
SARS-CoV-2 Is More Efficient than HCoV-NL63 in Infecting a Small Subpopulation of ACE2+ Human Respiratory Epithelial Cells
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Primary Human Respiratory Epithelial Cells (HREC)
2.2. LLC-MK2 Cells
2.3. HCoV-NL63 Culture and Propagation
2.4. SARS-CoV-2 Culture and Propagation In Vitro
2.5. HCoV-NL63 and SARS-CoV-2 Infectious Studies in HRECs
2.6. Immunohistochemistry Staining in Tissues and Cell Cultures
2.7. Cellular Characterization Using Flow Cytometry
2.8. Viral Binding Assay
2.9. Reverse Transcription-qPCR (RT-qPCR) Assay
2.10. Data Analysis
3. Results
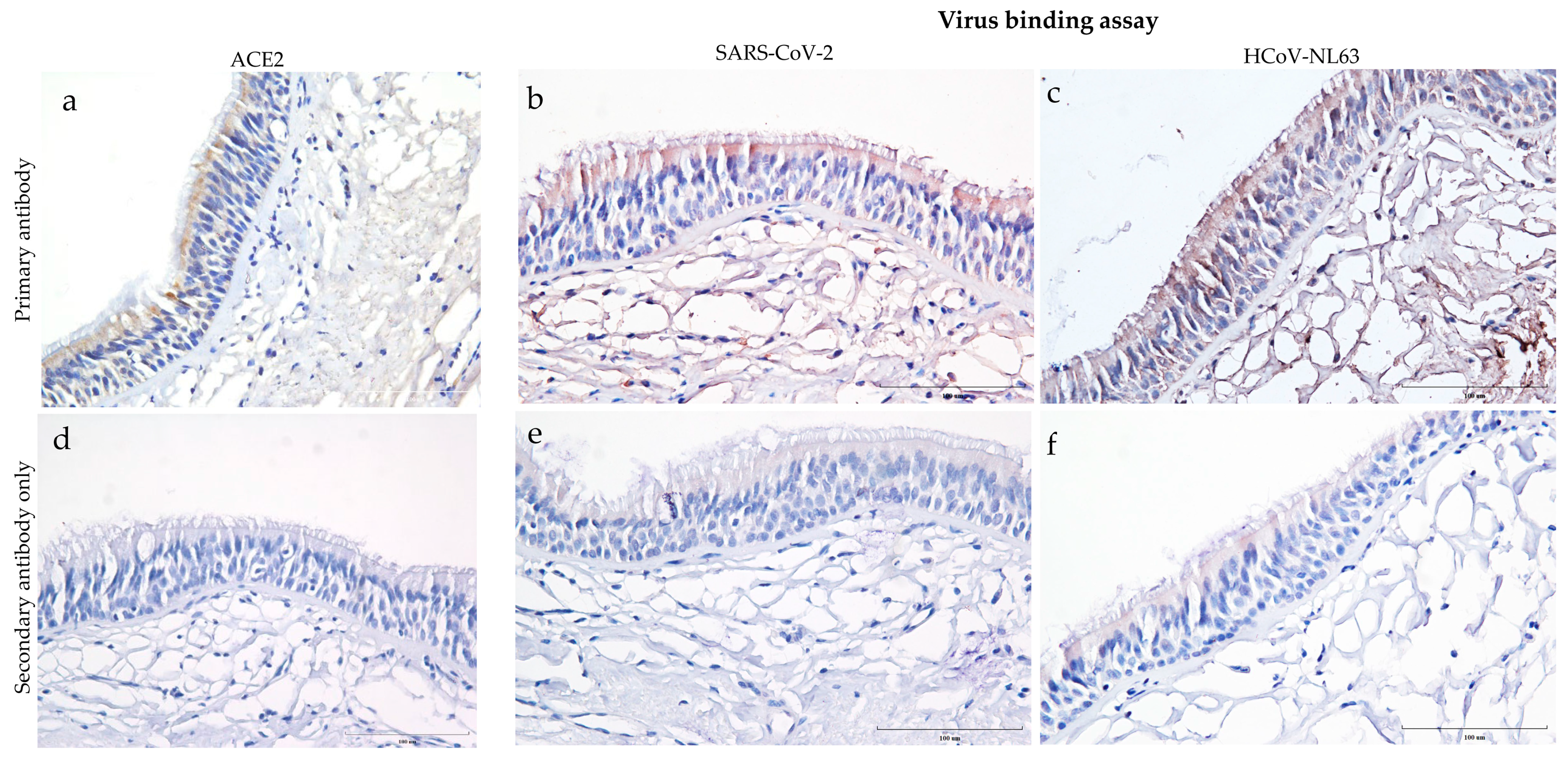
3.1. Distribution of ACE2 Receptor and Virus Binding on the Human Trachea
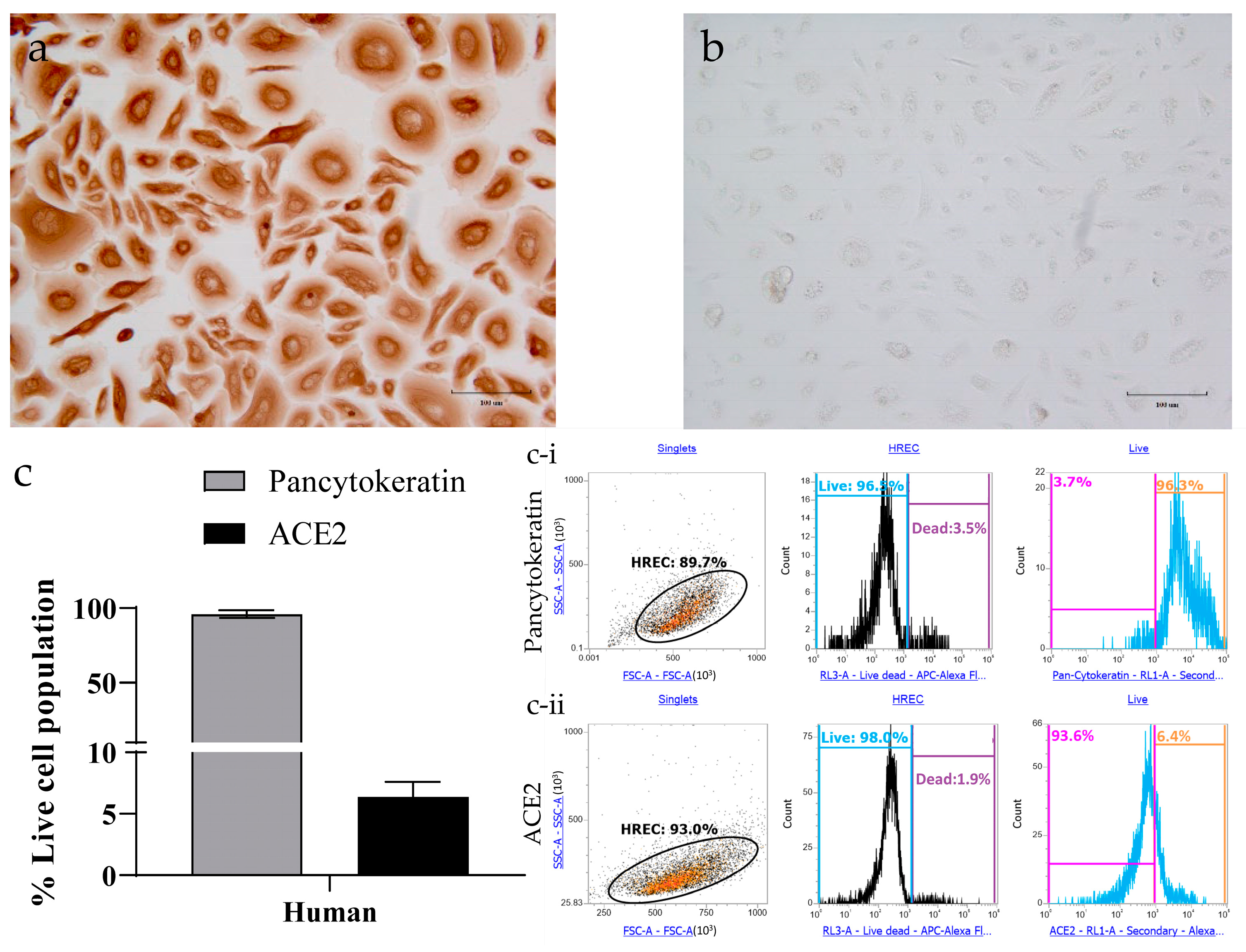
3.2. Determining ACE2 Receptors in Human Respiratory Epithelial Cells
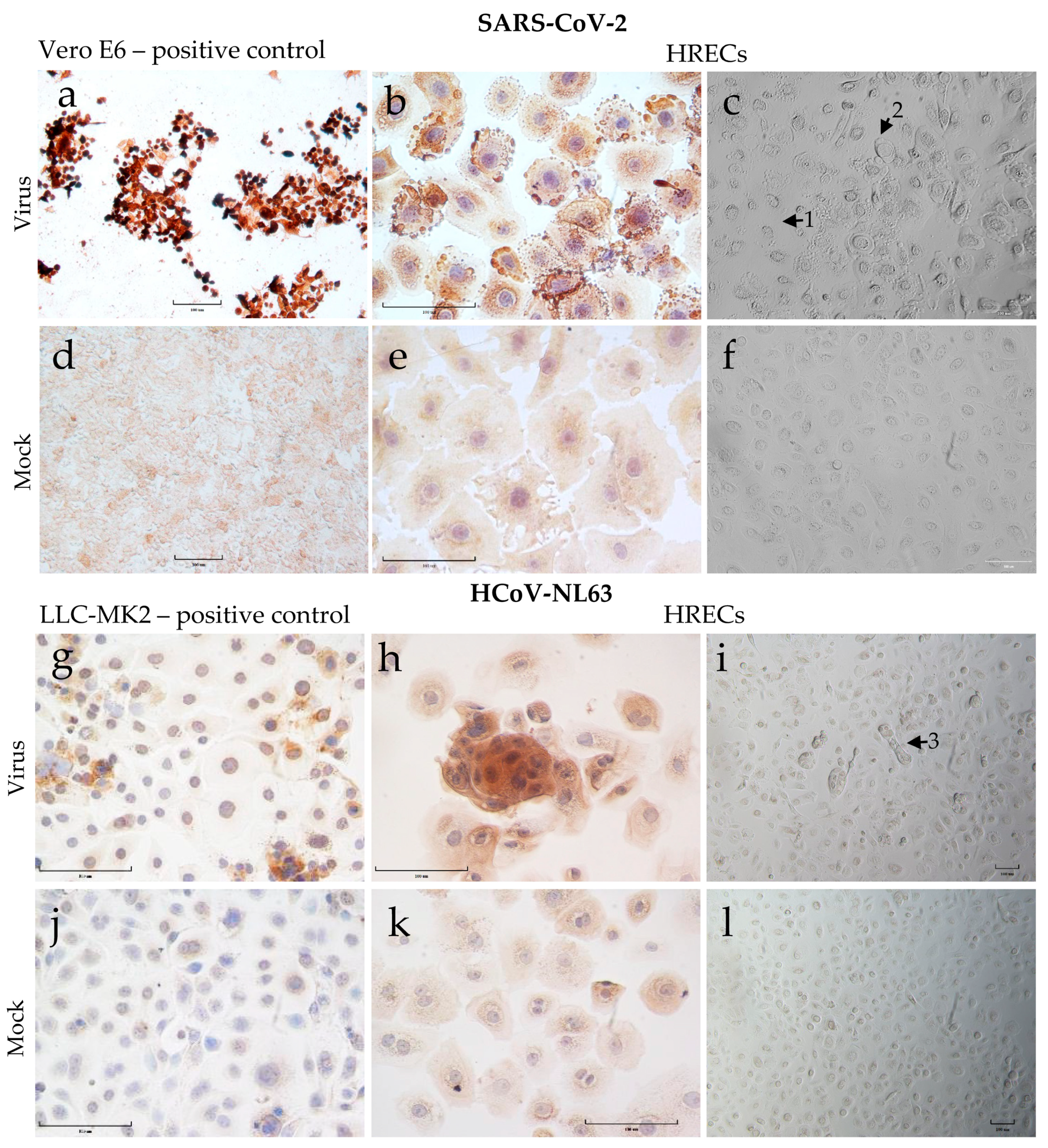
3.3. Demonstration of SARS-CoV-2 and HCoV-NL63 Virus Infection in HRECs by IHC
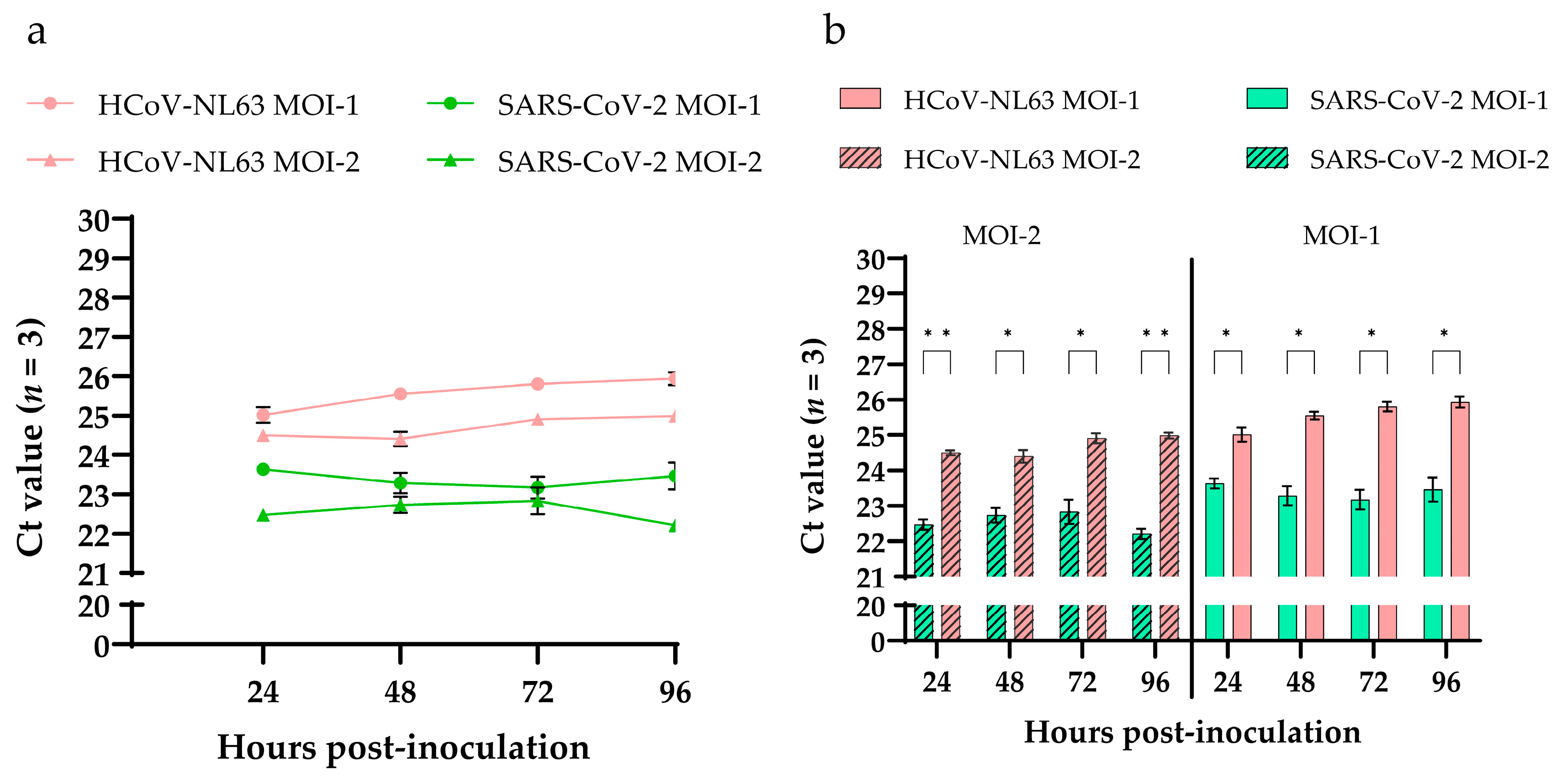
3.4. SARS-CoV-2 and HCoV-NL63 Virus Replication Kinetics in HRECs Using RT-qPCR
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fielding, B.C. Human Coronavirus NL63: A Clinically Important Virus? Future Microbiol. 2011, 6, 153–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peiris, J.; Lai, S.; Poon, L.; Guan, Y.; Yam, L.; Lim, W.; Nicholls, J.; Yee, W.; Yan, W.; Cheung, M.; et al. Coronavirus as a Possible Cause of Severe Acute Respiratory Syndrome. Lancet 2003, 361, 1319–1325. [Google Scholar] [CrossRef] [Green Version]
- Zaki, A.M.; van Boheemen, S.; Bestebroer, T.M.; Osterhaus, A.D.M.E.; Fouchier, R.A.M. Isolation of a Novel Coronavirus from a Man with Pneumonia in Saudi Arabia. N. Engl. J. Med. 2012, 367, 1814–1820. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Moore, M.J.; Vasilieva, N.; Sui, J.; Wong, S.K.; Berne, M.A.; Somasundaran, M.; Sullivan, J.L.; Luzuriaga, K.; Greenough, T.C.; et al. Angiotensin-Converting Enzyme 2 Is a Functional Receptor for the SARS Coronavirus. Nature 2003, 426, 450–454. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, H.; Pyrc, K.; van der Hoek, L.; Geier, M.; Berkhout, B.; Pöhlmann, S. Human Coronavirus NL63 Employs the Severe Acute Respiratory Syndrome Coronavirus Receptor for Cellular Entry. Proc. Natl. Acad. Sci. USA 2005, 102, 7988–7993. [Google Scholar] [CrossRef] [Green Version]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A Pneumonia Outbreak Associated with a New Coronavirus of Probable Bat Origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Belouzard, S.; Millet, J.K.; Licitra, B.N.; Whittaker, G.R. Mechanisms of Coronavirus Cell Entry Mediated by the Viral Spike Protein. Viruses 2012, 4, 1011–1033. [Google Scholar] [CrossRef] [Green Version]
- Millet, J.K.; Whittaker, G.R. Host Cell Proteases: Critical Determinants of Coronavirus Tropism and Pathogenesis. Virus Res. 2015, 202, 120–134. [Google Scholar] [CrossRef]
- Tikellis, C.; Thomas, M.C. Angiotensin-Converting Enzyme 2 (ACE2) Is a Key Modulator of the Renin Angiotensin System in Health and Disease. Int. J. Pept. 2012, 2012, 256294. [Google Scholar] [CrossRef]
- Wu, K.; Chen, L.; Peng, G.; Zhou, W.; Pennell, C.A.; Mansky, L.M.; Geraghty, R.J.; Li, F. A Virus-Binding Hot Spot on Human Angiotensin-Converting Enzyme 2 Is Critical for Binding of Two Different Coronaviruses. J. Virol. 2011, 85, 5331–5337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Struck, A.-W.; Axmann, M.; Pfefferle, S.; Drosten, C.; Meyer, B. A Hexapeptide of the Receptor-Binding Domain of SARS Corona Virus Spike Protein Blocks Viral Entry into Host Cells via the Human Receptor ACE2. Antivir. Res. 2012, 94, 288–296. [Google Scholar] [CrossRef] [PubMed]
- van der Hoek, L.; Pyrc, K.; Jebbink, M.F.; Vermeulen-Oost, W.; Berkhout, R.J.M.; Wolthers, K.C.; Wertheim-van Dillen, P.M.E.; Kaandorp, J.; Spaargaren, J.; Berkhout, B. Identification of a New Human Coronavirus. Nat. Med. 2004, 10, 368–373. [Google Scholar] [CrossRef] [PubMed]
- Harcourt, J.; Tamin, A.; Lu, X.; Kamili, S.; Sakthivel, S.K.; Murray, J.; Queen, K.; Tao, Y.; Paden, C.R.; Zhang, J.; et al. Isolation and Characterization of SARS-CoV-2 from the First US COVID-19 Patient. bioRxiv 2020. preprint. [Google Scholar] [CrossRef] [Green Version]
- Nelli, R.K.; Phadke, K.S.; Castillo, G.; Yen, L.; Saunders, A.; Rauh, R.; Nelson, W.; Bellaire, B.H.; Giménez-Lirola, L.G. Enhanced Apoptosis as a Possible Mechanism to Self-Limit SARS-CoV-2 Replication in Porcine Primary Respiratory Epithelial Cells in Contrast to Human Cells. Cell Death Discov. 2021, 7, 383. [Google Scholar] [CrossRef] [PubMed]
- Schildgen, O.; Jebbink, M.F.; de Vries, M.; Pyrc, K.; Dijkman, R.; Simon, A.; Müller, A.; Kupfer, B.; van der Hoek, L. Identification of Cell Lines Permissive for Human Coronavirus NL63. J. Virol. Methods 2006, 138, 207–210. [Google Scholar] [CrossRef]
- Castillo, G.; Mora-Díaz, J.C.; Nelli, R.K.; Giménez-Lirola, L.G. Human Air-Liquid-Interface Organotypic Airway Cultures Express Significantly More ACE2 Receptor Protein and Are More Susceptible to HCoV-NL63 Infection than Monolayer Cultures of Primary Respiratory Epithelial Cells. Microbiol. Spectr. 2022, 10, e01639-22. [Google Scholar] [CrossRef]
- Liu, Y.; Hu, G.; Wang, Y.; Ren, W.; Zhao, X.; Ji, F.; Zhu, Y.; Feng, F.; Gong, M.; Ju, X.; et al. Functional and Genetic Analysis of Viral Receptor ACE2 Orthologs Reveals a Broad Potential Host Range of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2021, 118, e2025373118. [Google Scholar] [CrossRef]
- Lan, J.; Chen, P.; Liu, W.; Ren, W.; Zhang, L.; Ding, Q.; Zhang, Q.; Wang, X.; Ge, J. Structural Insights into the Binding of SARS-CoV-2, SARS-CoV, and HCoV-NL63 Spike Receptor-Binding Domain to Horse ACE2. Structure 2022, 30, 1432–1442.e4. [Google Scholar] [CrossRef]
- Herzog, P.; Drosten, C.; Müller, M.A. Plaque Assay for Human Coronavirus NL63 Using Human Colon Carcinoma Cells. Virol. J. 2008, 5, 138. [Google Scholar] [CrossRef] [Green Version]
- Zhu, N.; Wang, W.; Liu, Z.; Liang, C.; Wang, W.; Ye, F.; Huang, B.; Zhao, L.; Wang, H.; Zhou, W.; et al. Morphogenesis and Cytopathic Effect of SARS-CoV-2 Infection in Human Airway Epithelial Cells. Nat. Commun. 2020, 11, 3910. [Google Scholar] [CrossRef] [PubMed]
- Dijkman, R.; Jebbink, M.F.; Koekkoek, S.M.; Deijs, M.; Jónsdóttir, H.R.; Molenkamp, R.; Ieven, M.; Goossens, H.; Thiel, V.; van der Hoek, L. Isolation and Characterization of Current Human Coronavirus Strains in Primary Human Epithelial Cell Cultures Reveal Differences in Target Cell Tropism. J. Virol. 2013, 87, 6081–6090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, I.T.; Nakayama, T.; Wu, C.T.; Goltsev, Y.; Jiang, S.; Gall, P.A.; Liao, C.K.; Shih, L.C.; Schürch, C.M.; McIlwain, D.R.; et al. Robust ACE2 protein expression localizes to the motile cilia of the respiratory tract epithelia and is not increased by ACE inhibitors or angiotensin receptor blockers. Nat. Commun. 2020, 11, 5453. [Google Scholar] [CrossRef]
- Bukowy-Bieryłło, Z. Long-term differentiating primary human airway epithelial cell cultures: How far are we? Cell Commun. Signal. 2021, 19, 63. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Oligo | Sequence |
|---|---|---|
| SARS-CoV-2 N1 | Forward | GAC CCC AAA ATC AGC GAA AT |
| Reverse | TCT GGT TAC TGC CAG TTG AAT CTG | |
| Probe | FAM-ACC CCG CAT TAC GTT TGG TGG ACC-BHQ1 | |
| SARS-CoV-2 N2 | Forward | TTA CAA ACA TTG GCC GCA AA |
| Reverse | GCG CGA CAT TCC GAA GAA | |
| Probe | HEX-ACA ATT TGC CCC CAG CGC TTC AG-BHQ1 | |
| HCoV-NL63 N [16] | Forward | GCGTGTTCCTACCAGAGAGGA |
| Reverse | GCTGTGGAAAACCTTTGGCA | |
| Probe | FAM-ATGTTATTCAGTGCTTTGGTCCTCGTGAT-BHQ1 | |
| Human RNase P | Forward | AGA TTT GGA CCT GCG AGC G |
| Reverse | GAG CGG CTG TCT CCA CAA GT | |
| Probe | Cy5-TTC TGA CCT GAA GGC TCT GCG CG-BHQ-1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castillo, G.; Nelli, R.K.; Phadke, K.S.; Bravo-Parra, M.; Mora-Díaz, J.C.; Bellaire, B.H.; Giménez-Lirola, L.G. SARS-CoV-2 Is More Efficient than HCoV-NL63 in Infecting a Small Subpopulation of ACE2+ Human Respiratory Epithelial Cells. Viruses 2023, 15, 736. https://doi.org/10.3390/v15030736
Castillo G, Nelli RK, Phadke KS, Bravo-Parra M, Mora-Díaz JC, Bellaire BH, Giménez-Lirola LG. SARS-CoV-2 Is More Efficient than HCoV-NL63 in Infecting a Small Subpopulation of ACE2+ Human Respiratory Epithelial Cells. Viruses. 2023; 15(3):736. https://doi.org/10.3390/v15030736
Chicago/Turabian StyleCastillo, Gino, Rahul K. Nelli, Kruttika S. Phadke, Marlene Bravo-Parra, Juan Carlos Mora-Díaz, Bryan H. Bellaire, and Luis G. Giménez-Lirola. 2023. "SARS-CoV-2 Is More Efficient than HCoV-NL63 in Infecting a Small Subpopulation of ACE2+ Human Respiratory Epithelial Cells" Viruses 15, no. 3: 736. https://doi.org/10.3390/v15030736