
HIV-1 Non-Group M Strains and ART

Abstract
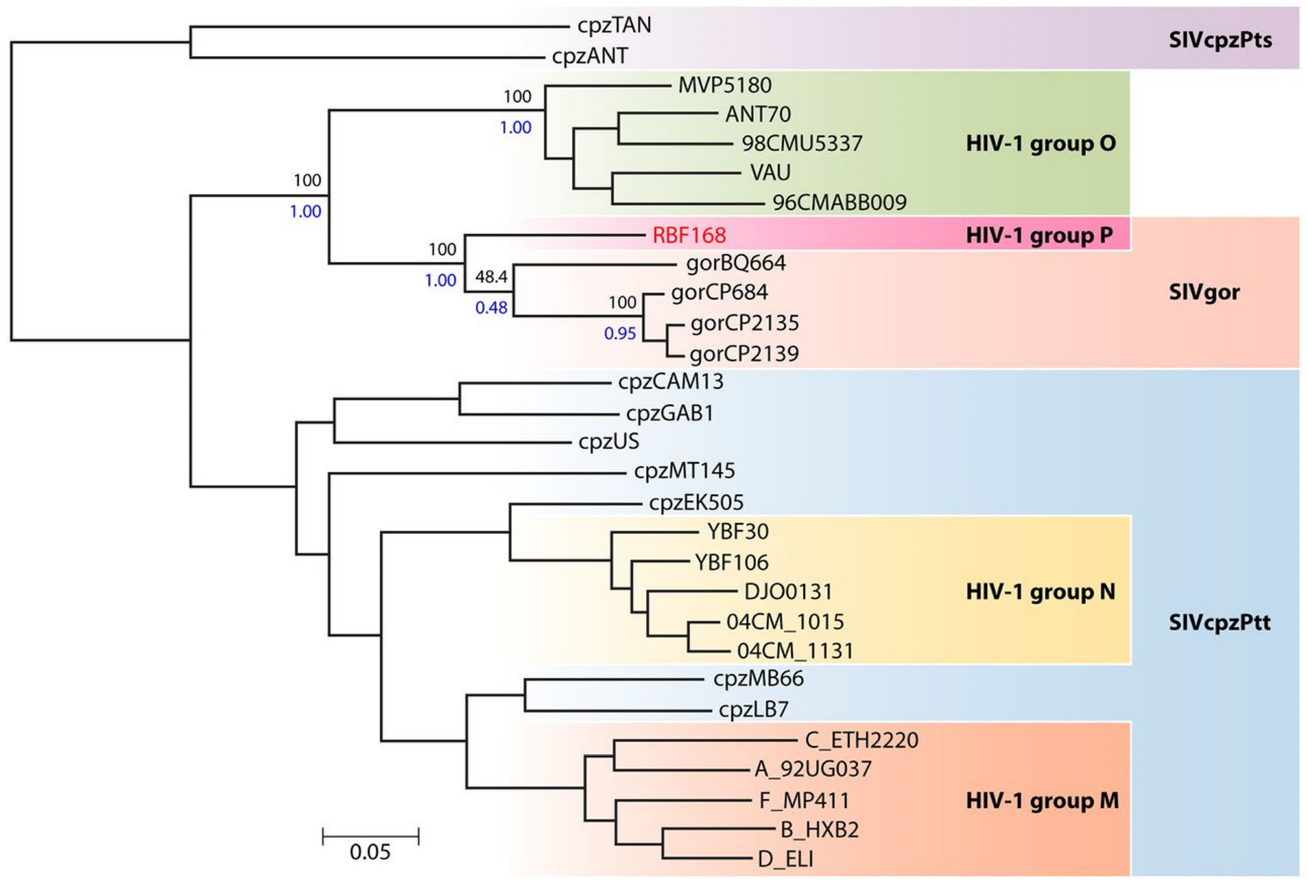
:1. HIV-1 Non-M Variants
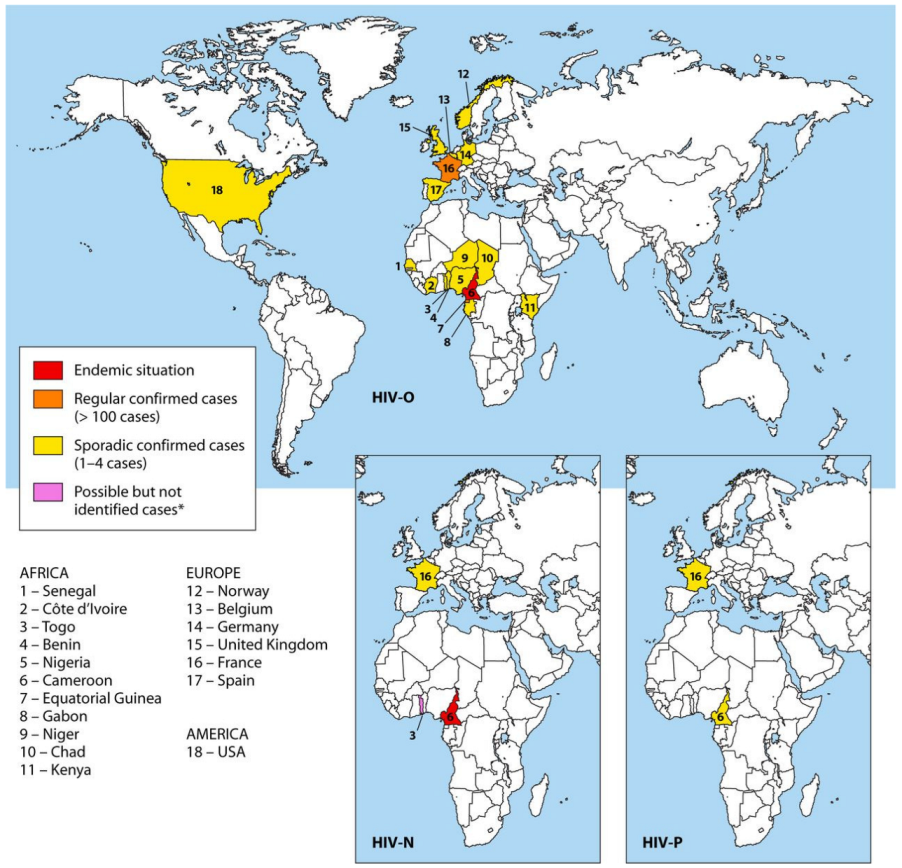
2. Epidemiology
3. Diagnosis and Virological Monitoring
4. Antiretroviral Agents
4.1. Natural Resistance (In Vitro Data)
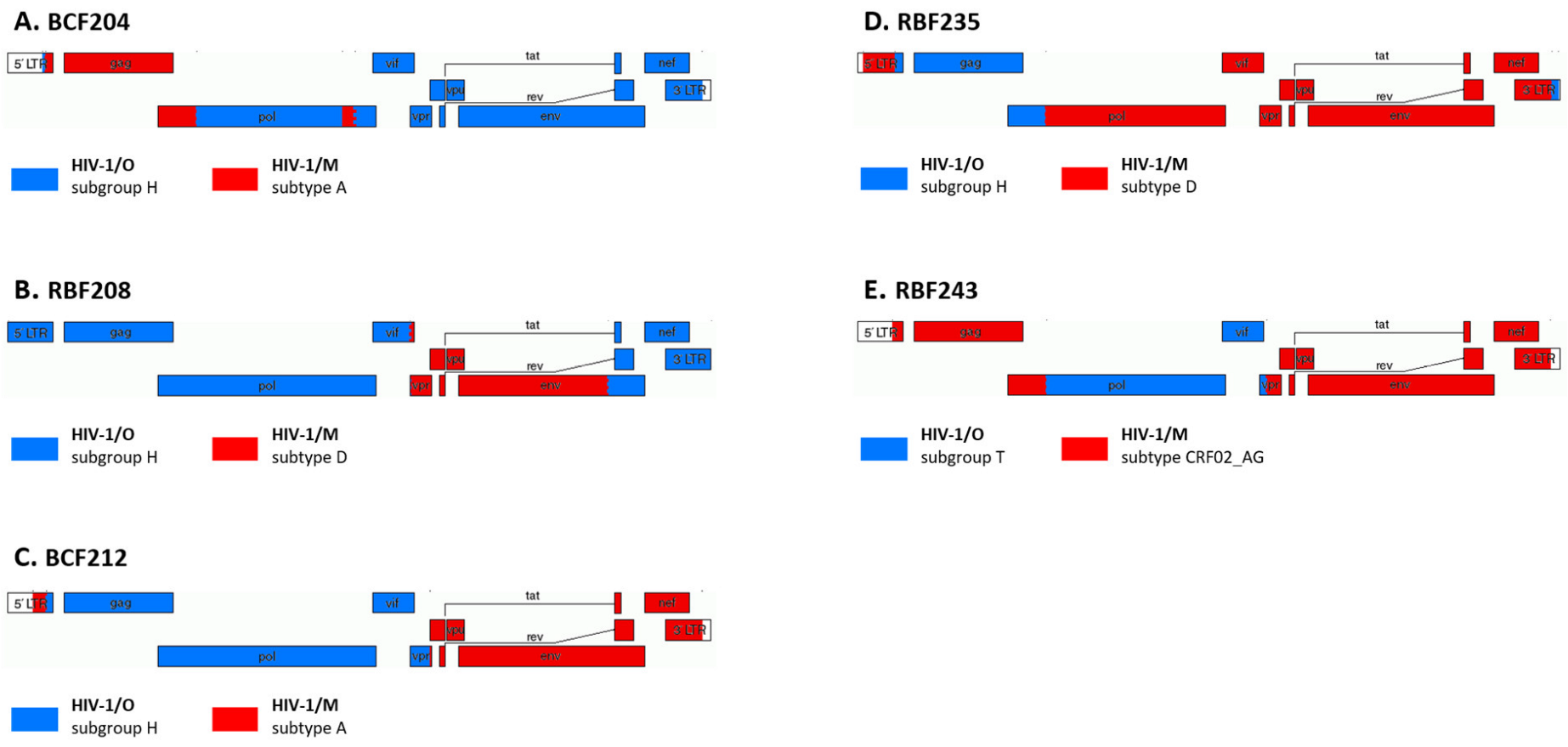
4.2. Antiretroviral Therapy and Virological Failure (In Vivo Data)
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barré-Sinoussi, F.; Chermann, J.C.; Rey, F.; Nugeyre, M.T.; Chamaret, S.; Gruest, J.; Dauguet, C.; Axler-Blin, C.; Vézinet-Brun, F.; Rouzioux, C.; et al. T-lymphotropic retrovirus from a patient at risk for acquired immune deficiency syndrome (AIDS). Science 1983, 220, 868–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barin, F.; Denis, F.; Allan, J.; M’Boup, S.; Kanki, P.; Lee, T.; Essex, M. Serological evidence for virus related to simian T-lymphotropic retrovirus III in residents of west Africa. Lancet 1985, 326, 1387–1389. [Google Scholar] [CrossRef] [PubMed]
- Clavel, F.; Guétard, D.; Brun-Vézinet, F.; Chamaret, S.; Rey, M.-A.; Santos-Ferreira, M.O.; Laurent, A.G.; Dauguet, C.; Katlama, C.; Rouzioux, C.; et al. Isolation of a new human retrovirus from West African patients with AIDS. Science 1986, 233, 343–346. [Google Scholar] [CrossRef] [PubMed]
- De Leys, R.; Vanderborght, B.; Vanden Haesevelde, M.; Heyndrickx, L.; van Geel, A.; Wauters, C.; Bernaerts, R.; Saman, E.; Nijs, P.; Willems, B. Isolation and partial characterization of an unusual human immunodeficiency retrovirus from two persons of west-central African origin. J. Virol. 1990, 64, 1207–1216. [Google Scholar] [CrossRef] [Green Version]
- Simon, F.; Mauclère, P.; Roques, P.; Loussert-Ajaka, I.; Müller-Trutwin, M.; Saragosti, S.; Georges-Courbot, M.C.; Barré-Sinoussi, F.; Brun-Vézinet, F. Identification of a new human immunodeficiency virus type 1 distinct from group M and group O. Nat. Med. 1998, 4, 1032–1037. [Google Scholar] [CrossRef]
- Plantier, J.-C.; Leoz, M.; Dickerson, J.E.; De Oliveira, F.; Cordonnier, F.; Lemée, V.; Damond, F.; Robertson, D.L.; Simon, F. A new human immunodeficiency virus derived from gorillas. Nat. Med. 2009, 15, 871–872. [Google Scholar] [CrossRef]
- Keele, B.F.; Van Heuverswyn, F.; Li, Y.; Bailes, E.; Takehisa, J.; Santiago, M.L.; Bibollet-Ruche, F.; Chen, Y.; Wain, L.V.; Liegeois, F.; et al. Chimpanzee reservoirs of pandemic and nonpandemic HIV-1. Science 2006, 313, 523–526. [Google Scholar] [CrossRef] [Green Version]
- Van Heuverswyn, F.; Li, Y.; Bailes, E.; Neel, C.; Lafay, B.; Keele, B.F.; Shaw, K.S.; Takehisa, J.; Kraus, M.H.; Loul, S.; et al. Genetic diversity and phylogeographic clustering of SIVcpzPtt in wild chimpanzees in Cameroon. Virology 2007, 368, 155–171. [Google Scholar] [CrossRef]
- D’Arc, M.; Ayouba, A.; Esteban, A.; Learn, G.H.; Boué, V.; Liegeois, F.; Etienne, L.; Tagg, N.; Leendertz, F.H.; Boesch, C.; et al. Origin of the HIV-1 group O epidemic in western lowland gorillas. Proc. Natl. Acad. Sci. USA 2015, 112, E1343–E1352. [Google Scholar] [CrossRef] [Green Version]
- Preston, B.D.; Poiesz, B.J.; Loeb, L.A. Fidelity of HIV-1 reverse transcriptase. Science 1988, 242, 1168–1171. [Google Scholar] [CrossRef]
- Mansky, L.M.; Temin, H.M. Lower in vivo mutation rate of human immunodeficiency virus type 1 than that predicted from the fidelity of purified reverse transcriptase. J. Virol. 1995, 69, 5087–5094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, D.D.; Neumann, A.U.; Perelson, A.S.; Chen, W.; Leonard, J.M.; Markowitz, M. Rapid turnover of plasma virions and CD4 lymphocytes in HIV-1 infection. Nature 1995, 373, 123–126. [Google Scholar] [CrossRef]
- Smyth, R.P.; Davenport, M.P.; Mak, J. The origin of genetic diversity in HIV-1. Virus Res. 2012, 169, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Vijay, N.N.V.; Vasantika; Ajmani, R.; Perelson, A.S.; Dixit, N.M. Recombination increases human immunodeficiency virus fitness, but not necessarily diversity. J. Gen. Virol. 2008, 89, 1467–1477. [Google Scholar] [CrossRef] [PubMed]
- Streeck, H.; Li, B.; Poon, A.; Schneidewind, A.; Gladden, A.D.; Power, K.A.; Daskalakis, D.; Bazner, S.; Zuniga, R.; Brander, C.; et al. Immune-driven recombination and loss of control after HIV superinfection. J. Exp. Med. 2008, 205, 1789–1796. [Google Scholar] [CrossRef] [Green Version]
- Nora, T.; Charpentier, C.; Tenaillon, O.; Hoede, C.; Clavel, F.; Hance, A.J. Contribution of recombination to the evolution of human immunodeficiency viruses expressing resistance to antiretroviral treatment. J. Virol. 2007, 81, 7620–7628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon-Loriere, E.; Holmes, E.C. Why do RNA viruses recombine? Nat. Rev. Microbiol. 2011, 9, 617–626. [Google Scholar] [CrossRef]
- Rambaut, A.; Posada, D.; Crandall, K.A.; Holmes, E. The causes and consequences of HIV evolution. Nat. Rev. Genet. 2004, 5, 52–61. [Google Scholar] [CrossRef]
- Hemelaar, J.; Gouws, E.; Ghys, P.D.; Osmanov, S. Global and regional distribution of HIV-1 genetic subtypes and recombinants in 2004. Aids 2006, 20, W13–W23. [Google Scholar] [CrossRef]
- Peeters, M.; Liegeois, F.; Torimiro, N.; Bourgeois, A.; Mpoudi, E.; Vergne, L.; Saman, E.; Delaporte, E.; Saragosti, S. Characterization of a highly replicative intergroup M/O human immunodeficiency virus type 1 recombinant isolated from a Cameroonian patient. J. Virol. 1999, 73, 7368–7375. [Google Scholar] [CrossRef] [Green Version]
- Takehisa, J.; Zekeng, L.; Ido, E.; Yamaguchi-Kabata, Y.; Mboudjeka, I.; Harada, Y.; Miura, T.; Kaptué, L.; Hayami, M. Human immunodeficiency virus type 1 intergroup (M/O) recombination in cameroon. J. Virol. 1999, 73, 6810–6820. [Google Scholar] [CrossRef] [Green Version]
- Mourez, T.; Simon, F.; Plantier, J.-C. Non-M Variants of Human Immunodeficiency Virus Type 1. Clin. Microbiol. Rev. 2013, 26, 448–461. [Google Scholar] [CrossRef] [Green Version]
- Delaugerre, C.; De Oliveira, F.; Lascoux-Combe, C.; Plantier, J.-C.; Simon, F. HIV-1 group N: Travelling beyond Cameroon. Lancet 2011, 378, 1894. [Google Scholar] [CrossRef]
- Delaporte, E.; Janssens, W.; Peeters, M.; Buvé, A.; Dibanga, G.; Perret, J.-L.; Ditsambou, V.; Mba, J.-R.; Courbot, M.-C.G.; Georges, A.; et al. Epidemiological and molecular characteristics of HIV infection in Gabon, 1986–1994. Aids 1996, 10, 903–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zekeng, L.; Sima, J.O.; Hampl, H.; Ndemesogo, J.M.; Ntutumu, J.; Sima, V.; Devare, S.; Kaptue, L.; Gürtler, L. Update on HIV-1 group O infection in Equatorial Guinea, Central Africa. Aids 1997, 11, 1410–1412. [Google Scholar] [PubMed]
- Hunt, J.C.; Brennan, C.A.; Golden, A.M.; Yamaguchi, J.; Lund, J.K.; Vallari, A.S.; Hickman, R.K.; Zekeng, L.; Gürtler, L.G.; Hampl, H.; et al. Molecular analyses of HIV-1 group O and HIV-2 variants from Africa. Leukemia 1997, 11 (Suppl. S3), 138–141. [Google Scholar] [PubMed]
- Bibollet-Ruche, F.; Peeters, M.; Mboup, S.; Ekaza, E.; Gandji, R.; Torimiro, J.; Mpoudi, E.N.; Amblard, J.; Dibanga, G.; Saidou, M.; et al. Molecular characterization of the envelope transmembrane glycoprotein of 13 new human immunodeficiency virus type 1 group O strains from six different African countries. AIDS Res. Hum. Retrovir. 1998, 14, 1281–1285. [Google Scholar] [CrossRef] [Green Version]
- Vessière, A.; Rousset, D.; Kfutwah, A.; Leoz, M.; Depatureaux, A.; Simon, F.; Plantier, J.-C. Diagnosis and monitoring of HIV-1 group O-infected patients in Cameroun. Am. J. Ther. 2010, 53, 107–110. [Google Scholar] [CrossRef]
- De Oliveira, F.; Mourez, T.; Vessiere, A.; Ngoupo, P.A.; Alessandri-Gradt, E.; Simon, F.; Rousset, D.; Plantier, J.-C. Multiple HIV-1/M + HIV-1/O dual infections and new HIV-1/MO inter-group recombinant forms detected in Cameroon. Retrovirology 2017, 14, 1. [Google Scholar] [CrossRef] [Green Version]
- Vergne, L.; Bourgeois, A.; Mpoudi-Ngole, E.; Mougnutou, R.; Mbuagbaw, J.; Liegeois, F.; Laurent, C.; Butel, C.; Zekeng, L.; Delaporte, E.; et al. Biological and genetic characteristics of HIV infections in Cameroon reveals dual group M and O infections and a correlation between SI-inducing phenotype of the predominant CRF02_AG variant and disease stage. Virology 2003, 310, 254–266. [Google Scholar] [CrossRef] [Green Version]
- Ngoupo, P.A.; Sadeuh-Mba, S.A.; De Oliveira, F.; Ngono, V.; Ngono, L.; Tchendjou, P.; Penlap, V.; Mourez, T.; Njouom, R.; Kfutwah, A.; et al. First evidence of transmission of an HIV-1 M/O intergroup recombinant virus. Aids 2016, 30, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, J.; Bodelle, P.; Vallari, A.S.; Coffey, R.; McArthur, C.P.; Schochetman, G.; Devare, S.G.; Brennan, C.A. HIV infections in northwestern Cameroon: Identification of HIV type 1 group O and dual HIV type 1 group M and group O infections. AIDS Res. Hum. Retrovir. 2004, 20, 944–957. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, J.; Coffey, R.; Vallari, A.; Ngansop, C.; Mbanya, D.; Ndembi, N.; Kaptué, L.; Gürtler, L.G.; Bodelle, P.; Schochetman, G.; et al. Identification of HIV type 1 group N infections in a husband and wife in Cameroon: Viral genome sequences provide evidence for horizontal transmission. AIDS Res. Hum. Retrovir. 2006, 22, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, J.; McArthur, C.P.; Vallari, A.; Coffey, R.; Bodelle, P.; Beyeme, M.; Schochetman, G.; Devare, S.G.; Brennan, C.A. HIV-1 Group N: Evidence of ongoing transmission in Cameroon. AIDS Res. Hum. Retrovir. 2006, 22, 453–457. [Google Scholar] [CrossRef]
- Vallari, A.; Bodelle, P.; Ngansop, C.; Makamche, F.; Ndembi, N.; Mbanya, D.; Kaptué, L.; Gürtler, L.G.; McArthur, C.P.; Devare, S.G.; et al. Four New HIV-1 Group N Isolates from Cameroon: Prevalence Continues to Be Low. AIDS Res. Hum. Retrovir. 2010, 26, 109–115. [Google Scholar] [CrossRef]
- Ayouba, A.; Souquières, S.; Njinku, B.; Martin, P.M.V.; Müller-Trutwin, M.C.; Roques, P.; Barré-Sinoussi, F.; Mauclère, P.; Simon, F.; Nerrienet, E. HIV-1 group N among HIV-1-seropositive individuals in Cameroon. Aids 2000, 14, 2623–2625. [Google Scholar] [CrossRef]
- Rodgers, M.; Vallari, A.; Harris, B.; Yamaguchi, J.; Holzmayer, V.; Forberg, K.; Berg, M.; Kenmenge, J.; Ngansop, C.; Awazi, B.; et al. Identification of rare HIV-1 Group N, HBV AE, and HTLV-3 strains in rural South Cameroon. Virology 2017, 504, 141–151. [Google Scholar] [CrossRef]
- Rodgers, M.A.; Vallari, A.S.; Yamaguchi, J.; Holzmayer, V.; Harris, B.; Toure-Kane, C.; Mboup, S.; Badreddine, S.; McArthur, C.; Ndembi, N.; et al. ARCHITECT HIV Combo Ag/Ab and RealTime HIV-1 Assays Detect Diverse HIV Strains in Clinical Specimens. AIDS Res. Hum. Retrovir. 2018, 34, 314–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roques, P.; Robertson, D.L.; Souquière, S.; Apetrei, C.; Nerrienet, E.; Barré-Sinoussi, F.; Müller-Trutwin, M.; Simon, F. Phylogenetic characteristics of three new HIV-1 N strains and implications for the origin of group N. Aids 2004, 18, 1371–1381. [Google Scholar] [CrossRef]
- Tagnouokam Ngoupo, P.A.; Sadeuh-Mba, S.A.; De Oliveira, F.; Ngo-Malabo, E.T.; Ngono, L.; Plantier, J.C.; Kfutwah, A.; Njouom, R. Short Communication: Characterization of a New HIV-1 Group N Isolate Originating from a Cameroonian Patient. AIDS Res. Hum. Retrovir. 2018, 34, 621–625. [Google Scholar] [CrossRef]
- Alessandri-Gradt, E.; De Oliveira, F.; Leoz, M.; Lemee, V.; Robertson, D.L.; Feyertag, F.; Ngoupo, P.-A.; Mauclere, P.; Simon, F.; Plantier, J.-C. HIV-1 group P infection: Towards a dead-end infection? Aids 2018, 32, 1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallari, A.; Holzmayer, V.; Harris, B.; Yamaguchi, J.; Ngansop, C.; Makamche, F.; Mbanya, D.; Kaptué, L.; Ndembi, N.; Gürtler, L.; et al. Confirmation of Putative HIV-1 Group P in Cameroon. J. Virol. 2011, 85, 1403–1407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabeya, C.M.; Esu-Williams, E.; Eni, E.; Peeters, M.; Saman, E.; Delaporte, E. Evidence for HIV-1 group O infection in Nigeria. Lancet 1995, 346, 308. [Google Scholar] [CrossRef] [PubMed]
- Songok, E.; Libondo, D.; Rotich, M.; Oogo, S.; Tukei, P. Surveillance for HIV-1 subtypes O and M in Kenya. Lancet 1996, 347, 1700. [Google Scholar] [CrossRef] [PubMed]
- Heyndrickx, L.; Alary, M.; Janssens, W.; Davo, N.; van der Groen, G. HIV-1 group O and group M dual infection in Bénin. Lancet 1996, 347, 902–903. [Google Scholar] [CrossRef] [PubMed]
- Peeters, M.; Gaye, A.; Mboup, S.; Badombena, W.; Bassabi, K.; Prince-David, M.; Develoux, M.; Liegeois, F.; van der Groen, G.; Saman, E.; et al. Presence of HIV-1 group O infection in West Africa. Aids 1996, 10, 343–344. [Google Scholar] [CrossRef]
- Peeters, M.; Gueye, A.; Mboup, S.; Bibollet-Ruche, F.; Ekaza, E.; Mulanga, C.; Ouedrago, R.; Gandji, R.; Mpele, P.; Dibanga, G.; et al. Geographical distribution of HIV-1 group O viruses in Africa. Aids 1997, 11, 493–498. [Google Scholar] [CrossRef]
- Nkengasong, J.; Sylla-Koko, F.; Peeters, M.; Ellenberger, D.; Sassan-Morokro, M.; Ekpini, R.-A.; Msellati, P.; Greenberg, A.E.; Combe, P.; Rayfield, M. HIV-1 group O virus infection in Abidjan, Côte d’lvoire. Aids 1998, 12, 1565–1566. [Google Scholar] [CrossRef]
- Hampl, H.; Sawitzky, D.; Stöffler-Meilicke, M.; Groh, A.; Schmitt, M.; Eberle, J.; Gürtler, L. First case of HIV-1 subtype 0 infection in Germany. Infection 1995, 23, 369–370. [Google Scholar] [CrossRef]
- Rayfield, M.A.; Sullivan, P.; Bandea, C.I.; Britvan, L.; Otten, R.A.; Pau, C.P.; Pieniazek, D.; Subbarao, S.; Simon, P.; Schable, C.A.; et al. HIV-1 group O virus identified for the first time in the United States. Emerg. Infect. Dis. 1996, 2, 209–212. [Google Scholar] [CrossRef] [Green Version]
- Soriano, V.; Gutierrez, M.; García-Lerma, G.; Aguilera, O.; Mas, A.; Bravo, R.; Pérez-Labad, M.L.; Baquero, M.; González-Lahoz, J. First case of HIV-1 group O infection in Spain. Vox Sang. 1996, 71, 66. [Google Scholar] [CrossRef]
- Sullivan, P.S.; Do, A.N.; Ellenberger, D.; Pau, C.-P.; Paul, S.; Robbins, K.; Kalish, M.; Storck, C.; Schable, C.A.; Wise, H.; et al. Human immunodeficiency virus (HIV) subtype surveillance of African-born persons at risk for group O and group N HIV infections in the United States. J. Infect. Dis. 2000, 181, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Gould, K. Infection with HIV-1 group O. AIDS Patient Care STDs 1997, 11, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Depatureaux, A.; Leoz, M.; De Oliveira, F.; Gueudin, M.; Damond, F.; Descamps, D.; Brun-Vézinet, F.; Lemée, V.; Simon, F.; Barin, F.; et al. Diagnostic spécifique et prise en charge des infections par un VIH-1 groupe O: Données de RES-O. Médecine Mal. Infect. 2010, 40, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Agut, H.; Rabanel, B.; Candotti, D.; Huraux, J.-M.; Remy, G.; Tabary, T.; Ingrand, D.; Chippaux, C.; Chamaret, S.; Dauguet, C.; et al. Isolation of atypical HIV-1-related retrovirus from AIDS patient. Lancet 1992, 340, 681–682. [Google Scholar] [CrossRef]
- Plantier, J.-C.; Lemée, V.; Dorval, I.; Gueudin, M.; Braun, J.; Hutin, P.; Ruffault, A.; Simon, F. HIV-1 group M superinfection in an HIV-1 group O-infected patient. Aids 2004, 18, 2444–2446. [Google Scholar]
- Brand, D.; Beby-Defaux, A.; Macé, M.; Brunet, S.; Moreau, A.; Godet, C.; Jais, X.; Cazein, F.; Semaille, C.; Barin, F. First identification of HIV-1 groups M and O dual infections in Europe. Aids 2004, 18, 2425–2428. [Google Scholar]
- De Oliveira, F.; Cappy, P.; Lemée, V.; Moisan, A.; Pronier, C.; Bocket, L.; Bouvier-Alias, M.; Chaix, M.-L.; Gault, E.; Morvan, O.; et al. Detection of numerous HIV-1/MO recombinants in France. Aids 2018, 32, 1289–1299. [Google Scholar] [CrossRef]
- Vessière, A.; Leoz, M.; Brodard, V.; Strady, C.; Lemée, V.; Depatureaux, A.; Simon, F.; Plantier, J.-C. First evidence of a HIV-1 M/O recombinant form circulating outside Cameroon. Aids 2010, 24, 1079–1082. [Google Scholar] [CrossRef]
- Moisan, A.; De Oliveira, F.; Pronier, C.; Cappy, P.; Maillard, A.; Plantier, J.C. In vivo emergence of an HIV-1/MO recombinant revealed undiagnosed HIV-1/M+O co-infection. Clin. Microbiol. Infect. 2020, 26, 262–264. [Google Scholar] [CrossRef]
- Moisan, A.; De Oliveira, F.; Cappy, P.; Ngoupo, P.A.; Njouom, R.; Plantier, J.C. Evolution of HIV-1 groups M and O: Genetic comparative analysis of 23 HIV-1/MO inter-group recombinant forms. In Proceedings of the 9th IAS Conference on HIV Science, Paris, France, 23 July 2017. [Google Scholar]
- Gautheret-Dejean, A.; Mesmin-Poho, S.; Birguel, J.; Lemée, V.; Huraux, J.-M.; Plantier, J.-C. Unequal detection of HIV type 1 group O infection by simple rapid tests. Clin. Infect. Dis. 2008, 46, 1936–1937. [Google Scholar] [CrossRef] [Green Version]
- Aghokeng, A.F.; Mpoudi-Ngole, E.; Dimodi, H.; Atem-Tambe, A.; Tongo, M.; Butel, C.; Delaporte, E.; Peeters, M. Inaccurate diagnosis of HIV-1 group M and O is a key challenge for ongoing universal access to antiretroviral treatment and HIV prevention in Cameroon. PLoS ONE 2009, 4, e7702. [Google Scholar] [CrossRef] [Green Version]
- Mourez, T.; Lemée, V.; Delbos, V.; Delaugerre, C.; Alessandri-Gradt, E.; Etienne, M.; Simon, F.; Chaix, M.-L.; Plantier, J.-C. HIV rapid screening tests and self-tests: Be aware of differences in performance and cautious of vendors. Ebiomedicine 2018, 37, 382–391. [Google Scholar] [CrossRef] [Green Version]
- Zouhair, S.; Roussin-Bretagne, S.; Moreau, A.; Brunet, S.; Laperche, S.; Maniez, M.; Barin, F.; Harzic, M. Group O Human Immunodeficiency Virus Type 1 Infection That Escaped Detection in Two Immmunoassays. J. Clin. Microbiol. 2006, 44, 662–665. [Google Scholar] [CrossRef] [Green Version]
- Plantier, J.-C.; Djemai, M.; Lemée, V.; Reggiani, A.; Leoz, M.; Burc, L.; Vessière, A.; Rousset, D.; Poveda, J.-D.; Henquell, C.; et al. Census and Analysis of Persistent False-Negative Results in Serological Diagnosis of Human Immunodeficiency Virus Type 1 Group O Infections. J. Clin. Microbiol. 2009, 47, 2906–2911. [Google Scholar] [CrossRef] [Green Version]
- Swanson, P.; de Mendoza, C.; Joshi, Y.; Golden, A.; Hodinka, R.L.; Soriano, V.; Devare, S.G.; Hackett, J. Impact of human immunodeficiency virus type 1 (HIV-1) genetic diversity on performance of four commercial viral load assays: LCx HIV RNA Quantitative, AMPLICOR HIV-1 MONITOR v1.5, VERSANT HIV-1 RNA 3.0, and NucliSens HIV-1 QT. J. Clin. Microbiol. 2005, 43, 3860–3868. [Google Scholar] [CrossRef] [Green Version]
- Plantier, J.-C.; Gueudin, M.; Damond, F.; Braun, J.; Mauclère, P.; Simon, F. Plasma RNA quantification and HIV-1 divergent strains. Am. J. Ther. 2003, 33, 1–7. [Google Scholar] [CrossRef]
- Gueudin, M.; Plantier, J.C.; Lemée, V.; Schmitt, M.P.; Chartier, L.; Bourlet, T.; Ruffault, A.; Damond, F.; Vray, M.; Simon, F. Evaluation of the Roche Cobas TaqMan and Abbott RealTime extraction-quantification systems for HIV-1 subtypes. Am. J. Ther. 2007, 44, 500–505. [Google Scholar] [CrossRef]
- Sire, J.-M.; Vray, M.; Merzouk, M.; Plantier, J.-C.; Pavie, J.; Maylin, S.; Timsit, J.; Lascoux-Combe, C.; Molina, J.-M.; Simon, F.; et al. Comparative RNA quantification of HIV-1 group M and non-M with the Roche Cobas AmpliPrep/Cobas TaqMan HIV-1 v2.0 and Abbott Real-Time HIV-1 PCR assays. Am. J. Ther. 2011, 56, 239–243. [Google Scholar] [CrossRef]
- Tang, N.; Huang, S.; Salituro, J.; Mak, W.B.; Cloherty, G.; Johanson, J.; Li, Y.H.; Schneider, G.; Robinson, J.; Hackett, J., Jr.; et al. A RealTime HIV-1 viral load assay for automated quantitation of HIV-1 RNA in genetically diverse group M subtypes A-H, group O and group N samples. J. Virol. Methods 2007, 146, 236–245. [Google Scholar] [CrossRef]
- Alessandri-Gradt, E.; Unal, G.; Baron, A.; Leoz, M.; Gueudin, M.; Plantier, J.-C.; Network, T.R.-O. Performance Analysis of Three Commercial Kits Designed for RNA Quantification of HIV-1 Group O Variants. Am. J. Ther. 2021, 88, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Berger, A.; Muenchhoff, M.; Hourfar, K.; Kortenbusch, M.; Ambiel, I.; Stegmann, L.; Heim, A.; Sarrazin, C.; Ehret, R.; Daniel, V.; et al. Severe underquantification of HIV-1 group O isolates by major commercial PCR-based assays. Clin. Microbiol. Infect. 2020, 26, 1688.e1–1688.e7. [Google Scholar] [CrossRef] [PubMed]
- Gueudin, M.; Baron, A.; Alessandri-Gradt, E.; Lemée, V.; Mourez, T.; Etienne, M.; Plantier, J.-C. Performance Evaluation of the New HIV-1 Quantification Assay, Xpert HIV-1 Viral Load, on a Wide Panel of HIV-1 Variants. Am. J. Ther. 2016, 72, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Mourez, T.; Delaugerre, C.; Vray, M.; Lemée, V.; Simon, F.; Plantier, J.C. Comparison of the bioMérieux NucliSENS EasyQ HIV-1 v2.0-HIV-1 RNA quantification assay versus Abbott RealTime HIV-1 and Roche Cobas TaqMan HIV-1 v2.0 on current epidemic HIV-1 variants. J. Clin. Virol. 2015, 71, 76–81. [Google Scholar] [CrossRef]
- Depatureaux, A.; Charpentier, C.; Leoz, M.; Unal, G.; Damond, F.; Kfutwah, A.; Vessière, A.; Simon, F.; Plantier, J.-C. Impact of HIV-1 group O genetic diversity on genotypic resistance interpretation by algorithms designed for HIV-1 group M. Am. J. Ther. 2011, 56, 139–145. [Google Scholar] [CrossRef]
- Péré, H.; Roques, P.; Talla, F.; Meillo, H.; Charpentier, C.; Bélec, L. Sustained virological failure in Cameroonese patient infected by HIV-1 group N evidenced by sequence-based genotyping assay. Aids 2015, 29, 1267–1269. [Google Scholar] [CrossRef]
- Descamps, D.; Collin, G.; Letourneur, F.; Apetrei, C.; Damond, F.; Loussert-Ajaka, I.; Simon, F.; Saragosti, S.; Brun-Vézinet, F. Susceptibility of human immunodeficiency virus type 1 group O isolates to antiretroviral agents: In vitro phenotypic and genotypic analyses. J. Virol. 1997, 71, 8893–8898. [Google Scholar] [CrossRef] [Green Version]
- Tebit, D.M.; Patel, H.; Ratcliff, A.; Alessandri, E.; Liu, J.; Carpenter, C.; Plantier, J.-C.; Arts, E.J. HIV-1 Group O Genotypes and Phenotypes: Relationship to Fitness and Susceptibility to Antiretroviral Drugs. AIDS Res. Hum. Retrovir. 2016, 32, 676–688. [Google Scholar] [CrossRef] [Green Version]
- Parkin, N.T.; Schapiro, J.M. Antiretroviral drug resistance in non-subtype B HIV-1, HIV-2 and SIV. Antivir. Ther. 2004, 9, 3–12. [Google Scholar] [CrossRef]
- Tuaillon, E.; Gueudin, M.; Lemée, V.; Gueit, I.; Roques, P.; Corrigan, G.E.; Plantier, J.-C.; Simon, F.; Braun, J. Phenotypic Susceptibility to Nonnucleoside Inhibitors of Virion-Associated Reverse Transcriptase From Different HIV Types and Groups. Am. J. Ther. 2004, 37, 1543–1549. [Google Scholar] [CrossRef]
- Villabona-Arenas, C.J.; Domyeum, J.; Mouacha, F.; Butel, C.; Delaporte, E.; Peeters, M.; Mpoudi-Ngole, E.; Aghokeng, A.F. HIV-1 group O infection in Cameroon from 2006 to 2013: Prevalence, genetic diversity, evolution and public health challenges. Infect. Genet. Evol. 2015, 36, 210–216. [Google Scholar] [CrossRef] [Green Version]
- Tebit, D.M.; Lobritz, M.; Lalonde, M.; Immonen, T.; Singh, K.; Sarafianos, S.; Herchenröder, O.; Kräusslich, H.-G.; Arts, E.J. Divergent Evolution in Reverse Transcriptase (RT) of HIV-1 Group O and M Lineages: Impact on Structure, Fitness, and Sensitivity to Nonnucleoside RT Inhibitors. J. Virol. 2010, 84, 9817–9830. [Google Scholar] [CrossRef] [Green Version]
- Leoz, M.; Feyertag, F.; Kfutwah, A.; Mauclère, P.; Lachenal, G.; Damond, F.; De Oliveira, F.; Lemée, V.; Simon, F.; Robertson, D.L.; et al. The Two-Phase Emergence of Non Pandemic HIV-1 Group O in Cameroon. PLOS Pathog. 2015, 11, e1005029. [Google Scholar] [CrossRef] [PubMed]
- de Baar, M.P.; Janssens, W.; de Ronde, A.; Fransen, K.; Colebunders, R.; Kestens, L.; van der Groen, G.; Goudsmit, J. Natural residues versus antiretroviral drug-selected mutations in HIV type 1 group O reverse transcriptase and protease related to virological drug failure in vivo. AIDS Res. Hum. Retrovir. 2000, 16, 1385–1394. [Google Scholar] [CrossRef] [PubMed]
- Luk, K.-C.; Kaptue, L.; Zekeng, L.; Soriano, V.; Gürtler, L.; Devare, S.G.; Schochetman, G.; Hackett, J. Naturally occurring sequence polymorphisms within HIV type 1 group O protease. AIDS Res. Hum. Retrovir. 2001, 17, 1555–1561. [Google Scholar] [CrossRef]
- Depatureaux, A.; Charpentier, C.; Collin, G.; Leoz, M.; Descamps, D.; Vessière, A.; Damond, F.; Rousset, D.; Brun-Vézinet, F.; Plantier, J.-C. Baseline Genotypic and Phenotypic Susceptibilities of HIV-1 Group O to Enfuvirtide. Antimicrob. Agents Chemother. 2010, 54, 4016–4019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roques, P.; Robertson, D.; Souquière, S.; Damond, F.; Ayouba, A.; Farfara, I.; Depienne, C.; Nerrienet, E.; Dormont, D.; Vézinetd, F.; et al. Phylogenetic Analysis of 49 Newly Derived HIV-1 Group O Strains: High Viral Diversity but No Group M-like Subtype Structure. Virology 2002, 302, 259–273. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, J.; Vallari, A.S.; Swanson, P.; Bodelle, P.; Kaptué, L.; Ngansop, C.; Zekeng, L.; Gürtler, L.G.; Devare, S.G.; Brennan, C.A. Evaluation of HIV type 1 group O isolates: Identification of five phylogenetic clusters. AIDS Res. Hum. Retrovir. 2002, 18, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Alessandri-Gradt, E.; Charpentier, C.; Leoz, M.; Mourez, T.; Descamps, D.; Plantier, J.-C. Impact of natural polymorphisms of HIV-1 non-group M on genotypic susceptibility to the attachment inhibitor fostemsavir. J. Antimicrob. Chemother. 2018, 73, 2716–2720. [Google Scholar] [CrossRef] [Green Version]
- Leoz, M.; Depatureaux, A.; Vessière, A.; Roquebert, B.; Damond, F.; Rousset, D.; Roques, P.; Simon, F.; Plantier, J.-C. Integrase polymorphism and HIV-1 group O diversity. Aids 2008, 22, 1239–1243. [Google Scholar] [CrossRef]
- Briz, V.; Garrido, C.; Poveda, E.; Morello, J.; Barreiro, P.; de Mendoza, C.; Soriano, V. Raltegravir and etravirine are active against HIV type 1 group O. AIDS Res. Hum. Retrovir. 2009, 25, 225–227. [Google Scholar] [CrossRef] [PubMed]
- Alessandri-Gradt, E.; Collin, G.; Tourneroche, A.; Bertine, M.; Leoz, M.; Charpentier, C.; Unal, G.; Descamps, D.; Plantier, J.C. HIV-1 non-group M phenotypic susceptibility to integrase strand transfer inhibitors. J. Antimicrob. Chemother. 2017, 72, 2431–2437. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.; Gracias, S.; Charpentier, C.; Descamps, D.; Le Hingrat, Q.; Plantier, J.-C.; Alessandri-Gradt, E. HIV-1 non-group M phenotypic susceptibility in vitro to bictegravir and cabotegravir. J. Antimicrob. Chemother. 2021, 76, 2306–2309. [Google Scholar] [CrossRef] [PubMed]
- Tsiang, M.; Jones, G.S.; Goldsmith, J.; Mulato, A.; Hansen, D.; Kan, E.; Tsai, L.; Bam, R.A.; Stepan, G.; Stray, K.M.; et al. Antiviral Activity of Bictegravir (GS-9883), a Novel Potent HIV-1 Integrase Strand Transfer Inhibitor with an Improved Resistance Profile. Antimicrob. Agents Chemother. 2016, 60, 7086–7097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Depatureaux, A.M.; Leoz, M.M.; Le Moal, G.; Pathé, J.-P.; Pavie, J.; Batisse, D.; Daneluzzi, V.; Genet, P.M.; Gerard, L.; Lascaux-Cametz, A.-S.; et al. Raltegravir-Based Regimens Are Effective in HIV-1 Group O–Infected Patients. Am. J. Ther. 2012, 61, e1–e3. [Google Scholar] [CrossRef]
- Rodes, B.; De Mendoza, C.; Rodgers, M.; Newell, A.; Jimenez, V.; Lopez-Brugada, R.M.; Soriano, V. Treatment response and drug resistance in patients infected with HIV type 1 group O viruses. AIDS Res. Hum. Retrovir. 2005, 21, 602–607. [Google Scholar] [CrossRef]
- Aghokeng, A.F.; Kouanfack, C.; Peeters, M.; Mpoudi-Ngole, E.; Delaporte, E. Successful integrase inhibitor-based highly active antiretroviral therapy for a multidrug-class-resistant HIV type 1 group O-infected patient in Cameroon. AIDS Res. Hum. Retrovir. 2013, 29, 1–3. [Google Scholar] [CrossRef]
- Poveda, E.; Barreiro, P.; Rodés, B.; Soriano, V. Enfuvirtide is active against HIV type 1 group O. AIDS Res. Hum. Retrovir. 2005, 21, 583–585. [Google Scholar] [CrossRef]
- Unal, G.; Alessandri-Gradt, E.; Leoz, M.; Pavie, J.; Lefèvre, C.; Panjo, H.; Charpentier, C.; Descamps, D.; Barin, F.; Simon, F.; et al. Human Immunodeficiency Virus Type 1 Group O Infection in France: Clinical Features and Immunovirological Response to Antiretrovirals. Clin. Infect. Dis. 2017, 66, 1785–1793. [Google Scholar] [CrossRef] [Green Version]
- Kouanfack, C.; Unal, G.; Schaeffer, L.; Kfutwah, A.; Aghokeng, A.; Mougnutou, R.; Tchemgui-Noumsi, N.; Alessandri-Gradt, E.; Delaporte, E.; Simon, F.; et al. Comparative Immunovirological and Clinical Responses to Antiretroviral Therapy Between HIV-1 Group O and HIV-1 Group M Infected Patients. Clin. Infect. Dis. 2019, 70, 1471–1477. [Google Scholar] [CrossRef]
- Alessandri-Gradt, E.; Unal, G.; Leoz, M.; Plantier, J.-C. Virological response to integrase strand transfer inhibitor-based antiretroviral combinations in HIV-1 group O-infected patients. Aids 2019, 33, 1327–1333. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.; Unal, G.; Plantier, J.C.; Alessandri-Gradt, E. Bictegravir-based antiretroviral therapy in HIV-1 group O patients: Data from real-life bictegravir/emtricitabine/tenofovir alafenamide switches. J. Antimicrob. Chemother. 2022, 77, 2305–2307. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Time of Follow-Up | Number of Samples | Year of Last Sample | pVL | CD4+ | |
|---|---|---|---|---|---|
| (Months) | (Log10 cp/mL) | Cells/µL | |||
| BCF204 | 67 | 35 | 2012 | undetectable | 731 |
| RBF208 | 176 | 30 | 2018 | undetectable | 277 |
| BCF212 | 139 | 39 | 2018 | undetectable | 689 |
| RBF235 | 259 | 56 | 2021 | 3.7 | 365 |
| RBF243 | 270 | 27 | 2022 | undetectable | 621 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alessandri-Gradt, E.; Moisan, A.; Plantier, J.-C. HIV-1 Non-Group M Strains and ART. Viruses 2023, 15, 780. https://doi.org/10.3390/v15030780
Alessandri-Gradt E, Moisan A, Plantier J-C. HIV-1 Non-Group M Strains and ART. Viruses. 2023; 15(3):780. https://doi.org/10.3390/v15030780
Chicago/Turabian StyleAlessandri-Gradt, Elodie, Alice Moisan, and Jean-Christophe Plantier. 2023. "HIV-1 Non-Group M Strains and ART" Viruses 15, no. 3: 780. https://doi.org/10.3390/v15030780