
Effects of SARS-CoV-2 Omicron BA.1 Spike Mutations on T-Cell Epitopes in Mice
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Vaccine and Mice
2.2. Splenocyte Isolation
2.3. Peptide Synthesis and Peptide Pool Preparation
2.4. Screening of T-Cell Epitopes by ELISpot
2.5. Intracellular Cytokine Staining
2.6. Statistical Analysis
3. Results
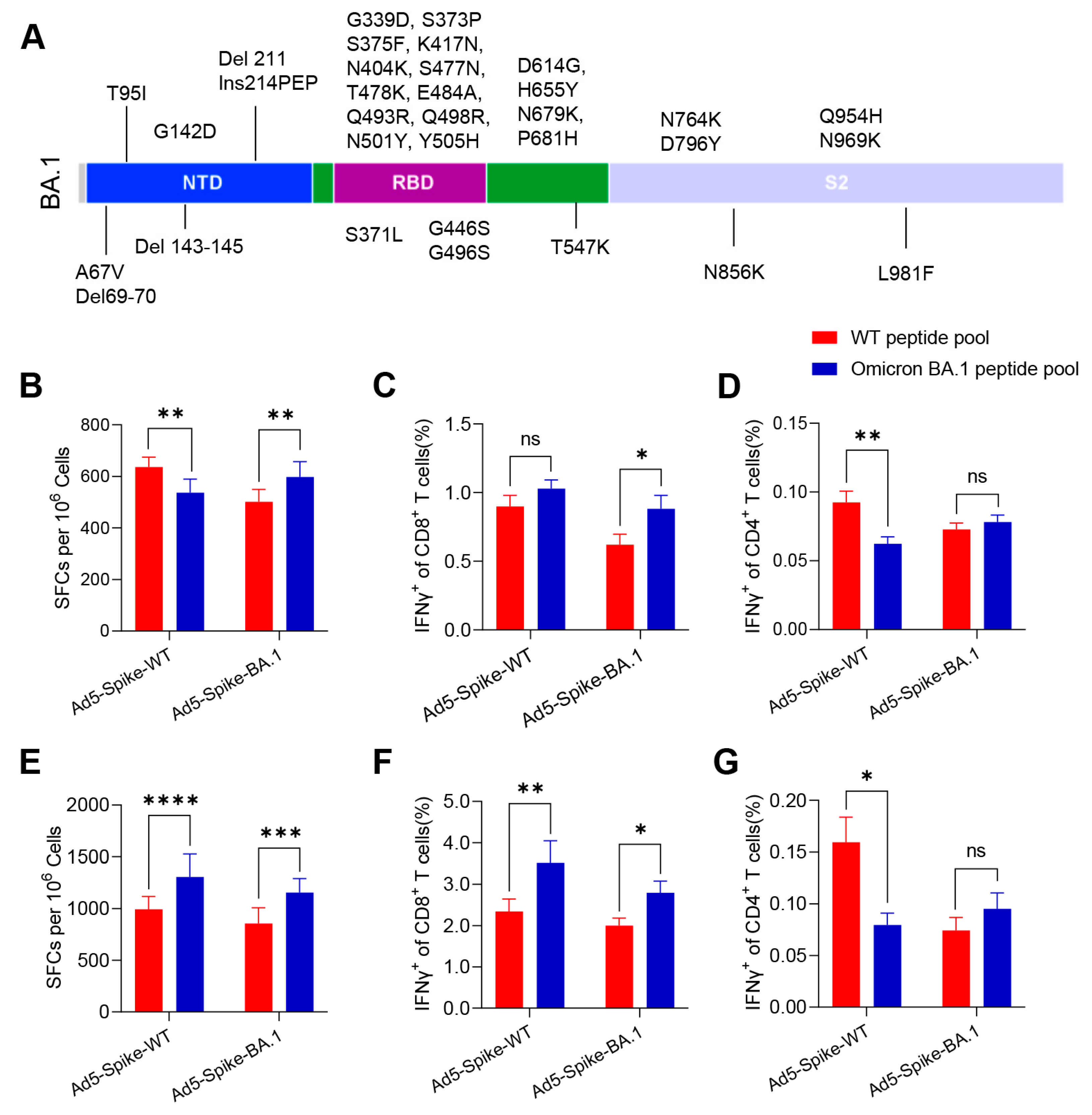
3.1. Ad5-Spike-WT and Ad5-Spike-BA.1 Induced Robust but Different Cellular Immune Responses in Mice
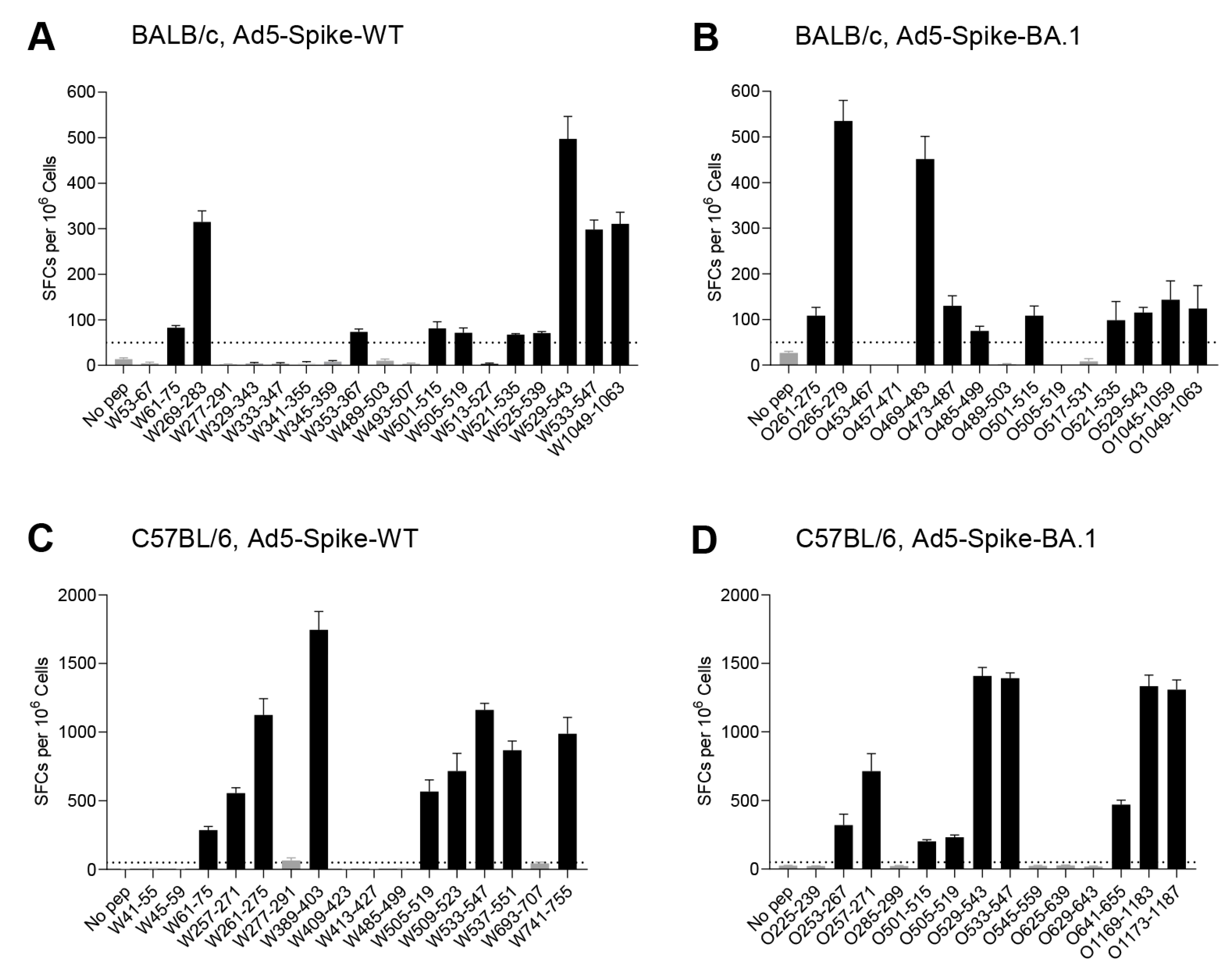
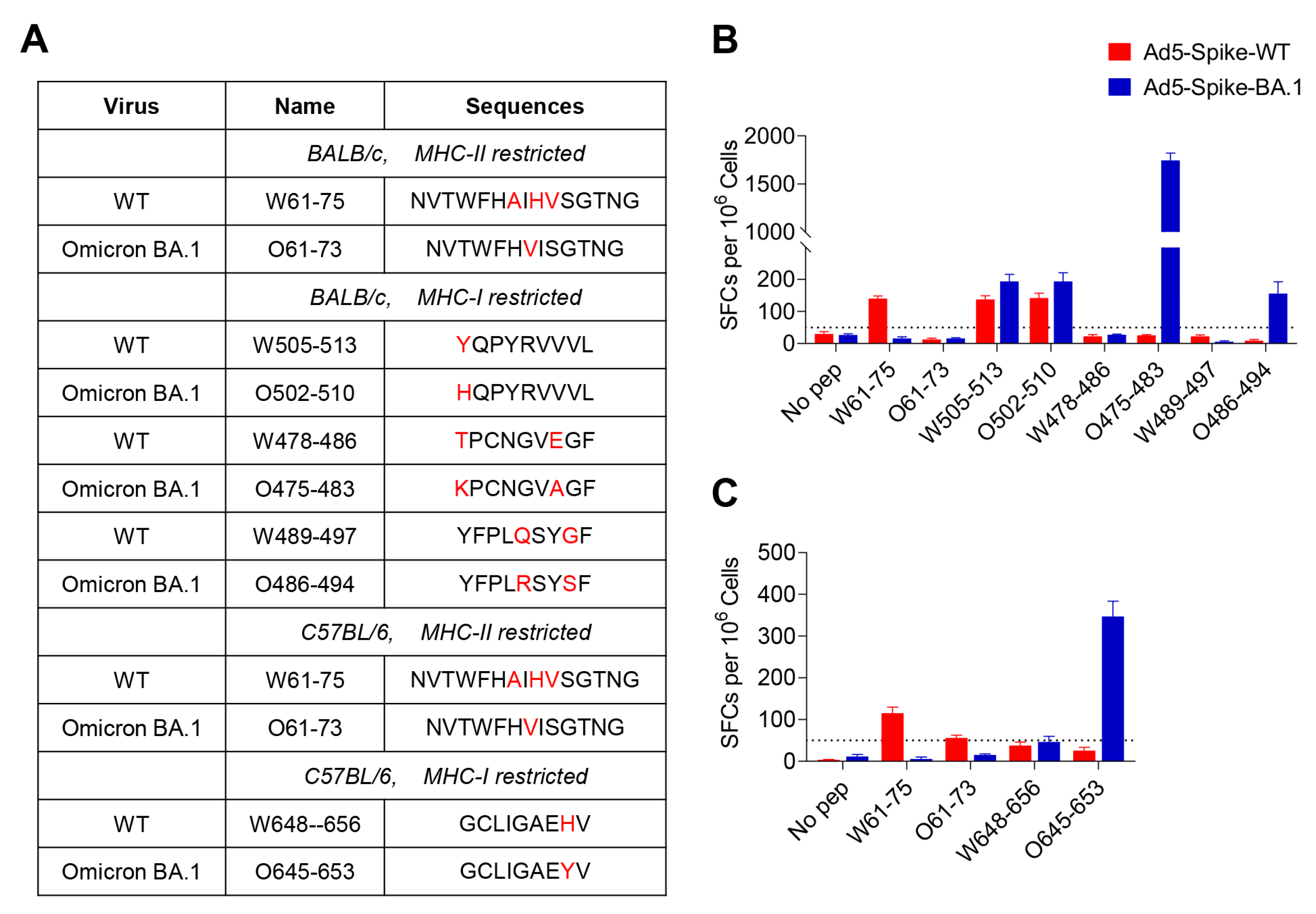
3.2. Identification of T-Cell Epitopes in WT and Omicron BA.1 Spike
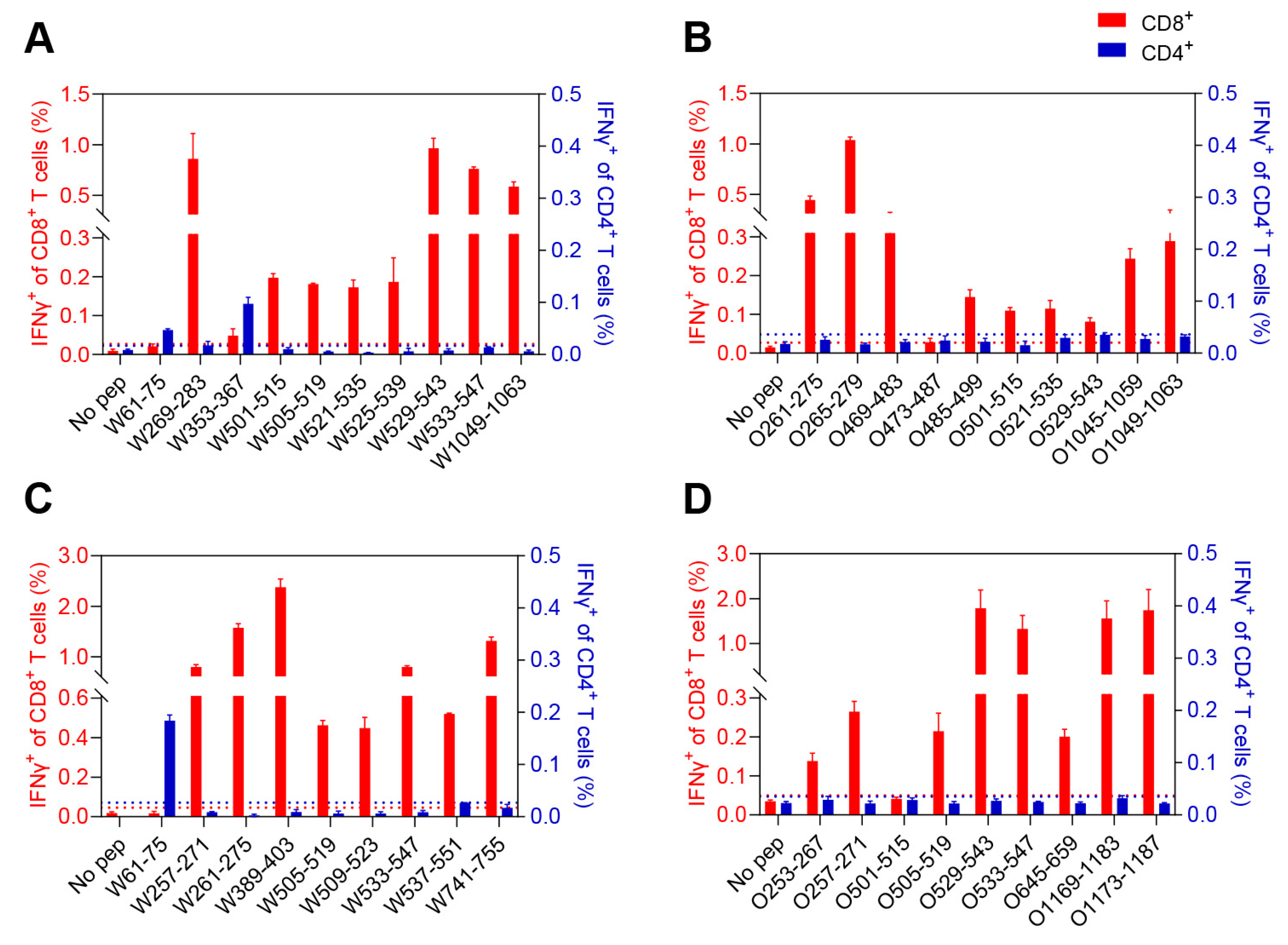
3.3. Characterization of the CD8+ and CD4+ T-Cell Epitopes
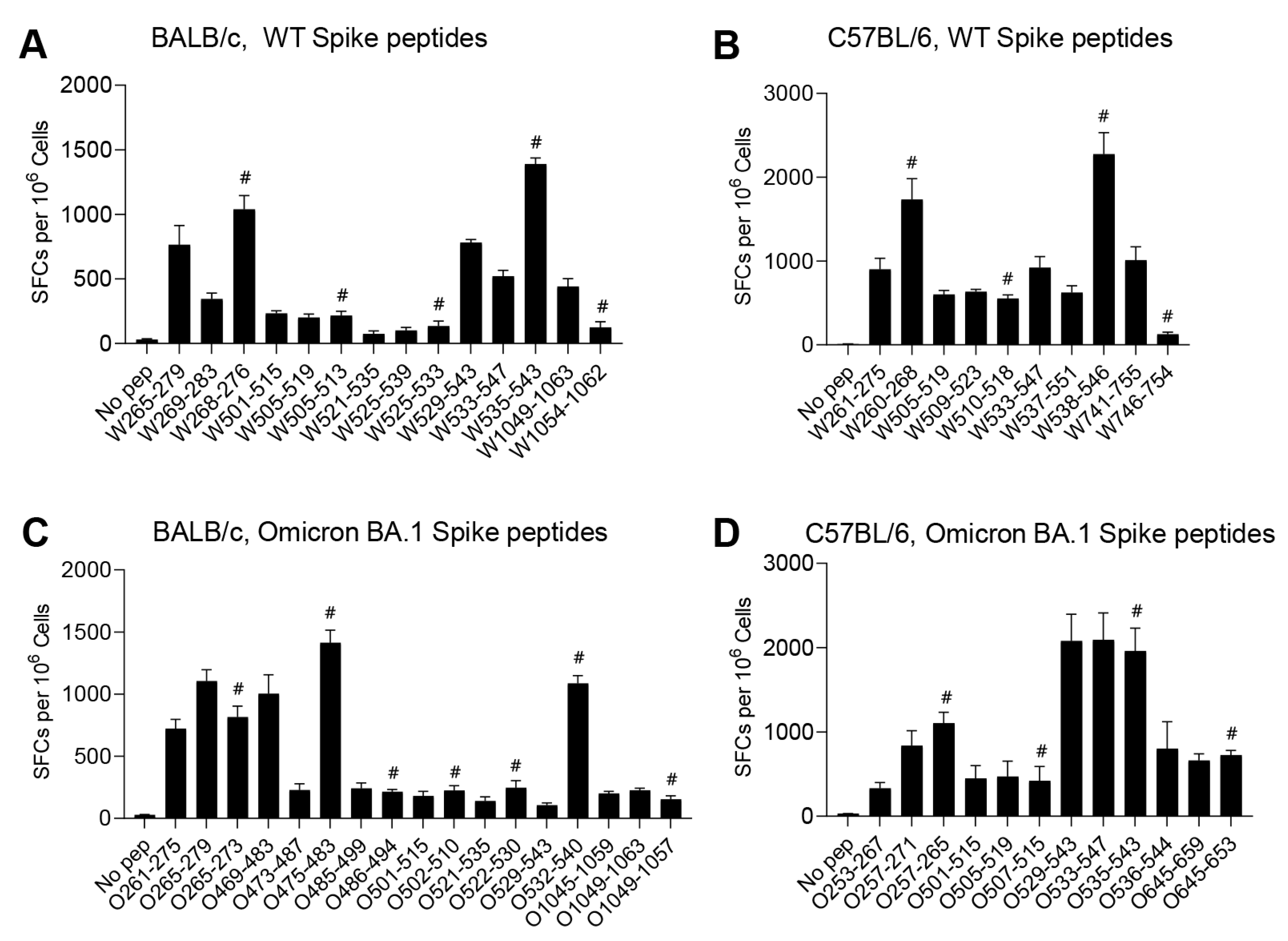
3.4. The Effect of Spike Mutations on T-Cell Epitopes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Tracking SARS-CoV-2 Variants. 2022. Available online: https://www.who.int/activities/tracking-SARS-CoV-2-variants/ (accessed on 15 September 2022).
- Uraki, R.; Ito, M.; Furusawa, Y.; Yamayoshi, S.; Iwatsuki-Horimoto, K.; Adachi, E.; Saito, M.; Koga, M.; Tsutsumi, T.; Yamamoto, S.; et al. Humoral immune evasion of the omicron subvariants BQ.1.1 and XBB. Lancet Infect. Dis. 2023, 23, 30–32. [Google Scholar] [CrossRef]
- Yue, C.; Song, W.; Wang, L.; Jian, F.; Chen, X.; Gao, F.; Shen, Z.; Wang, Y.; Wang, X.; Cao, Y. Enhanced transmissibility of XBB.1.5 is contributed by both strong ACE2 binding and antibody evasion. bioRxiv 2023. [Google Scholar] [CrossRef]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Sadoff, J.; Gray, G.; Vandebosch, A.; Cardenas, V.; Shukarev, G.; Grinsztejn, B.; Goepfert, P.A.; Truyers, C.; Fennema, H.; Spiessens, B.; et al. Safety and Efficacy of Single-Dose Ad26.COV2.S Vaccine against COVID-19. N. Engl. J. Med. 2021, 384, 2187–2201. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Marc, G.P.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA COVID-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Zhu, F.C.; Li, Y.H.; Guan, X.H.; Hou, L.H.; Wang, W.J.; Li, J.X.; Wu, S.P.; Wang, B.S.; Wang, Z.; Wang, L.; et al. Safety, tolerability, and immunogenicity of a recombinant adenovirus type-5 vectored COVID-19 vaccine: A dose-escalation, open-label, non-randomised, first-in-human trial. Lancet 2020, 395, 1845–1854. [Google Scholar] [CrossRef]
- Xia, S.; Wang, L.J.; Zhu, Y.; Lu, L.; Jiang, S.B. Origin, virological features, immune evasion and intervention of SARS-CoV-2 Omicron sublineages. Signal Transduct. Target. Ther. 2022, 7, 241. [Google Scholar] [CrossRef]
- Muik, A.; Lui, B.G.; Wallisch, A.K.; Bacher, M.; Muhl, J.; Reinholz, J.; Ozhelvaci, O.; Beckmann, N.; Garcia, R.D.G.; Poran, A.; et al. Neutralization of SARS-CoV-2 Omicron by BNT162b2 mRNA vaccine-elicited human sera. Science 2022, 375, 678–680. [Google Scholar] [CrossRef]
- Schmidt, F.; Muecksch, F.; Weisblum, Y.; Da Silva, J.; Bednarski, E.; Cho, A.L.; Wang, Z.J.; Gaebler, C.; Caskey, M.; Nussenzweig, M.C.; et al. Plasma Neutralization of the SARS-CoV-2 Omicron Variant. N. Engl. J. Med. 2022, 386, 599–601. [Google Scholar] [CrossRef]
- Rossler, A.; Riepler, L.; Bante, D.; von Laer, D.; Kimpel, J. SARS-CoV-2 Omicron Variant Neutralization in Serum from Vaccinated and Convalescent Persons. N. Engl. J. Med. 2022, 386, 698–700. [Google Scholar] [CrossRef]
- Nemet, I.; Kliker, L.; Lustig, Y.; Zuckerman, N.; Erster, O.; Cohen, C.; Kreiss, Y.; Alroy-Preis, S.; Regev-Yochay, G.; Mendelson, E.; et al. Third BNT162b2 Vaccination Neutralization of SARS-CoV-2 Omicron Infection. N. Engl. J. Med. 2022, 386, 492–494. [Google Scholar] [CrossRef]
- Wu, M.; Wall, E.C.; Carr, E.J.; Harvey, R.; Townsley, H.; Mears, H.V.; Adams, L.; Kjaer, S.; Kelly, G.; Warchal, S.; et al. Three-dose vaccination elicits neutralising antibodies against omicron. Lancet 2022, 399, 715–717. [Google Scholar] [CrossRef]
- Wang, Q.; Iketani, S.; Li, Z.; Liu, L.; Guo, Y.; Huang, Y.; Bowen, A.D.; Liu, M.; Wang, M.; Yu, J.; et al. Alarming antibody evasion properties of rising SARS-CoV-2 BQ and XBB subvariants. Cell 2023, 186, 279–286. [Google Scholar] [CrossRef]
- Lu, X.; Yamasaki, S. Current understanding of T cell immunity against SARS-CoV-2. Inflamm. Regen. 2022, 42, 51. [Google Scholar] [CrossRef]
- Tarke, A.; Sidney, J.; Kidd, C.K.; Dan, J.M.; Ramirez, S.I.; Yu, E.D.; Mateus, J.; da Silva Antunes, R.; Moore, E.; Rubiro, P.; et al. Comprehensive analysis of T cell immunodominance and immunoprevalence of SARS-CoV-2 epitopes in COVID-19 cases. Cell Rep. Med. 2021, 2, 100204. [Google Scholar] [CrossRef]
- Moss, P. The T cell immune response against SARS-CoV-2. Nat. Immunol. 2022, 23, 186–193. [Google Scholar] [CrossRef]
- Gao, Y.; Cai, C.; Grifoni, A.; Muller, T.R.; Niessl, J.; Olofsson, A.; Humbert, M.; Hansson, L.; Osterborg, A.; Bergman, P.; et al. Ancestral SARS-CoV-2-specific T cells cross-recognize the Omicron variant. Nat. Med. 2022, 28, 472–476. [Google Scholar] [CrossRef]
- GeurtsvanKessel, C.H.; Geers, D.; Schmitz, K.S.; Mykytyn, A.Z.; Lamers, M.M.; Bogers, S.; Scherbeijn, S.; Gommers, L.; Sablerolles, R.S.G.; Nieuwkoop, N.N.; et al. Divergent SARS-CoV-2 Omicron-reactive T and B cell responses in COVID-19 vaccine recipients. Sci. Immunol. 2022, 7, eabo2202. [Google Scholar] [CrossRef]
- Tarke, A.; Coelho, C.H.; Zhang, Z.L.; Dan, J.M.; Yu, E.D.; Methot, N.; Bloom, N.I.; Goodwin, B.; Phillips, E.; Mallal, S.; et al. SARS-CoV-2 vaccination induces immunological T cell memory able to cross-recognize variants from Alpha to Omicron. Cell 2022, 185, 847–859. [Google Scholar] [CrossRef]
- Naranbhai, V.; Nathan, A.; Kaseke, C.; Berrios, C.; Khatri, A.; Choi, S.; Getz, M.A.; Tano-Menka, R.; Ofoman, O.; Gayton, A. T cell reactivity to the SARS-CoV-2 Omicron variant is preserved in most but not all individuals. Cell 2022, 185, 1041–1051. [Google Scholar] [CrossRef]
- Keeton, R.; Tincho, M.B.; Ngomti, A.; Baguma, R.; Benede, N.; Suzuki, A.; Khan, K.; Cele, S.; Bernstein, M.; Karim, F.; et al. T cell responses to SARS-CoV-2 spike cross-recognize Omicron. Nature 2022, 603, 488–492. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Y.; Chandrashekar, A.; Sellers, D.; Barrett, J.; Jacob-Dolan, C.; Lifton, M.; McMahan, K.; Sciacca, M.; VanWyk, H.; Wu, C.; et al. Vaccines elicit highly conserved cellular immunity to SARS-CoV-2 Omicron. Nature 2022, 603, 493–496. [Google Scholar] [CrossRef]
- Emmelot, M.E.; Vos, M.; Boer, M.C.; Rots, N.Y.; de Wit, J.; van Els, C.A.C.M.; Kaaijk, P. Omicron BA.1 Mutations in SARS-CoV-2 Spike Lead to Reduced T-Cell Response in Vaccinated and Convalescent Individuals. Viruses 2022, 14, 1570. [Google Scholar] [CrossRef]
- Wang, B.; Xu, J.H.; Wu, S.P.; Zhang, Z.; Zhao, Z.H.; Zhang, J.; Fu, L.; Zai, X.D.; Wang, Y.D.; Zhang, G.Y.; et al. Comparative characterization of antibody responses induced by Ad5-vectored spike proteins of emerging SARS-CoV-2 VOCs. Signal Transduct. Target. Ther. 2022, 7, 257. [Google Scholar] [CrossRef]
- Wu, S.P.; Yu, T.; Song, X.H.; Yi, S.Q.; Hou, L.H.; Chen, W. Prediction and identification of mouse cytotoxic T lymphocyte epitopes in Ebola virus glycoproteins. Virol. J. 2012, 9, 111. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Zhong, G.; Zhang, J.; Shuai, L.; Zhang, Z.; Wen, Z.; Wang, B.; Zhao, Z.; Song, X.; Chen, Y.; et al. A single dose of an adenovirus-vectored vaccine provides protection against SARS-CoV-2 challenge. Nat. Commun. 2020, 11, 4081. [Google Scholar] [CrossRef]
- Laczkó, D.; Hogan, M.J.; Toulmin, S.A.; Hicks, P.; Lederer, K.; Gaudette, B.T.; Castaño, D.; Amanat, F.; Muramatsu, H.; Oguin, T.H.; et al. A Single Immunization with Nucleoside-Modified mRNA Vaccines Elicits Strong Cellular and Humoral Immune Responses against SARS-CoV-2 in Mice. Immunity 2020, 53, 724–732. [Google Scholar] [CrossRef]
- Graham, S.P.; McLean, R.K.; Spencer, A.J.; Belij-Rammerstorfer, S.; Wright, D.; Ulaszewska, M.; Edwards, J.C.; Hayes, J.W.P.; Martini, V.; Thakur, N.; et al. Evaluation of the immunogenicity of prime-boost vaccination with the replication-deficient viral vectored COVID-19 vaccine candidate ChAdOx1 nCoV-19. NPJ Vaccines 2020, 5, 69. [Google Scholar] [CrossRef]
- Liu, J.; Budylowski, P.; Samson, R.; Griffin, B.D.; Babuadze, G.; Rathod, B.; Colwill, K.; Abioye, J.A.; Schwartz, J.A.; Law, R.; et al. Preclinical evaluation of a SARS-CoV-2 mRNA vaccine PTX-COVID19-B. Sci. Adv. 2022, 8, eabj9815. [Google Scholar] [CrossRef]
- Goel, R.R.; Painter, M.M.; Apostolidis, S.A.; Mathew, D.; Meng, W.; Rosenfeld, A.M.; Lundgreen, K.A.; Reynaldi, A.; Khoury, D.S.; Pattekar, A.; et al. mRNA vaccines induce durable immune memory to SARS-CoV-2 and variants of concern. Science 2021, 374, abm0829. [Google Scholar] [CrossRef]
- Liwsrisakun, C.; Pata, S.; Laopajon, W.; Takheaw, N.; Chaiwong, W.; Duangjit, P.; Inchai, J.; Chuensirikulchai, K.; Cheyasawan, P.; Pothirat, C.; et al. Humoral and cellular immune responses against SARS-CoV-2 variants of concern induced by heterologous CoronaVac/ChAdOx-1 versus homologous ChAdOx-1 vaccination in the elderly. Asian Pac. J. Allergy Immunol. 2023. [Google Scholar] [CrossRef]
- Park, K.; Choi, S.J.; Shin, E.-C. Omicron Subvariants, Including BA.4 and BA.5, Substantially Preserve T Cell Epitopes of Ancestral SARS-CoV-2. Immune Netw. 2022, 22, e29. [Google Scholar] [CrossRef]
- Li, H.; Chen, Z.; Liu, X.; Hu, P. T cell epitopes are largely conserved in the SARS-CoV-2 Omicron subvariant (BA.1, BA.2, BA.3, and GKA). J. Med. Virol. 2022, 94, 4591–4592. [Google Scholar] [CrossRef]
- Choi, S.J.; Kim, D.-U.; Noh, J.Y.; Kim, S.; Park, S.-H.; Jeong, H.W.; Shin, E.-C. T cell epitopes in SARS-CoV-2 proteins are substantially conserved in the Omicron variant. Cell Mol. Immunol. 2022, 19, 447–448. [Google Scholar] [CrossRef]
- Dolton, G.; Rius, C.; Hasan, M.S.; Wall, A.; Szomolay, B.; Behiry, E.; Whalley, T.; Southgate, J.; Fuller, A.; Morin, T.; et al. Emergence of immune escape at dominant SARS-CoV-2 killer T cell epitope. Cell 2022, 185, 2936–2951. [Google Scholar] [CrossRef]
- Davenport, B.J.; Morrison, T.E.; Kedl, R.M.; Klarquist, J. Conserved and Novel Mouse CD8 T Cell Epitopes within SARS-CoV-2 Spike Receptor Binding Domain Protein Identified following Subunit Vaccination. J. Immunol. 2021, 206, 2503–2507. [Google Scholar] [CrossRef]
- Zhuang, Z.; Lai, X.; Sun, J.; Chen, Z.; Zhang, Z.; Dai, J.; Liu, D.; Li, Y.; Li, F.; Wang, Y.; et al. Mapping and role of T cell response in SARS-CoV-2-infected mice. J. Exp. Med. 2021, 218, e20202187. [Google Scholar] [CrossRef]
- Pardieck, I.N.; van der Sluis, T.C.; van der Gracht, E.T.I.; Veerkamp, D.M.B.; Behr, F.M.; van Duikeren, S.; Beyrend, G.; Rip, J.; Nadafi, R.; Beyranvand Nejad, E.; et al. A third vaccination with a single T cell epitope confers protection in a murine model of SARS-CoV-2 infection. Nat. Commun. 2022, 13, 3966. [Google Scholar] [CrossRef]
- Ju, B.; Fan, Q.; Wang, M.; Liao, X.; Guo, H.; Wang, H.; Ge, X.; Liu, L.; Zhang, Z. Antigenic sin of wild-type SARS-CoV-2 vaccine shapes poor cross-neutralization of BA.4/5/2.75 subvariants in BA.2 breakthrough infections. Nat. Commun. 2022, 13, 7120. [Google Scholar] [CrossRef]
- Gagne, M.; Moliva, J.I.; Foulds, K.E.; Andrew, S.F.; Flynn, B.J.; Werner, A.P.; Wagner, D.A.; Teng, I.T.; Lin, B.C.; Moore, C.; et al. mRNA-1273 or mRNA-Omicron boost in vaccinated macaques elicits similar B cell expansion, neutralizing responses, and protection from Omicron. Cell 2022, 185, 1556–1571.e18. [Google Scholar] [CrossRef]
- Kaku, C.I.; Bergeron, A.J.; Ahlm, C.; Normark, J.; Sakharkar, M.; Forsell, M.N.E.; Walker, L.M. Recall of preexisting cross-reactive B cell memory after Omicron BA.1 breakthrough infection. Sci. Immunol. 2022, 7, eabq3511. [Google Scholar] [CrossRef] [PubMed]
- Zehn, D.; Turner, M.J.; Lefrançois, L.; Bevan, M.J. Lack of original antigenic sin in recall CD8(+) T cell responses. J. Immunol. 2010, 184, 6320–6326. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Epitope | Protein | Start Position | End Position | CD4/CD8 | MHC |
|---|---|---|---|---|---|---|
| BALB/c | ||||||
| W61-75 | NVTWFHAIHVSGTNG | WT | 61 | 75 | CD4 | I-Ed |
| W353-367 | WNRKRISNCVADYSV | WT/Omicron BA.1 | 353 | 367 | CD4 | I-Ed |
| W268-276 | GYLQPRTFL | WT/Omicron BA.1 | 268 | 276 | CD8 | H2-Ld |
| W505-513 | YQPYRVVVL | WT | 505 | 513 | CD8 | H2-Kd |
| W525-533 | CGPKKSTNL | WT/Omicron BA.1 | 525 | 533 | CD8 | H2-Dd |
| W535-543 | KNKCVNFNF | WT/Omicron BA.1 | 535 | 543 | CD8 | H2-Ld |
| W1054-1062 | QSAPHGVVF | WT/Omicron BA.1 | 1054 | 1062 | CD8 | H2-Dd |
| O475-483 | KPCNGVAGF | Omicron BA.1 | 475 | 483 | CD8 | H2-Ld |
| O486-494 | YFPLRSYSF | Omicron BA.1 | 486 | 494 | CD8 | H2-Ld |
| O502-510 | HQPYRVVVL | Omicron BA.1 | 502 | 510 | CD8 | H2-Dd |
| O1049-1057 | FPQSAPHGV | WT/Omicron BA.1 | 1049 | 1057 | CD8 | H2-Kd |
| C57BL/6 | ||||||
| W61-75 | NVTWFHAIHVSGTNG | WT | 61 | 75 | CD4 | I-Ab |
| W260-268 | AGAAAYYVG | WT/Omicron BA.1 | 260 | 268 | CD8 | H2-Kb |
| W389-403 | DLCFTNVYADSFVIR | WT/Omicron BA.1 | 389 | 403 | CD8 | H-2-Db |
| W510-518 | VVVLSFELL | WT/Omicron BA.1 | 510 | 518 | CD8 | H2-Kb |
| W538-546 | CVNFNFNGL | WT/Omicron BA.1 | 538 | 546 | CD8 | H2-Kb |
| W746-754 | STECSNLLL | WT/Omicron BA.1 | 746 | 754 | CD8 | H2-Db |
| O506-514 | RVVVLSFEL | WT/Omicron BA.1 | 506 | 514 | CD8 | H2-Kb |
| O645-653 | GCLIGAEYV | Omicron BA.1 | 645 | 653 | CD8 | H2-Db |
| O1173-1187 | VVNIQKEIDRLNEVA | WT/Omicron BA.1 | 1173 | 1187 | CD8 | H2-Db |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Wang, B.; Zhao, Z.; Xu, J.; Zhang, Z.; Zhang, J.; Chen, Y.; Song, X.; Zheng, W.; Hou, L.; et al. Effects of SARS-CoV-2 Omicron BA.1 Spike Mutations on T-Cell Epitopes in Mice. Viruses 2023, 15, 763. https://doi.org/10.3390/v15030763
Wang Y, Wang B, Zhao Z, Xu J, Zhang Z, Zhang J, Chen Y, Song X, Zheng W, Hou L, et al. Effects of SARS-CoV-2 Omicron BA.1 Spike Mutations on T-Cell Epitopes in Mice. Viruses. 2023; 15(3):763. https://doi.org/10.3390/v15030763
Chicago/Turabian StyleWang, Yudong, Busen Wang, Zhenghao Zhao, Jinghan Xu, Zhe Zhang, Jinlong Zhang, Yi Chen, Xiaohong Song, Wanru Zheng, Lihua Hou, and et al. 2023. "Effects of SARS-CoV-2 Omicron BA.1 Spike Mutations on T-Cell Epitopes in Mice" Viruses 15, no. 3: 763. https://doi.org/10.3390/v15030763