizadeh, H.; Samiei, L.; Shakeri, A. Chrysanthemum, an ornamental genus with considerable medicinal value: A comprehensive review. S. Afr. J. Bot. 2022, 144, 23–43. [Google Scholar] [CrossRef]
- Zhong, X.; Yang, L.; Li, J.; Tang, Z.; Wu, C.; Zhang, L.; Zhou, X.; Wang, Y.; Wang, Z. Integrated next-generation sequencing and comparative transcriptomic analysis of leaves provides novel insights into the ethylene pathway of Chrysanthemum morifolium in response to a Chinese isolate of chrysanthemum virus B. Virol. J. 2022, 19, 182. [Google Scholar] [CrossRef] [PubMed]
- Nakano, M.; Hirakawa, H.; Fukai, E.; Toyoda, A.; Kajitani, R.; Minakuchi, Y.; Itoh, T.; Higuchi, Y.; Kozuka, T.; Bono, H.; et al. A chromosome-level genome sequence of Chrysanthemum seticuspe, a model species for hexaploid cultivated chrysanthemum. Commun. Biol. 2021, 4, 1167. [Google Scholar] [CrossRef] [PubMed]
- Eisa, E.A.; Tilly-Mándy, A.; Honfi, P.; Shala, A.Y.; Gururani, M.A. Chrysanthemum: A comprehensive review on recent developments on in vitro regeneration. Biology 2022, 11, 1774. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; He, X.; Wen, D.; Chen, S.; Chen, F.; Chen, F.; Jiang, Y. Fusarium oxysporum infection on root elicit aboveground terpene production and salicylic acid accumulation in Chrysanthemum morifolium. Plant Physiol. Biochem. 2022, 190, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Liu, X.; Ge, B.; Li, M.; Hong, B. A multiplex RT-PCR for simultaneous detection and identification of five viruses and two viroids infecting chrysanthemum. Arch. Virol. 2015, 160, 1145–1152. [Google Scholar] [CrossRef]
- Gobatto, D.; de Oliveira, L.A.; de Siqueira Franco, D.A.; Velásquez, N.; Daròs, J.A.; Eiras, M. Surveys in the chrysanthemum production areas of Brazil and Colombia reveal that weeds are potential reservoirs of chrysanthemum stunt viroid. Viruses 2019, 11, 355. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Gupta, A.; Jain, S.; Singh, D.; Verma, N. Chrysanthemum production, viral diseases and their management. In Virus Diseases of Ornamental Plants; Raj, S.K., Gaur, R.K., Yin, Z., Eds.; Springer: Singapore, 2021; pp. 261–275. [Google Scholar]
- Chirkov, S.N.; Sheveleva, A.; Snezhkina, A.; Kudryavtseva, A.; Krasnov, G.; Zakubanskiy, A.; Mitrofanova, I. Highly divergent isolates of chrysanthemum virus B and chrysanthemum virus R infecting chrysanthemum in Russia. PeerJ 2022, 10, e12607. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Xi, Y.; Shen, C.; Wang, M.; Wang, H. Occurrence of Nigrospora sphaerica causing leaf blight on Chrysanthemum morifolium in China. Crop Prot. 2022, 157, 105982. [Google Scholar] [CrossRef]
- Yan, K.R.; Zhang, Y.H.; Yang, C.B.; Ma, C.N.; He, B.W.; Mao, B.Z. First report of sweet potato feathery mottle virus infecting Chrysanthemum morifolium in China. Plant Dis. 2020, 104, 3273. [Google Scholar] [CrossRef]
- Yan, K.; Du, X.; Mao, B. Production of virus-free chrysanthemum (Chrysanthemum morifolium Ramat) by tissue culture techniques. Methods Mol. Biol. 2022, 2400, 171–186. [Google Scholar]
- Mitrofanova, I.V.; Zakubanskiy, A.V.; Mitrofanova, O.V. Viruses infecting main ornamental plants: An overview. Ornam. Hortic. 2018, 24, 95–102. [Google Scholar] [CrossRef]
- Liu, X.L.; Zhao, X.T.; Muhammad, I.; Ge, B.B.; Hong, B. Multiplex reverse transcription loop-mediated isothermal amplification for the simultaneous detection of CVB and CSVd in chrysanthemum. J. Virol. Methods 2014, 210, 26–31. [Google Scholar] [CrossRef]
- Wang, R.; Dong, J.; Wang, Z.; Zhou, T.; Li, Y.; Ding, W. Complete nucleotide sequence of a new carlavirus in chrysanthemums in China. Arch. Virol. 2018, 163, 1973–1976. [Google Scholar] [CrossRef]
- Zhao, X.T.; Liu, X.X.; Hong, B. Characterization of tomato aspermy virus isolated from chrysanthemum and elucidation of its complete nucleotide sequence. Acta Virol. 2015, 59, 204–206. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, D.; Yu, C.; Wang, Z.; Dong, J.; Shi, K.; Yuan, X. Identification of three new isolates of tomato spotted wilt virus from different hosts in China: Molecular diversity, phylogenetic and recombination analyses. Virol. J. 2016, 13, 8. [Google Scholar] [CrossRef]
- Liu, X.L.; Wei, Q.; Hong, B.; Zhao, X.T. First report of potato virus Y strain N-Wilga infecting chrysanthemum in China. Plant Dis. 2014, 98, 1589. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Z.; Pan, S.; Li, S.F. First report of chrysanthemum chlorotic mottle viroid in chrysanthemum in China. Plant Dis. 2011, 95, 1320. [Google Scholar] [CrossRef] [PubMed]
- Ohkawa, A.; Yamada, M.; Sayama, H.; Sugiyama, N.; Okuda, S.; Natsuaki, T. Complete nucleotide sequence of a Japanese isolate of chrysanthemum virus B (genus Carlavirus). Arch. Virol. 2007, 152, 2253–2258. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Li, Z.; Bernardy, M.; Wiersma, P.A.; Cheng, Z.; Xiang, Y. The complete nucleotide sequence and genome organization of pea streak virus (genus Carlavirus). Arch. Virol. 2015, 160, 2651–2654. [Google Scholar] [CrossRef]
- Thekke-Veetil, T.; McCoppin, N.K.; Hobbs, H.A.; Hartman, G.L.; Lambert, K.N.; Lim, H.S.; Domier, L.L. Discovery of a novel member of the Carlavirus genus from soybean (Glycine max L. Merr.). Pathogens 2021, 10, 223. [Google Scholar] [CrossRef]
- Diaz-Lara, A.; Mollov, D.; Golino, D.; Al Rwahnih, M. Complete genome sequence of rose virus A, the first carlavirus identified in rose. Arch. Virol. 2020, 165, 241–244. [Google Scholar] [CrossRef]
- Xing, F.; Gao, D.; Habili, N.; Wang, H.; Zhang, Z.; Cao, M.; Li, S. Identification and molecular characterization of a novel carlavirus infecting rose plants (Rosa chinensis Jacq.). Arch. Virol. 2021, 166, 3499–3502. [Google Scholar] [CrossRef]
- Villamor, D.E.V.; Sierra Mejia, A.; Martin, R.R.; Tzanetakis, I. Genomic analysis and development of infectious clone of a novel carlavirus infecting blueberry. Phytopathology 2023, 113, 98–103. [Google Scholar] [CrossRef]
- Fujita, N.; Komatsu, K.; Ayukawa, Y.; Matsuo, Y.; Hashimoto, M.; Netsu, O.; Teraoka, T.; Yamaji, Y.; Namba, S.; Arie, T. N-terminal region of cysteine-rich protein (CRP) in carlaviruses is involved in the determination of symptom types. Mol. Plant Pathol. 2018, 19, 180–190. [Google Scholar] [CrossRef]
- Singh, L.; Hallan, V.; Martin, D.P.; Ram, R.; Zaidi, A.A. Genomic sequence analysis of four new chrysanthemum virus B isolates: Evidence of RNA recombination. Arch. Virol. 2012, 157, 531–537. [Google Scholar] [CrossRef]
- Pei, Y.N.; Xian, S.; Qi, Y.H.; Zhang, L.; Wang, D.F.; Niu, Y.B. Identification and analysis of complete genomic sequence of chrysanthemum virus R isolated from Chrysanthemum morifolium. Chin. J. Biochem. Mol. Biol. 2020, 36, 217–224. [Google Scholar]
- Li, J.; Liu, M.; Wu, X.; Ye, Z.; Zhong, X.; Wang, Y.; Wang, Z. Genome-wide cloning, evolution, and recombination analyses of chrysanthemum virus R isolates from Huzhou city, Zhejiang, China. Acta Phytopathol. Sin. 2023. Available online: https://kns.cnki.net/kcms/detail/11.2184.Q.20220905.1413.001.html (accessed on 6 September 2022).
- Xing, F.; Hou, W.; Massart, S.; Gao, D.; Li, W.; Cao, M.; Zhang, Z.; Wang, H.; Li, S. RNA-Seq reveals hawthorn tree as a new natural host for apple necrotic mosaic virus, possibly associated with hawthorn mosaic disease. Plant Dis. 2020, 104, 2713–2719. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Song, Y.; Wang, Y.; Cao, M.; Hu, T.; Zhou, X. Discovery and characterization of a novel ampelovirus on firespike. Viruses 2020, 12, 1452. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Zhang, S.; Li, M.; Liu, Y.; Dong, P.; Li, S.; Kuang, M.; Li, R.; Zhou, Y. Discovery of four novel viruses associated with flower yellowing disease of green Sichuan pepper (Zanthoxylum armatum) by virome analysis. Viruses 2019, 11, 696. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, L.; Ma, L.; Tian, X.; Li, R.; Zhou, C.; Cao, M. Virome of Camellia japonica: Discovery of and molecular characterization of new viruses of different taxa in Camellias. Front. Microbiol. 2020, 11, 945. [Google Scholar] [CrossRef]
- Raco, M.; Vainio, E.J.; Sutela, S.; Eichmeier, A.; Hakalová, E.; Jung, T.; Botella, L. High diversity of novel viruses in the tree pathogen Phytophthora castaneae revealed by high-throughput sequencing of total and small RNA. Front. Microbiol. 2022, 13, 911474. [Google Scholar] [CrossRef]
- Wu, X.; Lai, Y.; Lv, L.; Han, K.; Chen, Z.; Lu, Y.; Peng, J.; Lin, L.; Chen, J.; Zheng, H.; et al. Complete genome sequence of a new achyranthes virus A isolate from Achyranthes bidentata in China. Arch. Virol. 2021, 166, 287–290. [Google Scholar] [CrossRef]
- Madeira, F.; Pearce, M.; Tivey, A.R.N.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and sequence analysis tools services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, W276–W279. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Gonzales, N.R.; Gwadz, M.; Lu, S.; Marchler, G.H.; Song, J.S.; Thanki, N.; Yamashita, R.A.; et al. The conserved domain database in 2023. Nucleic Acids Res. 2023, 51, D384–D388. [Google Scholar] [CrossRef] [PubMed]
- Paysan-Lafosse, T.; Blum, M.; Chuguransky, S.; Grego, T.; Pinto, B.L.; Salazar, G.A.; Bileschi, M.L.; Bork, P.; Bridge, A.; Colwell, L.; et al. InterPro in 2022. Nucleic Acids Res. 2023, 51, D418–D427. [Google Scholar] [CrossRef]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Wang, Z.Q.; Xiao, R.; Cao, L.; Wang, Y.; Xie, Y.; Zhou, X. Mimic phosphorylation of a βC1 protein encoded by TYLCCNB impairs its functions as a viral suppressor of RNA silencing and a symptom determinant. J. Virol. 2017, 91, e00300-17. [Google Scholar] [CrossRef]
- Hu, T.; Song, Y.; Wang, Y.; Zhou, X. Functional analysis of a novel βV1 gene identified in a geminivirus betasatellite. Sci. China Life Sci. 2020, 63, 688–696. [Google Scholar] [CrossRef]
- Qin, X.; Xue, B.; Tian, H.; Fang, C.; Yu, J.; Chen, C.; Xue, Q.; Jones, J.; Wang, X. An unconventionally secreted effector from the root knot nematode Meloidogyne incognita, Mi-ISC-1, promotes parasitism by disrupting salicylic acid biosynthesis in host plants. Mol. Plant Pathol. 2022, 23, 516–529. [Google Scholar] [CrossRef]
- Zhong, X.; Wang, Z.Q.; Xiao, R.; Wang, Y.; Xie, Y.; Zhou, X. iTRAQ analysis of the tobacco leaf proteome reveals that RNA-directed DNA methylation (RdDM) has important roles in defense against geminivirus-betasatellite infection. J. Proteomics 2017, 152, 88–101. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, H.; Wang, Z.; Guo, Y.; Hu, T.; Zhou, X. P25 and P37 proteins encoded by firespike leafroll-associated virus are viral suppressors of RNA silencing. Front. Microbiol. 2022, 13, 964156. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Q.; Zhao, Q.M.; Zhong, X.; Xiao, L.; Ma, L.X.; Wu, C.F.; Zhang, Z.; Zhang, L.Q.; Tian, Y.; Fan, W. Comparative analysis of maca (Lepidium meyenii) proteome profiles reveals insights into response mechanisms of herbal plants to high-temperature stress. BMC Plant Biol. 2020, 20, 431. [Google Scholar] [CrossRef]
- Gui, X.; Liu, C.; Qi, Y.; Zhou, X. Geminiviruses employ host DNA glycosylases to subvert DNA methylation-mediated defense. Nat. Commun. 2022, 13, 575. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Chen, B.; Li, Y.; Cao, M.; Ding, W. Complete nucleotide sequence of a new carlavirus infecting Aconitum carmichaelii in China. Arch. Virol. 2021, 166, 1513–1515. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Larrea-Sarmiento, A.; Olmedo-Velarde, A.; Borth, W.; Suzuki, J.Y.; Wall, M.M.; Melzer, M.; Hu, J. Complete genome organization and characterization of Hippeastrum latent virus. Virus Genes 2022, 58, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Lukhovitskaya, N.I.; Ignatovich, I.V.; Savenkov, E.I.; Schiemann, J.; Morozov, S.Y.; Solovyev, A.G. Role of the zinc-finger and basic motifs of chrysanthemum virus B p12 protein in nucleic acid binding, protein localization and induction of a hypersensitive response upon expression from a viral vector. J. Gen. Virol. 2009, 90, 723–733. [Google Scholar] [CrossRef] [PubMed]
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Adriaenssens, E.M.; Alfenas-Zerbini, P.; Dempsey, D.M.; Dutilh, B.E.; García, M.L.; Curtis Hendrickson, R.; et al. Recent changes to virus taxonomy ratified by the International Committee on Taxonomy of Viruses (2022). Arch. Virol. 2022, 167, 2429–2440. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y.; Zhang, R.N.; Xiang, H.Y.; Abouelnasr, H.; Li, D.W.; Yu, J.L.; McBeath, J.H.; Han, C.G. Discovery and characterization of a novel carlavirus infecting potatoes in China. PLoS ONE 2013, 8, e69255. [Google Scholar] [CrossRef] [PubMed]
- Tiberini, A.; Fontana, I.; Mercati, F.; Adams, I.; Fox, A.; Tomassoli, L. Complete genome sequence of a new isolate of caper latent virus in caper. Arch. Virol. 2021, 166, 2619–2621. [Google Scholar] [CrossRef] [PubMed]
- Mumo, N.N.; Mamati, G.E.; Ateka, E.M.; Rimberia, F.K.; Asudi, G.O.; Boykin, L.M.; Machuka, E.M.; Njuguna, J.N.; Pelle, R.; Stomeo, F. Metagenomic analysis of plant viruses associated with papaya ringspot disease in Carica papaya L. in Kenya. Front. Microbiol. 2020, 11, 205. [Google Scholar] [CrossRef]
- Lukhovitskaya, N.I.; Solovieva, A.D.; Boddeti, S.K.; Thaduri, S.; Solovyev, A.G.; Savenkov, E.I. An RNA virus-encoded zinc-finger protein acts as a plant transcription factor and induces a regulator of cell size and proliferation in two tobacco species. Plant Cell 2013, 25, 960–973. [Google Scholar] [CrossRef]
- Vetukuri, R.R.; Kalyandurg, P.B.; Saripella, G.V.; Sen, D.; Gil, J.F.; Lukhovitskaya, N.I.; Grenville-Briggs, L.J.; Savenkov, E.I. Effect of RNA silencing suppression activity of chrysanthemum virus B p12 protein on small RNA species. Arch. Virol. 2020, 165, 2953–2959. [Google Scholar] [CrossRef]
- Wu, Q.; Ding, S.W.; Zhang, Y.; Zhu, S. Identification of viruses and viroids by next-generation sequencing and homology-dependent and homology-independent algorithms. Annu. Rev. Phytopathol. 2015, 53, 425–444. [Google Scholar] [CrossRef]
- Cantalupo, P.G.; Pipas, J.M. Detecting viral sequences in NGS data. Curr. Opin. Virol. 2019, 39, 41–48. [Google Scholar] [CrossRef]
- Villamor, D.E.V.; Ho, T.; Al Rwahnih, M.; Martin, R.R.; Tzanetakis, I.E. High throughput sequencing for plant virus detection and discovery. Phytopathology 2019, 109, 716–725. [Google Scholar] [CrossRef] [PubMed]
- Rubio, L.; Galipienso, L.; Ferriol, I. Detection of plant viruses and disease management: Relevance of genetic diversity and evolution. Front. Plant Sci. 2020, 11, 1092. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.M.; Ostendorf, B.; Gautam, D.; Habili, N.; Pagay, V. Plant viral disease detection: From molecular diagnosis to optical sensing technology—A multidisciplinary review. Remote Sens. 2022, 14, 1542. [Google Scholar] [CrossRef]
- Tatineni, S.; Hein, G.L. Plant viruses of agricultural importance: Current and future perspectives of virus disease management strategies. Phytopathology 2023, 113, 117–141. [Google Scholar] [CrossRef]
- Jones, S.; Baizan-Edge, A.; MacFarlane, S.; Torrance, L. Viral diagnostics in plants using next generation sequencing: Computational analysis in practice. Front. Plant Sci. 2017, 8, 1770. [Google Scholar] [CrossRef]
- Pallás, V.; Sánchez-Navarro, J.A.; James, D. Recent advances on the multiplex molecular detection of plant viruses and viroids. Front. Microbiol. 2018, 9, 2087. [Google Scholar] [CrossRef]
- Plyusnin, I.; Kant, R.; Jääskeläinen, A.J.; Sironen, T.; Holm, L.; Vapalahti, O.; Smura, T. Novel NGS pipeline for virus discovery from a wide spectrum of hosts and sample types. Virus Evol. 2020, 6, veaa091. [Google Scholar] [CrossRef]
- Mehetre, G.T.; Leo, V.V.; Singh, G.; Sorokan, A.; Maksimov, I.; Yadav, M.K.; Upadhyaya, K.; Hashem, A.; Alsaleh, A.N.; Dawoud, T.M.; et al. Current developments and challenges in plant viral diagnostics: A systematic review. Viruses 2021, 13, 412. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Viruses | Accession No. | Genome (nt a) | 5′-UTR (nt a) | 3′-UTR (nt a) | ORF1 (aa b) | ORF2 (aa b) | ORF3 (aa b) | ORF4 (aa b) | ORF5 (aa b) | ORF6 (aa b) |
|---|---|---|---|---|---|---|---|---|---|---|
| CVR-X6 | MZ514906 | 71.3% | 79.2% | 84.8% | 76.7% | 80.3% | 71.0% | 52.4% | 90.2% | 71.3% |
| CVR-X13 | MZ514907 | 71.1% | 71.0% | 84.8% | 77.0% | 80.3% | 70.1% | 52.4% | 89.3% | 72.2% |
| CVR-BJ | MG432107 | 71.0% | 84.3% | 87.3% | 76.8% | 78.6% | 73.8% | 60.9% | 88.9% | 78.5% |
| CVR-X21 | MZ514905 | 70.9% | 71.0% | 84.8% | 77.1% | 76.9% | 74.8% | 54.0% | 89.3% | 79.4% |
| CVR-ZJHU1 | ON137989 | 70.7% | 84.5% | 87.3% | 76.0% | 76.0% | 72.9% | 54.0% | 89.6% | 77.6% |
| CVR-ZJHU2 | ON137990 | 70.4% | 84.5% | 87.3% | 76.2% | 75.5% | 73.8% | 58.7% | 89.9% | 77.6% |
| CVR-TX | MN652896 | 70.4% | 84.5% | 88.5% | 76.2% | 76.0% | 73.8% | 54.0% | 90.2% | 78.5% |
| CVB-UK | AM765838 | 60.9% | NA c | 44.6% | 61.8% | 52.4% | 49.5% | 25.0% | 39.8% | 42.5% |
| CVB-UP | AM765837 | 60.7% | NA c | 44.6% | 62.1% | 52.4% | 49.5% | 31.3% | 41.9% | 37.2% |
| CVB-PB | AM493895 | 60.6% | NA c | 44.6% | 61.7% | 51.3% | 49.5% | 32.8% | 46.1% | 42.5% |
| CVB-TN | AM765839 | 60.5% | NA c | 44.6% | 61.6% | 53.9% | 49.5% | 23.5% | 43.6% | 42.5% |
| CVB-FY | MZ514910 | 56.9% | 47.1% | 67.9% | 47.0% | 53.7% | 50.5% | 29.9% | 46.7% | 42.5% |
| CVB-GS2 | MZ514909 | 56.4% | 50.7% | 69.0% | 47.0% | 52.8% | 50.5% | 26.9% | 45.2% | 38.8% |
| CVB-GS1 | MZ514908 | 56.4% | 50.7% | 69.0% | 47.3% | 53.9% | 49.5% | 20.6% | 45.8% | 42.5% |
| CVB-CN5 | MW691877 | 56.3% | 60.0% | 67.9% | 47.1% | 54.7% | 49.5% | 22.1% | 41.9% | 38.3% |
| CVB-S | AB245142 | 56.3% | 60.0% | 70.2% | 46.7% | 52.6% | 49.5% | 26.9% | 46.4% | 40.5% |
| CVB-CN2 | MW691876 | 56.2% | 60.0% | 67.9% | 47.1% | 54.3% | 49.5% | 22.1% | 44.7% | 39.2% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Wu, X.; Liu, H.; Wang, X.; Yi, S.; Zhong, X.; Wang, Y.; Wang, Z. Identification and Molecular Characterization of a Novel Carlavirus Infecting Chrysanthemum morifolium in China. Viruses 2023, 15, 1029. https://doi.org/10.3390/v15041029
Li J, Wu X, Liu H, Wang X, Yi S, Zhong X, Wang Y, Wang Z. Identification and Molecular Characterization of a Novel Carlavirus Infecting Chrysanthemum morifolium in China. Viruses. 2023; 15(4):1029. https://doi.org/10.3390/v15041029
Chicago/Turabian StyleLi, Jiapeng, Xiaoyin Wu, Hui Liu, Xiaomei Wang, Shaokui Yi, Xueting Zhong, Yaqin Wang, and Zhanqi Wang. 2023. "Identification and Molecular Characterization of a Novel Carlavirus Infecting Chrysanthemum morifolium in China" Viruses 15, no. 4: 1029. https://doi.org/10.3390/v15041029