
The Synthetic Opioid Fentanyl Increases HIV Replication and Chemokine Co-Receptor Expression in Lymphocyte Cell Lines
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Cell Lines and Reagents
2.2. Quantification of Mu Opioid Receptor
2.3. Propagation of HIV
2.4. HIV Infection, Drug Exposure, and p24 Protein Quantification
2.5. Quantification of Integrated HIV DNA
2.6. Cell Viability
2.7. Chemokine Receptor Expression
2.8. Primary Cell Isolation and Culture
2.9. Cellular RNA Isolation and Purification
2.10. miRNA-Seq and Data Analysis
2.11. RNAseq Analysis
2.12. Statistical Analysis
3. Results
3.1. HIV-Susceptible and HIV-Infected Cell Lines Express Mu Opioid Receptor
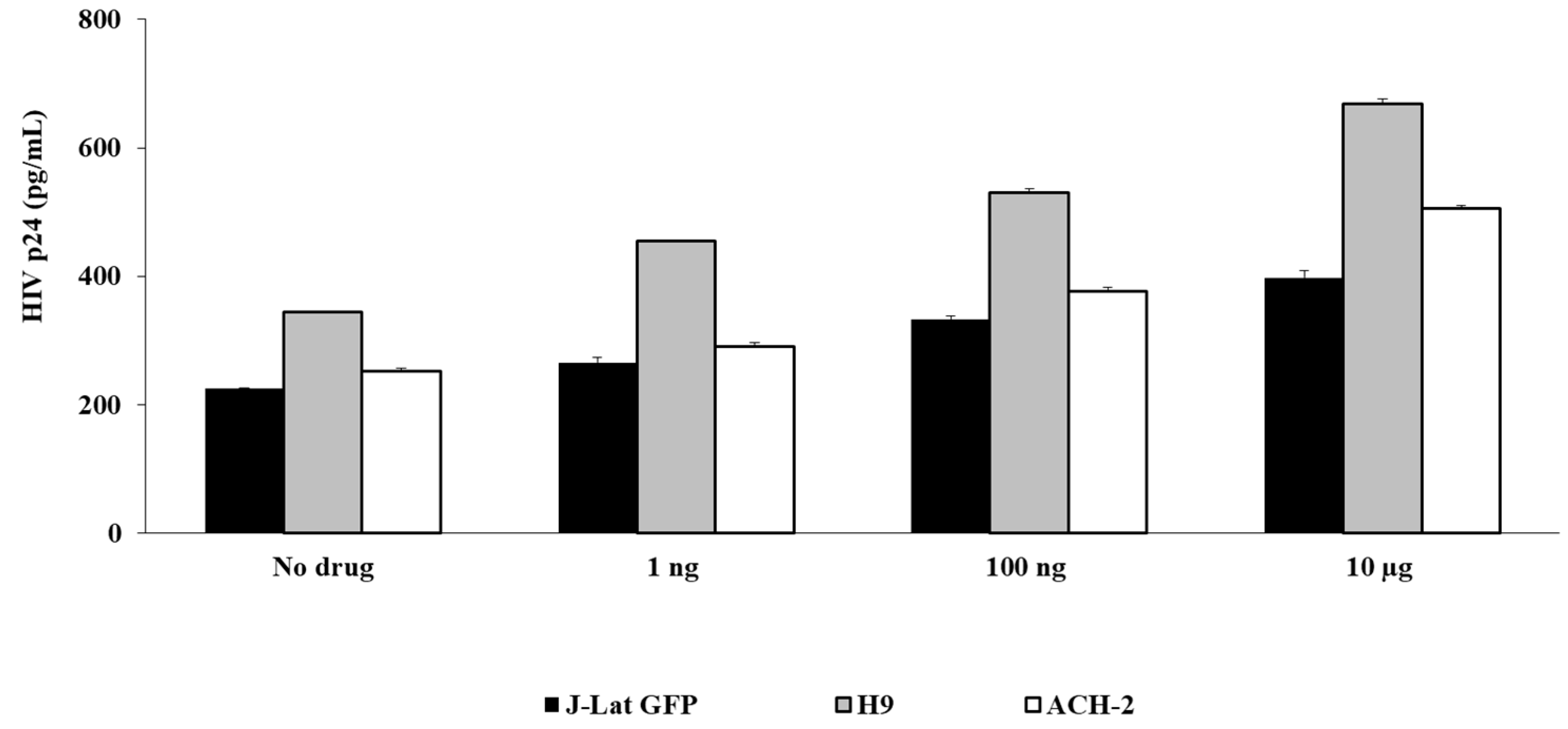
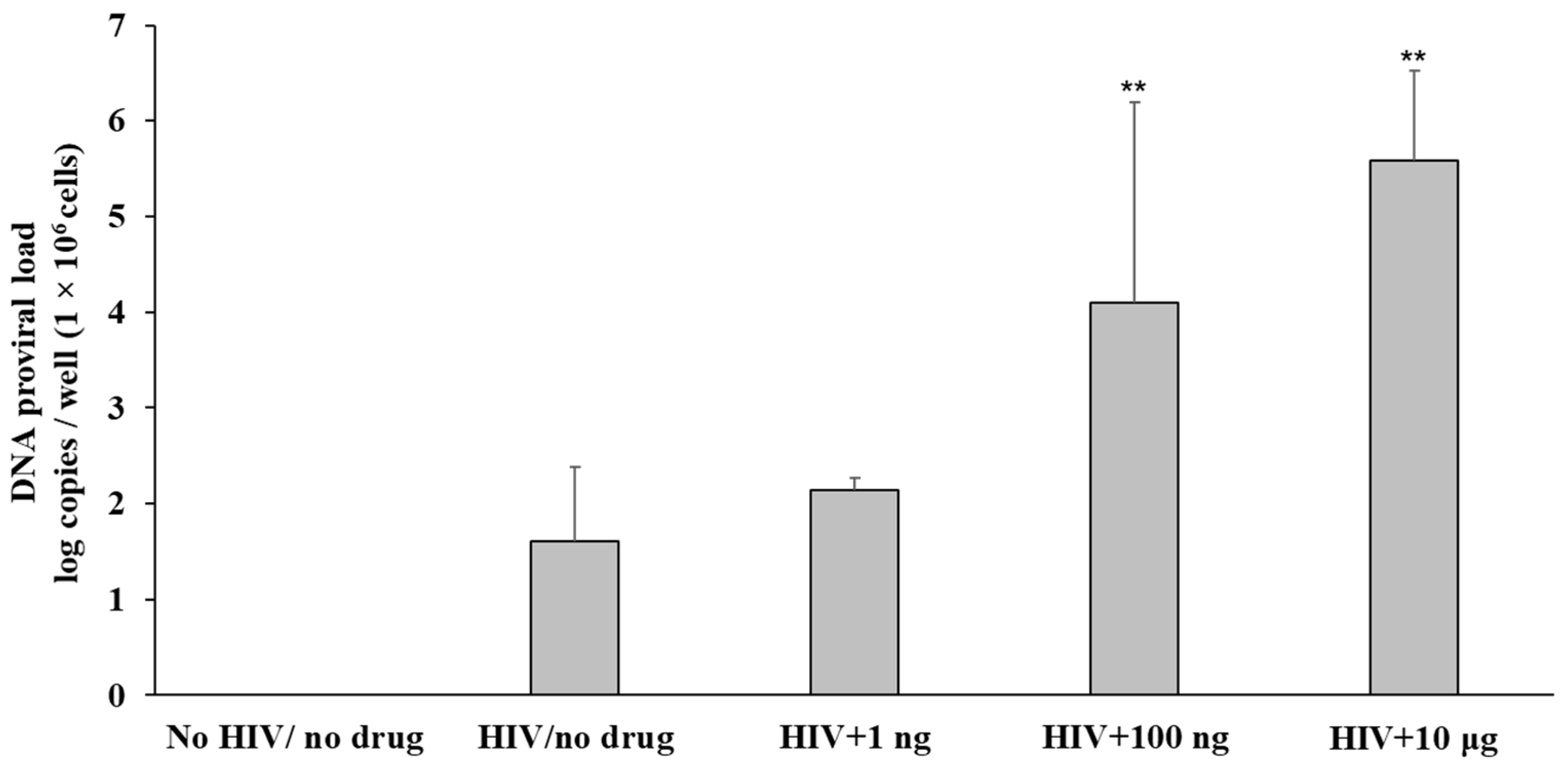
3.2. Fentanyl Enhances HIV Replication
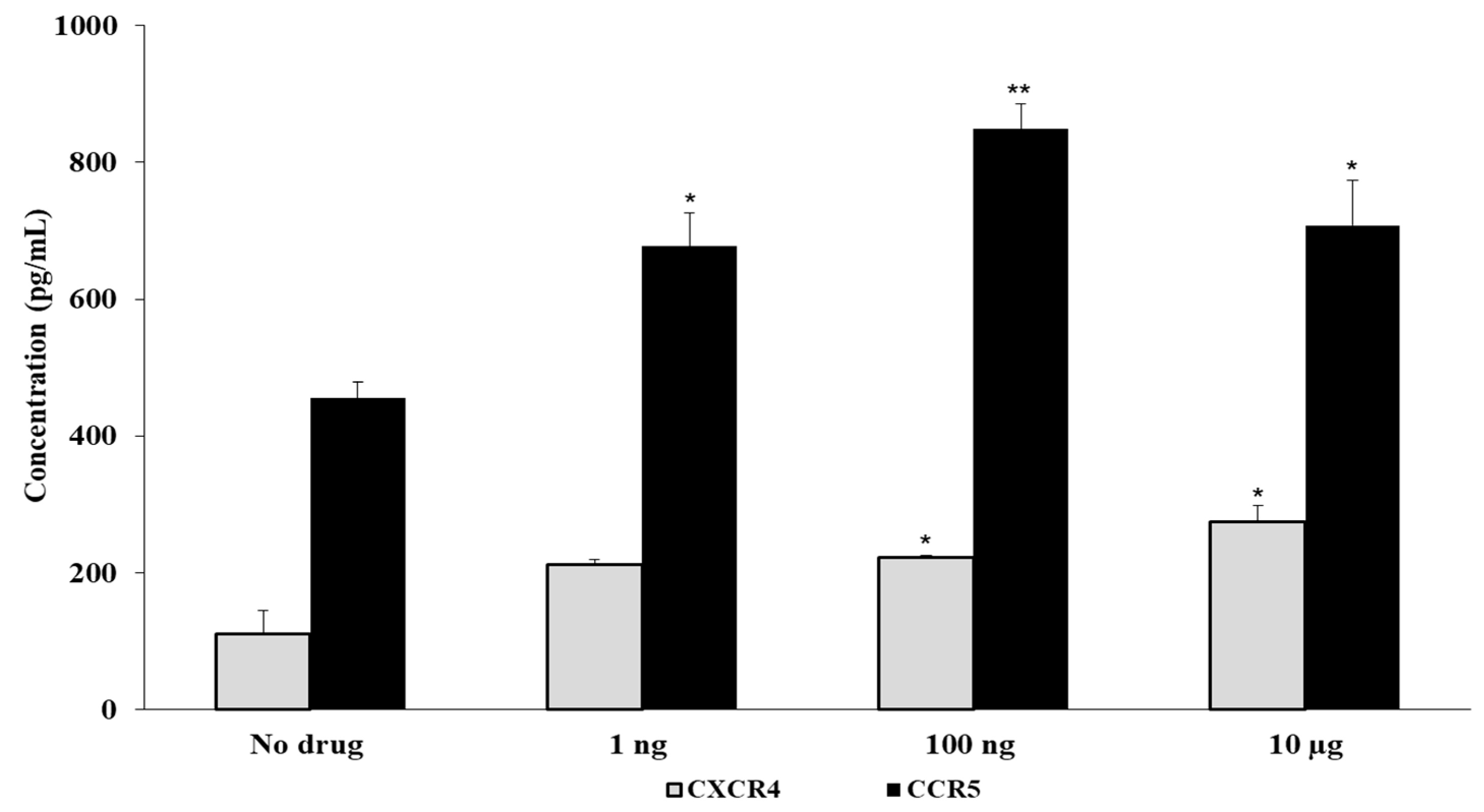
3.3. Fentanyl Increases HIV Co-Receptor Expression
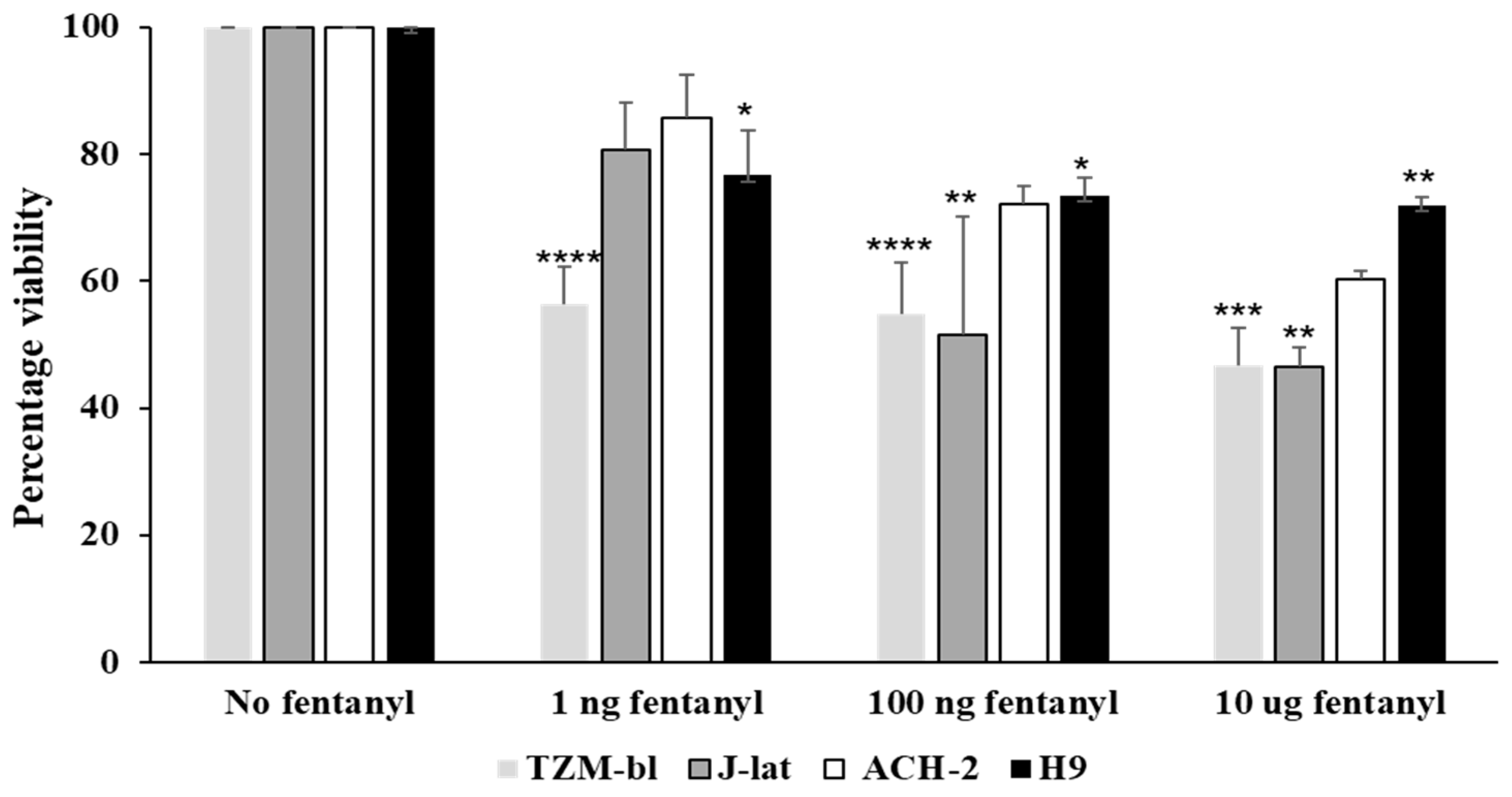
3.4. Fentanyl Alters Cell Viability
3.5. Fentanyl Enhances HIV Replication in Primary PBMC Derived T Cells
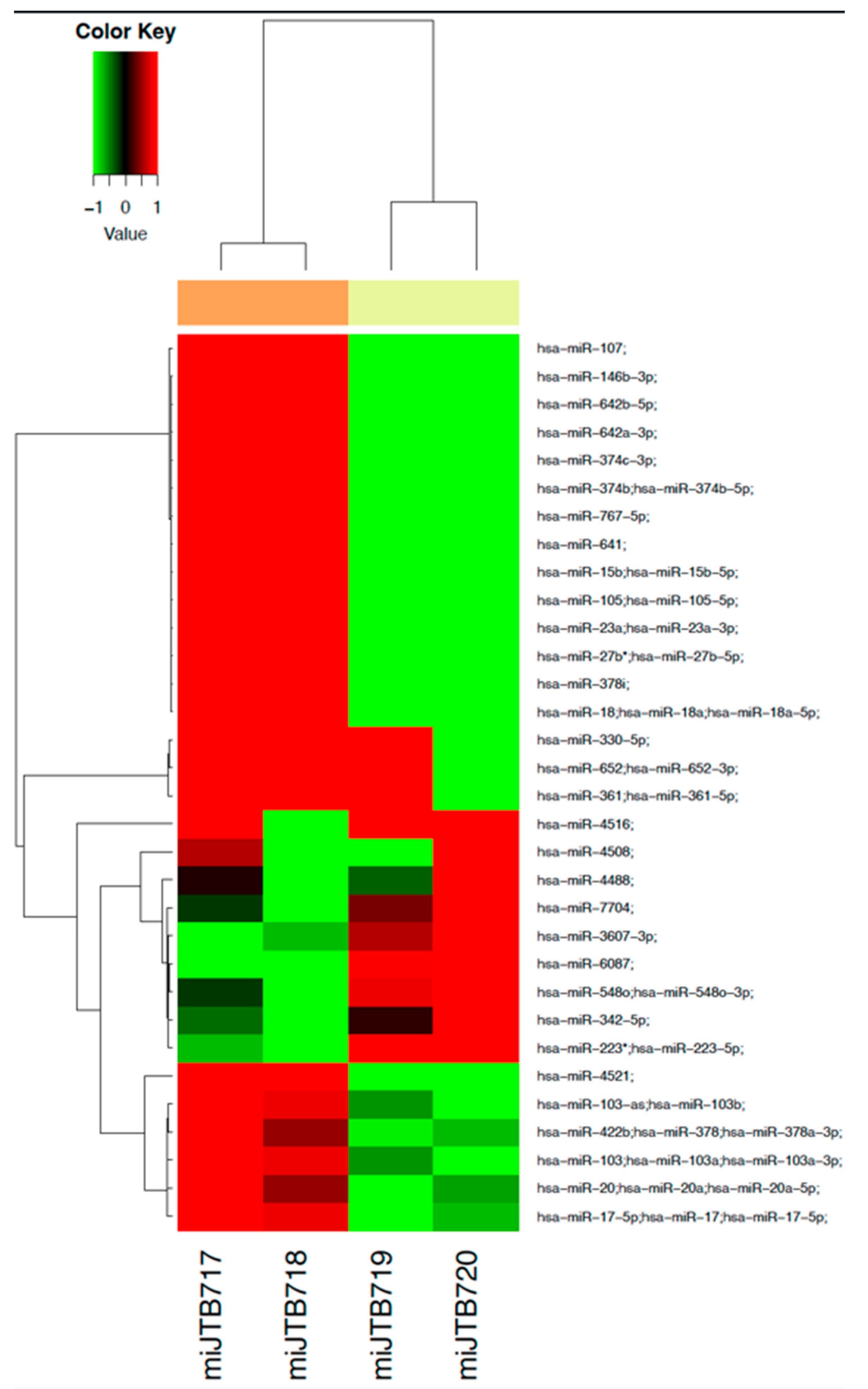
3.6. Dysregulation of miRNA Profile by Fentanyl
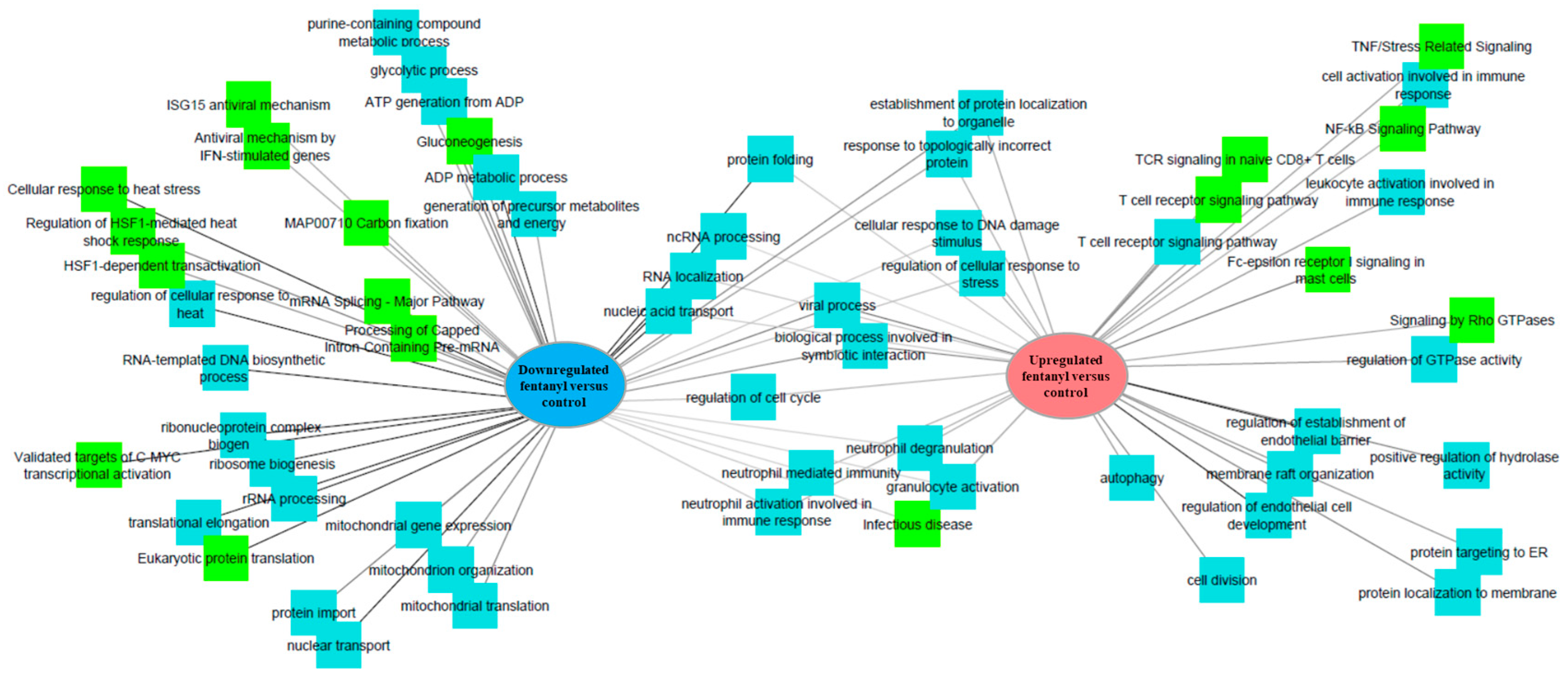
3.7. Fentanyl Alters the Cellular Transcriptome
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- O’Donnell, J.K.; Gladden, R.M.; Seth, P. Trends in deaths involving heroin and synthetic opioids excluding methadone, and law enforcement drug product reports, by census region—United States, 2006–2015. MMWR Morb. Mortal. Wkly. Rep. 2017, 66, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Mattson, C.L.; Tanz, L.J.; Quinn, K.; Kariisa, M.; Patel, P.; Davis, N.L. Trends and geographic patterns in drug and synthetic opioid overdose deaths—United States, 2013–2019. MMWR Morb. Mortal. Wkly. Rep. 2021, 70, 202. [Google Scholar] [CrossRef] [PubMed]
- Daniulaityte, R.; Juhascik, M.P.; Strayer, K.E.; Sizemore, I.E.; Harshbarger, K.E.; Antonides, H.M.; Carlson, R.R. Overdose deaths related to fentanyl and its analogs—Ohio, January–February 2017. MMWR Morb. Mortal. Wkly. Rep. 2017, 66, 904–908. [Google Scholar] [CrossRef] [PubMed]
- Peterson, A.B.; Gladden, R.M.; Delcher, C.; Spies, E.; Garcia-Williams, A.; Wang, Y.; Halpin, J.; Zibbell, J.; McCarty, C.L.; DeFiore-Hyrmer, J.; et al. Increases in fentanyl-related overdose deaths—Florida and Ohio, 2013–2015. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 844–849. [Google Scholar] [CrossRef] [PubMed]
- Strayer, K.E.; Antonides, H.M.; Juhascik, M.P.; Daniulaityte, R.; Sizemore, I.E. LC-MS/MS-based method for the multiplex detection of 24 fentanyl analogues and metabolites in whole blood at sub ng mL-1 concentrations. ACS Omega 2018, 3, 514–523. [Google Scholar] [CrossRef]
- Samji, H.; Yu, A.; Wong, S.; Wilton, J.; Binka, M.; Alvarez, M.; Bartlett, S.; Pearce, M.; Adu, P.; Jeong, D.; et al. Drug-related deaths in a population-level cohort of people living with and without hepatitis C virus in British Columbia, Canada. Int. J. Drug Policy 2020, 86, 102989. [Google Scholar] [CrossRef]
- Banerjee, A.; Strazza, R.; Wigdahl, B.; Pirrone, V.; Meucci, O.; Nonnemacher, M. Role of mu-opioid receptors as cofactors in human immunodeificiency virus type 1 disease progression and neuorpathogenesis. J. Neuroviral. 2011, 17, 291–302. [Google Scholar] [CrossRef]
- Eisenstein, T.K. The role of opioid receptors in immune system function. Front Immunol. 2019, 10, 2904. [Google Scholar] [CrossRef]
- Kraus, J.; Börner, C.; Giannini, E.; Höllt, V. The role of nuclear factor kappaB in tumor necrosis factor-regulated transcription of the human mu-opioid receptor gene. Mol. Pharmacol. 2003, 64, 876–884. [Google Scholar] [CrossRef]
- Plein, L.M.; Rittner, H.L. Opioids and the immune system—Friend or foe. Br. J. Pharmacol. 2018, 175, 2717–2725. [Google Scholar] [CrossRef]
- Liang, X.; Liu, R.; Chen, C.; Ji, F.; Li, T. Opioid system modulates the immune function: A review. Transl. Perioper. Pain Med. 2016, 1, 5–13. [Google Scholar] [PubMed]
- Brejchova, J.; Holan, V.; Svoboda, P. Expression of opioid receptors in cells of the immune system. Int. J. Mol. Sci. 2021, 22, 315. [Google Scholar] [CrossRef] [PubMed]
- Fentanyl Drug Facts. National Institute on Drug Abuse: 2021. Available online: https://www.drugabuse.gov/publications/drugfacts/fentanyl (accessed on 10 January 2022).
- Lambdin, B.H.; Bluthenthal, R.N.; Zibbell, J.E.; Wenger, L.; Simpson, K.; Kral, A.H. Associations between perceived illicit fentanyl use and infectious disease risks among people who inject drugs. Int. J. Drug Policy 2019, 74, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Blackard, J.T.; Brown, J.L.; Lyons, M.S. Synthetic opioid use and common injection-associated viruses: Expanding the translational research agenda. Curr. HIV Res. 2019, 17, 94–101. [Google Scholar] [CrossRef]
- Tahamtan, A.; Tavakoli-Yaraki, M.; Mokhtari-Azad, T.; Teymoori-Rad, M.; Bont, L.; Shokri, F.; Salimi, V. Opioids and viral infections: A double-edged sword. Front. Microbiol. 2016, 7, 970. [Google Scholar] [CrossRef]
- Ellwanger, J.H.; Kaminski, V.D.; Rodrigues, A.G.; Kulmann-Leal, B.; Chies, J.A. CCR5 and CCR5Δ32 in bacterial and parasitic infections: Thinking chemokine receptors outside the HIV box. Int. J. Immunogenet. 2020, 47, 261–285. [Google Scholar] [CrossRef]
- Pozzobon, T.; Goldoni, G.; Viola, A.; Molon, B. CXCR4 signaling in health and disease. Immunol. Lett. 2016, 177, 6–15. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, X.; Ye, L.; Li, J.; Song, L.; Fulambarkar, N.; Ho, W. Morphine Suppresses IFN Signaling Pathway and Enhances AIDS Virus Infection. PLoS ONE 2012, 7, e31167. [Google Scholar] [CrossRef]
- Kopnisky, K.L.; Bao, J.; Lin, Y.W. Neurobiology of HIV, psychiatric and substance abuse comorbidity research: Workshop report. Brain Behav. Immun. 2007, 21, 428–441. [Google Scholar] [CrossRef]
- Guo, C.J.; Li, Y.; Tian, S.; Wang, X.; Douglas, S.D.; Ho, W. Morphine enhances HIV infection of human blood mononuclear phagocytes through modulation of beta-chemokines and CCR5 receptor. J. Investig. Med. 2002, 50, 435–442. [Google Scholar] [CrossRef]
- Li, Y.; Merrill, J.D.; Mooney, K.; Song, L.; Wang, X.; Guo, C.-J.; Savani, R.C.; Metzger, D.S.; Douglas, S.D.; Ho, W.-Z. Morphine Enhances HIV Infection of Neonatal Macrophages. Pediatr. Res. 2003, 54, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Peterson, P.K.; Sharp, B.M.; Gekker, G.; Portoghese, P.S.; Sannerud, K.; Balfour, H.H., Jr. Morphine promotes the growth of HIV-1 in human peripheral blood mononuclear cell cocultures. AIDS 1990, 4, 869–873. [Google Scholar] [CrossRef] [PubMed]
- Schweitzer, C.; Keller, F.; Schmitt, M.P.; Jaeck, D.; Adloff, M.; Schmitt, C.; Royer, C.; Kirn, A.; Aubertin, A. Morphine stimulates HIV replication in primary cultures of human Kupffer cells. Res. Virol. 1991, 142, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Chuang, A.J.; Chuang, L.F.; Doi, R.H.; Chuang, R.Y. Morphine promotes simian acquired immunodeficiency syndrome virus replication in monkey peripheral mononuclear cells: Induction of CC chemokine receptor 5 expression for virus entry. J. Infect. Dis. 2002, 185, 1826–1829. [Google Scholar] [CrossRef]
- Peterson, P.K.; Gekker, G.; Hu, S.; Anderson, W.; Kravitz, F.; Portoghese, P.S.; Balfour, H.H.; Chao, C.C. Morphine amplifies HIV-1 expression in chronically infected promonocytes cocultured with human brain cells. J. Neuroimmunol. 1994, 50, 167–175. [Google Scholar] [CrossRef]
- Nair, M.P.; Schwartz, S.A.; Polasani, R.; Hou, J.; Sweet, A.; Chadha, K.C. Immunoregulatory effects of morphine on human lymphocytes. Clin. Diagn. Lab. Immunol. 1997, 4, 127–132. [Google Scholar] [CrossRef]
- El-Hage, N.; Wu, G.; Wang, J.; Ambati, J.; Knapp, P.E.; Reed, J.L.; Bruce-Keller, A.J.; Hauser, K.F. HIV-1 Tat and opiate-induced changes in astrocytes promote chemotaxis of microglia through the expression of MCP-1 and alternative chemokines. Glia 2006, 53, 132–146. [Google Scholar] [CrossRef]
- Wang, X.; Tan, N.; Douglas, S.D.; Zhang, T.; Wang, Y.J.; Ho, W.Z. Morphine inhibits CD8+ T cell-mediated, noncytolytic, anti-HIV activity in latently infected immune cells. J. Leukoc. Biol. 2005, 78, 772–776. [Google Scholar] [CrossRef]
- Squinto, S.P.; Mondal, D.; Block, A.L.; Prakash, O. Morphine-Induced Transactivation of HIV-1 LTR in Human Neuroblastoma Cells. AIDS Res. Hum. Retrovir. 1990, 6, 1163–1168. [Google Scholar] [CrossRef]
- Nair, M.P.N.; Mahajan, S.; Chadha, K.C.; Nair, N.M.; Hewitt, R.G.; Pillai, S.K.; Chadha, P.; Sukumaran, P.C.; Schwartz, S.A. Effect of cocaine on chemokine and CCR-5 gene expression by mononuclear cells from normal donors and HIV-1 infected patients. Adv. Exp. Med. Biol. 2001, 493, 235–240. [Google Scholar]
- Jiang, J.; Wang, M.; Liang, B.; Shi, Y.; Su, Q.; Chen, H.; Huang, J.; Su, J.; Pan, P.; Li, Y.; et al. In vivo effects of methamphetamine on HIV-1 replication: A population-based study. Drug Alcohol Depend. 2016, 159, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Derdeyn, C.A.; Decker, J.M.; Sfakianos, J.N.; Wu, X.; O’Brien, W.A.; Ratner, L.; Kappes, J.C.; Shaw, G.M.; Hunter, E. Sensitivity of human immunodeficiency virus type 1 to the fusion inhibitor T-20 is modulated by coreceptor specificity defined by the V3 loop of gp120. J. Virol. 2000, 74, 8358–8367. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, Y.; McClure, M.O.; Pizzato, M. Identification of gamma retroviruses constitutively released from cell lines used for human immunodeficiency virus research. J. Virol. 2008, 82, 12585–12588. [Google Scholar] [CrossRef] [PubMed]
- Jordan, A.; Bisgrove, D.; Verdin, E. HIV reproducibly establishes a latent infection after acute infection of T cells in vitro. EMBO J. 2003, 22, 1868–1877. [Google Scholar] [CrossRef] [PubMed]
- Jordan, A.; Defechereux, P.; Verdin, E. The site of HIV-1 integration in the human genome determines basal transcriptional activity and response to Tat transactivation. EMBO J. 2001, 20, 1726–1738. [Google Scholar] [CrossRef] [PubMed]
- Mann, D.L.; O’Brien, S.J.; Gilbert, D.A.; Reid, Y.; Popovic, M.; Read-Connole, E.; Gallo, R.C.; Gazdar, A.F. Origin of the HIV-susceptible human CD4+ cell line H9. AIDS Res. Hum. Retrovir. 1989, 5, 253–255. [Google Scholar] [CrossRef]
- Clouse, K.A.; Powell, D.; Washington, I.; Poli, G.; Strebel, K.; Farrar, W.; Barstad, P.; Kovacs, J.; Fauci, A.S.; Folks, T.M. Monokine regulation of human immunodeficiency virus-1 expression in a chronically infected human T cell clone. J. Immunol. 1989, 142, 431–438. [Google Scholar] [CrossRef]
- Armenian, P.; Vo, K.T.; Barr-Walker, J.; Lynch, K.L. Fentanyl, fentanyl analogs and novel synthetic opioids: A comprehensive review. Neuropharmacology 2018, 134, 121–132. [Google Scholar] [CrossRef]
- Gibellini, D.; Vitone, F.; Schiavone, P.; Ponti, C.; La Placa, M.; Re, M.C. Quantitative detection of human immunodeficiency virus type 1 (HIV-1) proviral DNA in peripheral blood mononuclear cells by SYBR green real-time PCR technique. J. Clin. Virol. 2004, 29, 282–289. [Google Scholar] [CrossRef]
- Langevin, S.M.; Kuhnell, D.; Orr-Asman, M.A.; Biesiada, J.; Zhang, X.; Medvedovic, M.; Thomas, H.E. Balancing yield, purity and practicality: A modified differential ultracentrifugation protocol for efficient isolation of small extracellular vesicles from human serum. RNA Biol. 2019, 16, 5–12. [Google Scholar] [CrossRef]
- Paradis, E.; Schliep, K. ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Huber, W.; Carey, V.J.; Gentleman, R.; Anders, S.; Carlson, M.; Carvalho, B.S.; Bravo, H.C.; Davis, S.; Gatto, L.; Girke, T.; et al. Orchestrating high-throughput genomic analysis with Bioconductor. Nat. Methods 2015, 12, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Kozomara, A.; Griffiths-Jones, S. miRBase: Annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. 2014, 42, 68–73. [Google Scholar] [CrossRef]
- Robinson, M.; McCarthy, D.; Smyth, G.K. edgeR: Differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Walsh, K.B.; Zhang, X.; Zhu, X.; Wohleb, E.; Woo, D.; Lu, L.; Adeoye, O. Intracerebral hemorrhage induces inflammatory gene expression in peripheral blood: Global transcriptional profiling in intracerebral hemorrhage patients. DNA Cell Biol. 2019, 38, 660–669. [Google Scholar] [CrossRef]
- Rapp, S.J.; Dershem, V.; Zhang, X.; Schutte, S.C.; Chariker, M.E. Varying negative pressure wound therapy acute effects on human split-thickness autografts. J. Burn. Care Res. 2020, 41, 104–112. [Google Scholar] [CrossRef]
- U.S. Overdose Deaths In 2021 Increased Half as Much as in 2020—But Are Still Up 15% 2022.2022). Available online: https://www.cdc.gov/nchs/pressroom/nchs_press_releases/2022/202205.htm (accessed on 11 May 2022).
- Lyden, J.; Binswanger, I.A. The United States opioid epidemic. Semin. Perinatol. 2019, 43, 123–131. [Google Scholar] [CrossRef]
- Lavonas, E.J.; Dezfulian, C. Impact of the Opioid Epidemic. Crit. Care Clin. 2020, 36, 753–769. [Google Scholar] [CrossRef]
- Reider, B. Opioid epidemic. Am. J. Sports Med. 2019, 47, 1039–1042. [Google Scholar] [CrossRef]
- Volkow, N.D.; Blanco, C. The changing opioid crisis: Development, challenges and opportunities. Mol. Psychiatry 2021, 26, 218–233. [Google Scholar] [CrossRef] [PubMed]
- Csete, J.; Kamarulzaman, A.; Kazatchkine, M.; Altice, F.L.; Balicki, M.; Buxton, J.; Cepeda, J.; Comfort, M.; Goosby, E.; Goulão, J.; et al. Public health and international drug policy. Lancet 2016, 387, 1427–1480. [Google Scholar] [CrossRef] [PubMed]
- Ball, L.J.; Puka, K.; Speechley, M.; Wong, R.; Hallam, B.; Wiener, J.C.; Koivu, S.; Silverman, M.S. Sharing of injection drug preparation equipment is associated with HIV infection: A cross-sectional study. J. Acquir. Immune Defic. Syndr. 2019, 81, e99–e103. [Google Scholar] [CrossRef]
- Bulled, N.; Singer, M. Syringe-mediated syndemics. AIDS Behav. 2011, 15, 1539–1545. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, M.; Weber, J.; Bukrinsky, M.; Simon, G.L. The effects of cocaine on HIV transcription. J. Neurovirol. 2016, 22, 261–274. [Google Scholar] [CrossRef]
- Moradi, G.; Hajarizadeh, B.; Rahmani, K.; Mohamadi-Bolbanabad, A.; Darvishi, S.; Zareie, B.; Zavareh, F.A.; Sharafi, H.; Alavian, S.M.; Ramazani, R.; et al. Drug use and risk behaviour profile, and the prevalence of HIV, hepatitis C and hepatitis B among people with methamphetamine use in Iran. Int. J. Drug Policy 2019, 73, 129–134. [Google Scholar] [CrossRef]
- Degenhardt, L.; Peacock, A.; Colledge, S.; Leung, J.; Grebely, J.; Vickerman, P.; Stone, J.; Cunningham, E.B.; Trickey, A.; Dumchev, K.; et al. Global prevalence of injecting drug use and sociodemographic characteristics and prevalence of HIV, HBV, and HCV in people who inject drugs: A multistage systematic review. Lancet Glob. Health 2017, 5, e1192–e1207. [Google Scholar] [CrossRef]
- Illenberger, J.M.; Harrod, S.B.; Mactutus, C.F.; McLaurin, K.A.; Kallianpur, A.; Booze, R.M. HIV Infection and Neurocognitive Disorders in the Context of Chronic Drug Abuse: Evidence for Divergent Findings Dependent upon Prior Drug History. J. Neuroimmune Pharmacol. 2020, 15, 715–728. [Google Scholar] [CrossRef]
- Cernasev, A.; Veve, M.P.; Cory, T.J.; Summers, N.A.; Miller, M.; Kodidela, S.; Kumar, S. Opioid Use Disorders in People Living with HIV/AIDS: A Review of Implications for Patient Outcomes, Drug Interactions, and Neurocognitive Disorders. Pharmacy 2020, 8, 168. [Google Scholar] [CrossRef]
- Strain, E. Opioid Use Disorder: Epidemiology, Pharmacology, Clinical Manifestations, Course, Screening, Assessment, and Diagnosis; Saxon, A., Hermann, R., Eds.; Wolters Kluwer, UpToDate: Waltham, MA, USA, 2018. [Google Scholar]
- Weisberg, D.F.; Gordon, K.S.; Barry, D.T.; Becker, W.C.; Crystal, S.; Edelman, E.J.; Gaither, J.; Gordon, A.J.; Goulet, J.; Kerns, R.D.; et al. Long-term Prescription of Opioids and/or Benzodiazepines and Mortality among HIV-Infected and Uninfected Patients. J Acquir. Immune Defic. Syndr. 2015, 69, 223–233. [Google Scholar] [CrossRef]
- Sundar, K.S.; Kamaraju, L.S.; Dingfelder, J.; McMahon, J.; Gollapudi, S.; Wilson, W.H.; Kong, L.Y.; Hong, J.S.; Weiss, J.M.; Lee, J. beta-Endorphin enhances the replication of neurotropic human immunodeficiency virus in fetal perivascular microglia. J. Neuroimmunol. 1995, 61, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Peterson, P.K.; Gekker, G.; Hu, S.; Lokensgard, J.; Portoghese, P.S.; Chao, C. Endomorphin-1 potentiates HIV-1 expression in human brain cell cultures: Implication of an atypical mu-opioid receptor. Neuropharmacology 1999, 38, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.C.; Gekker, G.; Hu, S.; Sheng, W.S.; Portoghese, P.S.; Peterson, P.K. Upregulation of HIV-1 expression in cocultures of chronically infected promonocytes and human brain cells by dynorphin. Biochem. Pharmacol. 1995, 50, 715–722. [Google Scholar] [CrossRef] [PubMed]
- Prottengeier, J.; Koutsilieri, E.; Scheller, C. The effects of opioids on HIV reactivation in latently-infected T-lymphoblasts. AIDS Res. Ther. 2014, 11, 17. [Google Scholar] [CrossRef]
- Xu, D.; Fu, J.; Jin, L.; Zhang, H.; Zhou, C.; Zou, Z.; Zhao, J.-M.; Zhang, B.; Shi, M.; Ding, X.; et al. Circulating and Liver Resident CD4+CD25+ Regulatory T Cells Actively Influence the Antiviral Immune Response and Disease Progression in Patients with Hepatitis B. J. Immunol. 2006, 177, 739–747. [Google Scholar] [CrossRef]
- Wang, X.; Liu, J.; Zhou, L.; Ho, W.-Z. MorphineWithdrawal Enhances HIV Infection of Macrophages. Front. Immunol. 2019, 10, 2601. [Google Scholar] [CrossRef]
- Tang, B.; Zhang, Y.; Liang, R.; Yuan, P.; Du, J.; Wang, H.; Wang, L. Activation of the -opioid receptor inhibits serum deprivation induced apoptosis of human liver cells via the activation of PKC and the mitochondrial pathway. Int. J. Mol. Med. 2011, 28, 1077–1085. [Google Scholar]
- Wang, X.; Ma, T.-C.; Li, J.-L.; Zhou, Y.; Geller, E.B.; Adler, M.W.; Peng, J.-S.; Zhou, W.; Zhou, D.-J.; Ho, W.-Z. Heroin inhibits HIV-restriction miRNAs and enhances HIV infection of macrophages. Front. Microbiol. 2015, 6, 1230. [Google Scholar] [CrossRef]
- Wang, X.; Sun, L.; Zhou, Y.; Su, Q.-J.; Li, J.-L.; Ye, L.; Liu, M.-Q.; Zhou, W.; Ho, W.-Z. Heroin Abuse and/or HIV Infection Dysregulate Plasma Exosomal miRNAs. J. Neuroimmune Pharmacol. 2020, 15, 400–408. [Google Scholar] [CrossRef]
- Gornalusse, G.G.; Vojtech, L.N.; Levy, C.N.; Hughes, S.M.; Kim, Y.; Valdez, R.; Pandey, U.; Ochsenbauer, C.; Astronomo, R.; McElrath, J.; et al. Buprenorphine Increases HIV-1 Infection In Vitro but Does Not Reactivate HIV-1 from Latency. Viruses 2021, 13, 1472. [Google Scholar] [CrossRef]
- Wang, X.; Tian, S.; Guo, C.J.; Douglas, S.D.; Ho, W. Methadone enhances human immunodeficiency virus infection of human immune cells. J. Infect. Dis. 2002, 185, 118–122. [Google Scholar]
- Wang, M.-R.; Wu, D.-D.; Luo, F.; Zhong, C.-J.; Wang, X.; Zhu, N.; Wu, Y.-J.; Hu, H.-T.; Feng, Y.; Wang, X.; et al. Methadone Inhibits Viral Restriction Factors and Facilitates HIV Infection in Macrophages. Front. Immunol. 2020, 11, 1253. [Google Scholar] [CrossRef] [PubMed]
- Facts about Fentanyl. DEA. In United States Drug Enforcement Administration: 2021. Available online: https://www.dea.gov/resources/facts-about-fentany (accessed on 20 March 2023).
- Woodall, K.L.; Martin, T.L.; McLellan, B.A. Oral abuse of fentanyl patches (Duragesic®): Seven case reports. J. Forensic Sci. 2008, 53, 222–225. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Slawson, M.H.; Johnson-Davis, K.L. Metabolic patterns of fentanyl, meperidine, methylphenidate, tapentadol and tramadol observed in urine, serum or plasma. J. Anal. Toxicol. 2017, 41, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Karns, R.; Shata, M.; Brown, J.L.; Lyons, M.S.; Sherman, K.E.; Blackard, J.T. The synthetic opioid fentanyl enhances viral replication in vitro. PLoS ONE 2021, 16, e0249581. [Google Scholar] [CrossRef]
- Tang, B.; Li, Y.; Yuan, S.; Tomlinson, S.; He, S. Upregulation of the opioid receptor in liver cancer promotes liver cancer progression both in vitro and in vivo. Int. J. Oncol. 2013, 43, 1281–1290. [Google Scholar] [CrossRef]
- Lu, J.; Liu, Z.; Zhao, L.; Tian, H.; Liu, X.; Yuan, C. In vivo and in vitro inhibition of human liver cancer progress by downregulation of the μ-opioid receptor and relevant mechanisms. Oncol. Rep. 2013, 30, 1731–1738. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, L.-C.; Chen, E.-Q.; Chen, X.-B.; Chen, L.-Y.; Liu, L.; Lei, X.-Z.; Liu, C.; Tang, H. Prospective Evaluation of FibroScan for the Diagnosis of Hepatic Fibrosis Compared with Liver Biopsy/AST Platelet Ratio Index and FIB-4 in Patients with Chronic HBV Infection. Dig. Dis. Sci. 2011, 56, 2742–2749. [Google Scholar] [CrossRef]
- Jia, K.; Sun, D.; Ling, S.; Tian, Y.; Yang, X.; Sui, J.; Tang, B.; Wang, L. Activated-opioid receptors inhibit hydrogen peroxideinduced apoptosis in liver cancer cells through the PKC/ERK signaling pathwa. Mol. Med. Rep. 2014, 10, 839–847. [Google Scholar] [CrossRef]
- De Minicis, S.; Candelaresi, C.; Marzioni, M.; Saccomano, S.; Roskams, T.; Casini, A.; Risaliti, A.; Salzano, R.; Cautero, N.; di Francesco, F.; et al. Role of endogenous opioids in modulating HSC activity in vitro and liver fibrosis in vivo. Gut 2008, 57, 352–364. [Google Scholar] [CrossRef]
- Ye, L.; Peng, J.S.; Wang, X.; Wang, Y.J.; Luo, G.X.; Ho, W.Z. Methamphetamine enhances Hepatitis C virus replication in human hepatocytes. J. Viral Hepat. 2008, 15, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.-Y.; Chang, M.-H.; Ni, Y.-H.; Chen, H. Survey of hepatitis B surface variant infection in children 15 years after a nationwide vaccination programme in Taiwan. Gut 2004, 53, 1499–1503. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Wang, X.; Chen, H.; Song, L.; Ye, L.; Wang, S.-H.; Wang, Y.-J.; Zhou, L.; Ho, W.-Z. Methamphetamine Enhances HIV Infection of Macrophages. Am. J. Pathol. 2008, 172, 1617–1624. [Google Scholar] [CrossRef]
- Liu, X.; Zha, J.; Nishitani, J.; Chen, H.; Zack, J.A. HIV-1 infection in peripheral blood lymphocytes (PBLs) exposed to alcohol. Virology 2003, 307, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Nair, M.P.; Chadha, K.C.; Hewitt, R.G.; Mahajan, S.; Sweet, A.; Schwartz, S.A. Cocaine differentially modulates chemokine production by mononuclear cells from normal donors and human immunodeficiency virus type 1-infected patients. Clin. Diagn. Lab. Immunol. 2000, 7, 96–100. [Google Scholar] [CrossRef]
- Nair, M.P.; Saiyed, Z.M. Effect of methamphetamine on expression of HIV coreceptors and CC-chemokines by dendritic cells. Life Sci. 2011, 88, 987–994. [Google Scholar] [CrossRef]
- Nair, M.P.; Saiyed, Z.M.; Nair, N.; Gandhi, N.H.; Rodriguez, J.W.; Boukli, N.; Provencio-Vasquez, E.; Malow, R.M.; Miguez-Burbano, M. Methamphetamine enhances HIV-1 infectivity in monocyte derived dendritic cells. J. Neuroimmune Pharmacol. 2009, 4, 129–139. [Google Scholar] [CrossRef]
- Roth, M.D.; Whittaker, K.M.; Choi, R.; Tashkin, D.P.; Baldwin, G.C. Cocaine and -1 receptors modulate HIV infection, chemokine receptors, and the HPA axis in the huPBL-SCID model. J. Leukoc. Biol. 2005, 78, 1198–1203. [Google Scholar] [CrossRef]
- Wang, X.; Douglas, S.D.; Metzger, D.S.; Guo, C.; Li, Y.; O’Brien, C.P.; Song, L.; Davis-Vogal, A.; Ho, W. Alcohol potentiates HIV-1 infection of human blood mononuclear phagocytes. Alcohol. Clin. Exp. Res. 2002, 26, 1880–1886. [Google Scholar] [CrossRef]
- Miyagi, T.; Chuang, L.F.; Doi, R.H.; Carlos, M.P.; Torres, J.V.; Chuang, R.Y. Morphine induces gene expression of CCR5 in human CEM x174 lymphocytes. J. Biol. Chem. 2000, 275, 31305–31310. [Google Scholar] [CrossRef]
- Steele, A.D.; Szabo, I.; Bednar, F.; Rogers, T.J. Interactions between opioid and chemokine receptors: Heterologous desensitization. Cytokine Growth Factor Rev. 2002, 13, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Shata, M.T.M.; Brown, J.L.; Lyons, M.S.; Sherman, K.E.; Blackard, J.T. The synthetic opioid fentanyl increases HIV replication and chemokine co-receptor expression in vitro. J. NeuroVirology 2022, 28, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Nie, D.-H.; Bai, C.-S.; Rehman, A.; Yang, A.; Mou, X.-L.; Zhang, Y.-Q.; Xu, Y.-Q.; Xiang, Q.-Q.; Ren, Y.-T.; et al. Fentanyl enhances HIV infection in vitro. Virology 2022, 577, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Ma, P.; Lu, H.; Liu, S.; Cao, Q. Fentanyl Suppresses the Survival of CD4+ T Cells Isolated from Human Umbilical Cord Blood through Inhibition of IKK s-mediated NF-κB Activation. Scand. J. Immunol. 2017, 85, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Kandathil, A.J.; Sugawara, S.; Balagopal, A. Are T cells the only HIV-1 reservoir? Retrovirology 2016, 13, 86. [Google Scholar] [CrossRef]
- Su, B.; Fu, Y.; Liu, Y.; Wu, H.; Ma, P.; Zeng, W.; Zhang, T.; Lian, S.; Wu, H. Potential Application of MicroRNA Profiling to the Diagnosis and Prognosis of HIV-1 Infection. Front. Microbiol. 2018, 9, 3185. [Google Scholar] [CrossRef]
- Triboulet, R.; Mari, B.; Lin, Y.L.; Chable-Bessia, C.; Bennasser, Y.; Lebrigand, K.; Cardinaud, B.; Maurin, T.; Barbry, P.; Baillat, V.; et al. Suppression of microRNA-silencing pathway by HIV-1 during virus replication. Science. 2007, 315, 1579–1582. [Google Scholar] [CrossRef]
- Sung, T.-L.; Rice, A.P. miR-198 Inhibits HIV-1 Gene Expression and Replication in Monocytes and Its Mechanism of Action Appears To Involve Repression of Cyclin T1. PLOS Pathog. 2009, 5, e1000263. [Google Scholar] [CrossRef]
- Chiang, K.; Liu, H.; Rice, A.P. miR-132 enhances HIV-1 replication. Virology 2013, 438, 1–4. [Google Scholar] [CrossRef]
- Hariharan, M.; Scaria, V.; Pillai, B.; Brahmachari, S.K. Targets for human encoded microRNAs in HIV genes. Biochem. Biophys. Res. Commun. 2005, 337, 1214–1218. [Google Scholar] [CrossRef]
- Pilakka-Kanthikeel, S.; Nair, M.P.N. Interaction of drugs of abuse and microRNA with HIV: A brief review. Front. Microbiol. 2015, 6, 967. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Cheng, L.; Höxtermann, S.; Xie, T.; Lu, X.; Wu, H.; Skaletz-Rorowski, A.; Brockmeyer, N.H.; Wu, N. Micro RNA-155 is a biomarker of T-cell activation and immune dysfunction in HIV-1-infected patients. HIV Med. 2017, 18, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.-S.; Ott, M. The ups and downs of SIRT1. Trends Biochem Sci. 2008, 33, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Gorroño, L.E.; Escribà, T.; Boulanger, N.; Guardo, A.C.; Leon, A.; Bargalló, M.E.; García, F.; Gatell, J.M.; Plana, M.; Arnedo, M.; et al. Differential MicroRNA Expression Profile between Stimulated PBMCs from HIV-1 Infected Elite Controllers and Viremic Progressors. PLOS ONE 2014, 9, e106360. [Google Scholar] [CrossRef]
- Houzet, L.; Klase, Z.; Yeung, M.L.; Wu, A.; Le, S.Y.; Quinones, M.; Jeang, K.T. The extent of sequence complementarity correlates with the potency of cellular miRNA-mediated restriction of HIV-1. Nucleic Acids Res. 2012, 40, 11684–11696. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.K.; Sengupta, P.; Waki, K.; Van Engelenburg, S.B.; Ochiya, T.; Ablan, S.D.; Freed, E.O.; Lippincott-Schwartz, J. MicroRNA binding to the HIV-1 Gag protein inhibits Gag assembly and virus production. Proc. Natl. Acad. Sci. 2014, 111, E2676–E2683. [Google Scholar] [CrossRef]
- Wang, P.; Qu, X.; Zhou, X.; Shen, Y.; Ji, H.; Fu, Z.; Deng, J.; Lu, P.; Yu, W.; Lu, H.; et al. Two cellular microRNAs, miR-196b and miR-1290, contribute to HIV-1 latency. Virology 2015, 486, 228–238. [Google Scholar] [CrossRef]
- Huang, J.; Wang, F.; Argyris, E.; Chen, K.; Liang, Z.; Tian, H.; Huang, W.; Squires, K.; Verlinghieri, G.; Zhang, H. Cellular microRNAs contribute to HIV-1 latency in resting primary CD4+ T lymphocytes. Nat. Med. 2007, 13, 1241–1247. [Google Scholar] [CrossRef]
- Egaña-Gorroño, L.; Guardo, A.C.; Bargalló, M.E.; Planet, E.; Vilaplana, E.; Escribà, T.; Pérez, I.; Gatell, J.M.; García, F.; Arnedo, M.; et al. MicroRNA Profile in CD8+ T-Lymphocytes from HIV-Infected Individuals: Relationship with Antiviral Immune Response and Disease Progression. PLOS ONE 2016, 11, e0155245. [Google Scholar] [CrossRef]
- Asahchop, E.L.; Akinwumi, S.M.; Branton, W.G.; Fujiwara, E.; Gill, M.J.; Power, C. Plasma microRNA profiling predicts HIV-associated neurocognitive disorder. Aids 2016, 30, 2021–2031. [Google Scholar] [CrossRef]
- Dey, R.; Soni, K.; Saravanan, S.; Balakrishnan, P.; Kumar, V.; Boobalan, J.; Solomon, S.S.; Scaria, V.; Solomon, S.; Brahmachari, S.K.; et al. Anti-HIV microRNA expression in a novel Indian cohort. Sci. Rep. 2016, 6, 28279. [Google Scholar] [CrossRef] [PubMed]
- Pilakka-Kanthikeel, S.; Raymond, A.; Atluri, V.S.R.; Sagar, V.; Saxena, S.K.; Diaz, P.; Chevelon, S.; Concepcion, M.; Nair, M. Sterile alpha motif and histidine/aspartic acid domain-containing protein 1 (SAMHD1)-facilitated HIV restriction in astrocytes is regulated by miRNA-181a. J. Neuroinflammation 2015, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Yang, Z.; Jin, C.; Chen, C.; Wu, N. hsa-miR-191-5p inhibits replication of human immunodeficiency virus type 1 by downregulating the expression of NUP50. Arch. Virol. 2021, 166, 755–766. [Google Scholar] [CrossRef] [PubMed]
- Nathans, R.; Chu, C.-Y.; Serquina, A.K.; Lu, C.-C.; Cao, H.; Rana, T.M. Cellular MicroRNA and P Bodies Modulate Host-HIV-1 Interactions. Mol. Cell 2009, 34, 696–709. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Li, H.; Wu, X.; Covarrubias, M.; Scherer, L.; Meinking, K.; Luk, B.; Chomchan, P.; Alluin, J.; Gombart, A.F.; et al. Interplay between HIV-1 infection and host microRNAs. Nucleic Acids Res. 2011, 40, 2181–2196. [Google Scholar] [CrossRef]
- Reynoso, R.; Laufer, N.; Hackl, M.; Skalicky, S.; Monteforte, R.; Turk, G.; Carobene, M.; Quarleri, J.; Cahn, P.; Werner, R.; et al. MicroRNAs differentially present in the plasma of HIV elite controllers reduce HIV infection in vitro. Sci. Rep. 2014, 4, srep05915. [Google Scholar] [CrossRef]
- Huang, J.; Lai, J.; Liang, B.; Jiang, J.; Ning, C.; Liao, Y.; Zang, N.; Wang, M.; Qin, F.; Yu, J.; et al. mircoRNA-3162-3p is a potential biomarker to identify new infections in HIV-1-infected patients. Gene 2018, 662, 21–27. [Google Scholar] [CrossRef]
- Chang, S.T.; Thomas, M.J.; Sova, P.; Green, R.R.; Palermo, R.E.; Katze, M.G. Next-Generation Sequencing of Small RNAs from HIV-Infected Cells Identifies Phased microRNA Expression Patterns and Candidate Novel microRNAs Differentially Expressed upon Infection. Mbio 2013, 4, e00549-12. [Google Scholar] [CrossRef]
- Whisnant, A.; Bogerd, H.P.; Flores, O.; Ho, P.; Powers, J.G.; Sharova, N.; Stevenson, M.; Chen, C.-H.; Cullen, B.R. In-Depth Analysis of the Interaction of HIV-1 with Cellular microRNA Biogenesis and Effector Mechanisms. Mbio 2013, 4, e00193-13. [Google Scholar] [CrossRef]
- Kaul, D.; Ahlawat, A.; Gupta, S.D. HIV-1 genome-encoded hiv1-mir-H1 impairs cellular responses to infection. Mol. Cell. Biochem. 2009, 323, 143–148. [Google Scholar] [CrossRef]
- Zhang, Y.; Fan, M.; Geng, G.; Liu, B.; Huang, Z.; Luo, H.; Zhou, J.; Guo, X.; Cai, W.; Zhang, H. A novel HIV-1-encoded microRNA enhances its viral replication by targeting the TATA box region. Retrovirology 2014, 11, 23. [Google Scholar] [CrossRef] [PubMed]
- Omoto, S.; Ito, M.; Tsutsumi, Y.; Ichikawa, Y.; Okuyama, H.; Brisibe, E.A.; Saksena, N.K.; Fujii, Y.R. HIV-1 nef suppression by virally encoded microRNA. Retrovirology 2004, 1, 44. [Google Scholar] [CrossRef] [PubMed]
- Harwig, A.; Jongejan, A.; van Kampen, A.H.; Berkhout, B.; Das, A.T. Tat-dependent production of an HIV-1 TAR-encoded miRNA-like small RNA. Nucleic Acids Res. 2016, 44, 4340–4353. [Google Scholar] [CrossRef] [PubMed]
- Duskova, K.; Nagilla, P.; Le, H.-S.; Iyer, P.; Thalamuthu, A.; Martinson, J.; Bar-Joseph, Z.; Buchanan, W.; Rinaldo, C.; Ayyavo, V. MicroRNA regulation and its effects on cellular transcriptome in Human Immunodeficiency Virus-1 (HIV-1) infected individuals with distinct viral load and CD4 cell counts. BMC Infect. Dis. 2013, 13, 1–18. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Madhuravasal Krishnan, J.; Kong, L.; Karns, R.; Medvedovic, M.; Sherman, K.E.; Blackard, J.T. The Synthetic Opioid Fentanyl Increases HIV Replication and Chemokine Co-Receptor Expression in Lymphocyte Cell Lines. Viruses 2023, 15, 1027. https://doi.org/10.3390/v15041027
Madhuravasal Krishnan J, Kong L, Karns R, Medvedovic M, Sherman KE, Blackard JT. The Synthetic Opioid Fentanyl Increases HIV Replication and Chemokine Co-Receptor Expression in Lymphocyte Cell Lines. Viruses. 2023; 15(4):1027. https://doi.org/10.3390/v15041027
Chicago/Turabian StyleMadhuravasal Krishnan, Janani, Ling Kong, Rebekah Karns, Mario Medvedovic, Kenneth E. Sherman, and Jason T. Blackard. 2023. "The Synthetic Opioid Fentanyl Increases HIV Replication and Chemokine Co-Receptor Expression in Lymphocyte Cell Lines" Viruses 15, no. 4: 1027. https://doi.org/10.3390/v15041027