
Cis-Allosteric Regulation of HIV-1 Reverse Transcriptase by Integrase

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Construction of Expression Vectors
2.2. Preparation of Recombinant RTs
2.3. Proteolysis of RTIN by HIV-1 Protease
2.4. In Vitro Reverse Transcription Assay
2.5. Reagents
2.6. In Silico Analysis of HIV-1 RT and RTIN
3. Results
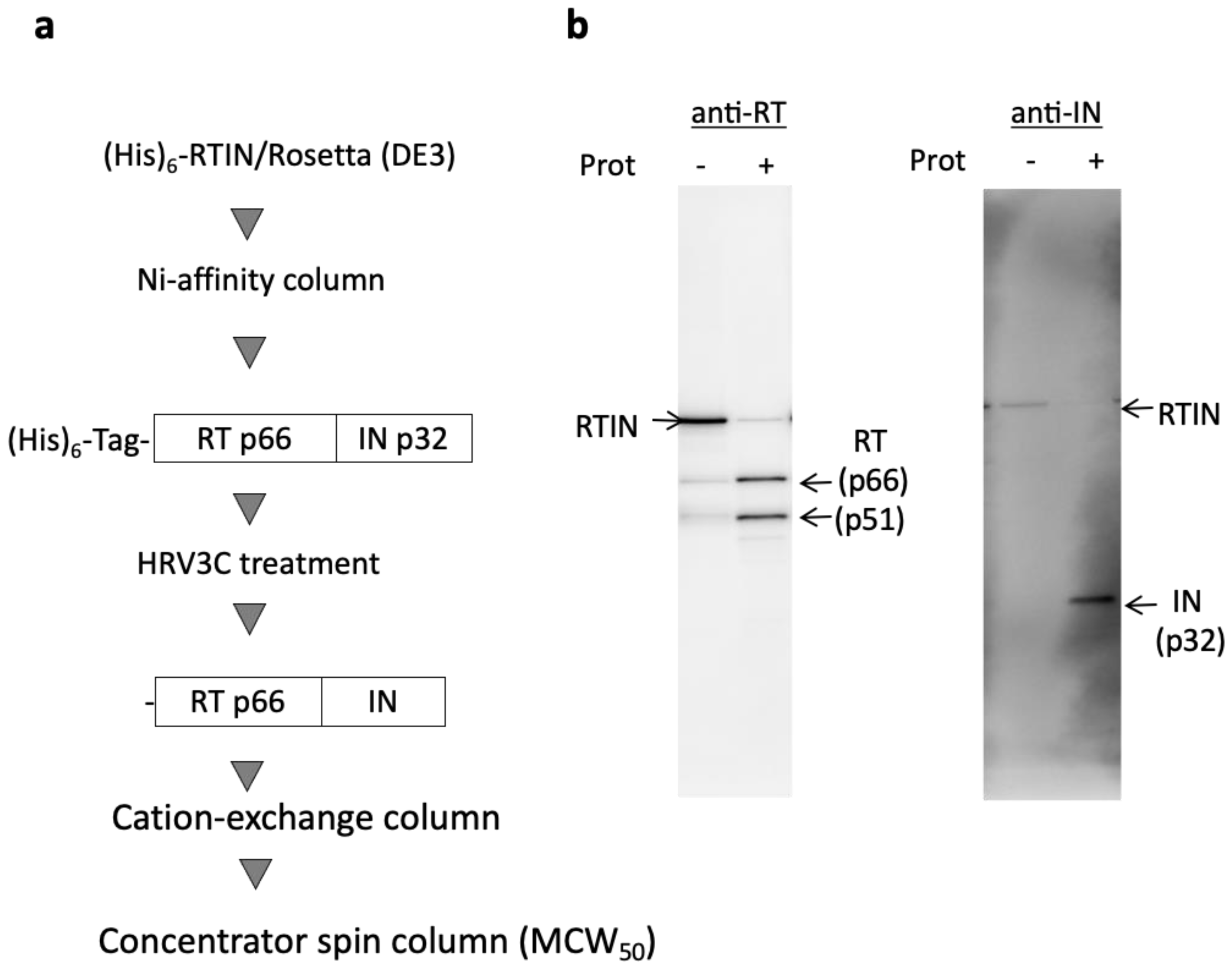
3.1. Preparation of Recombinant RTIN
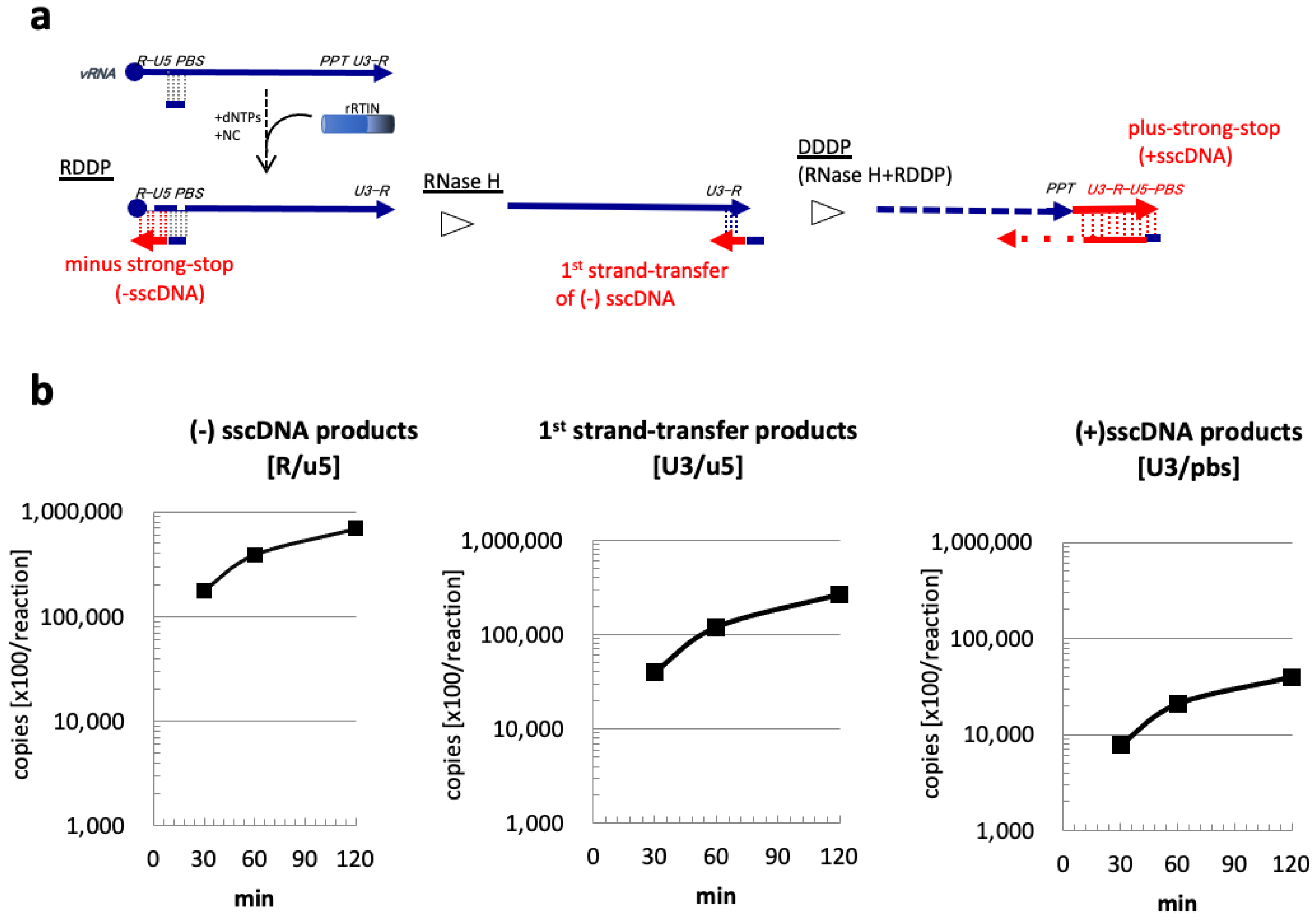
3.2. Evaluation of rRTIN Enzymatic Activities during Reverse Transcription
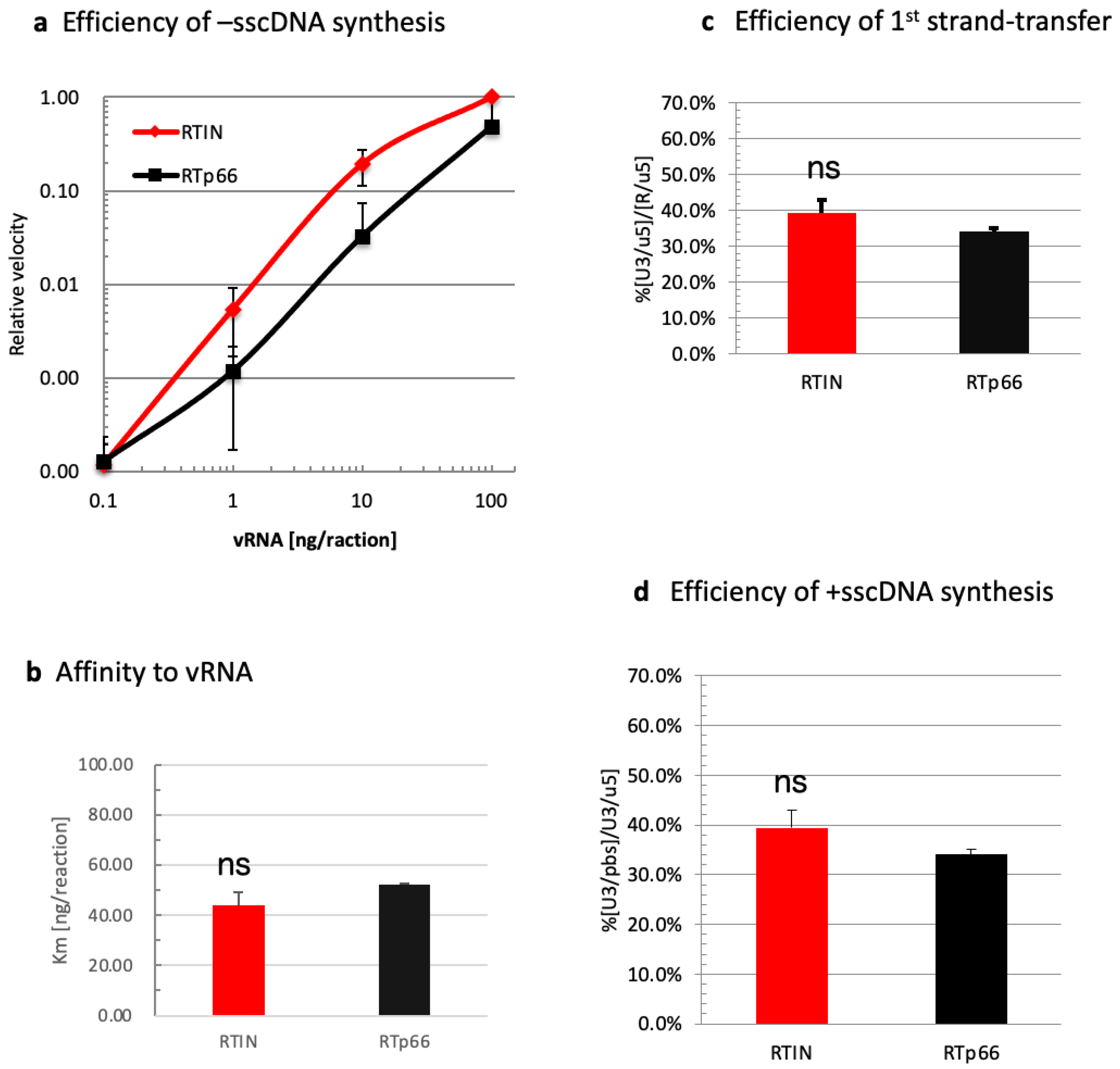
3.3. Impact of IN Fusion on RT Functions
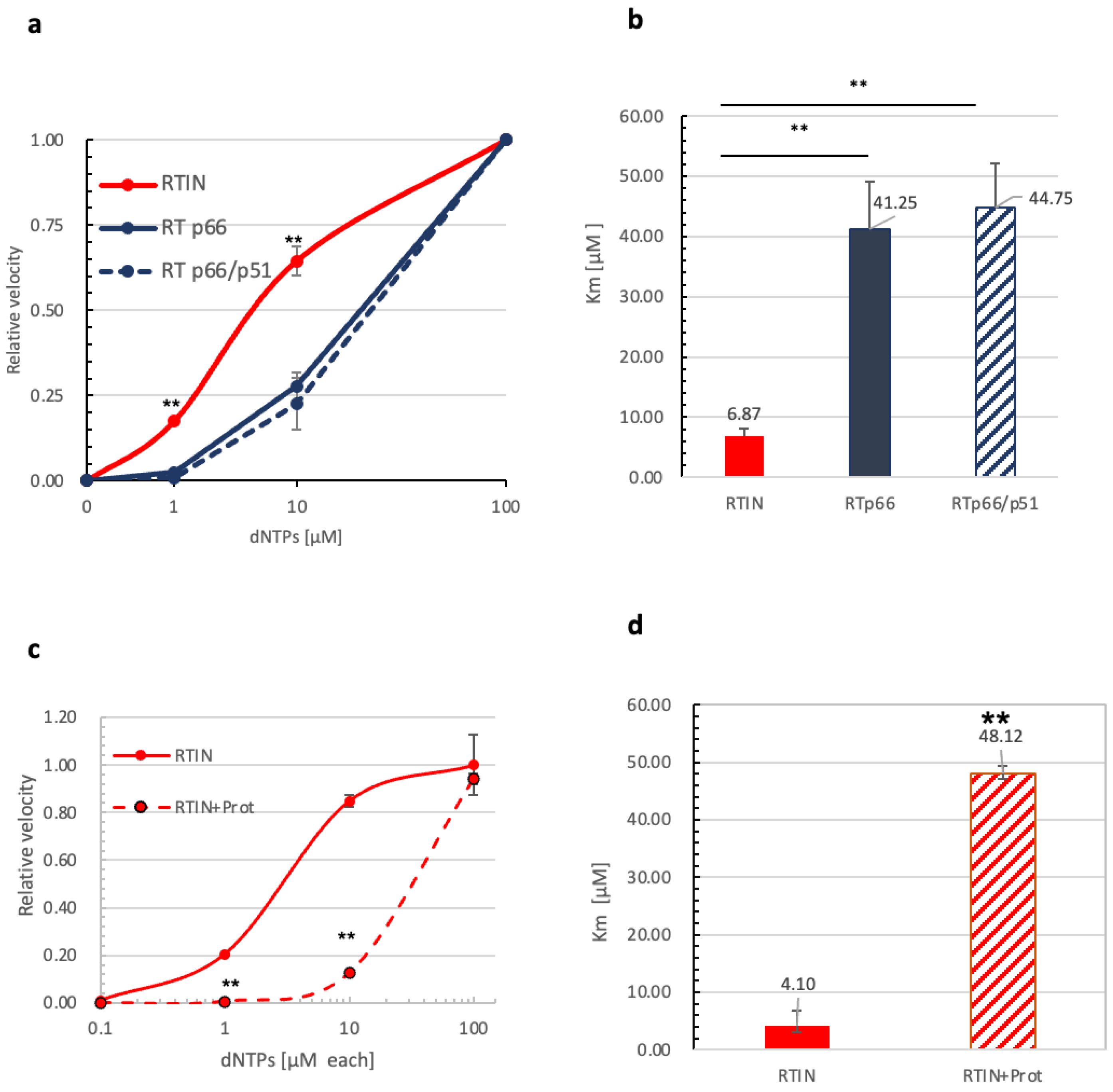
3.4. Impact of IN Fusion on Affinity of RT to dNTPs
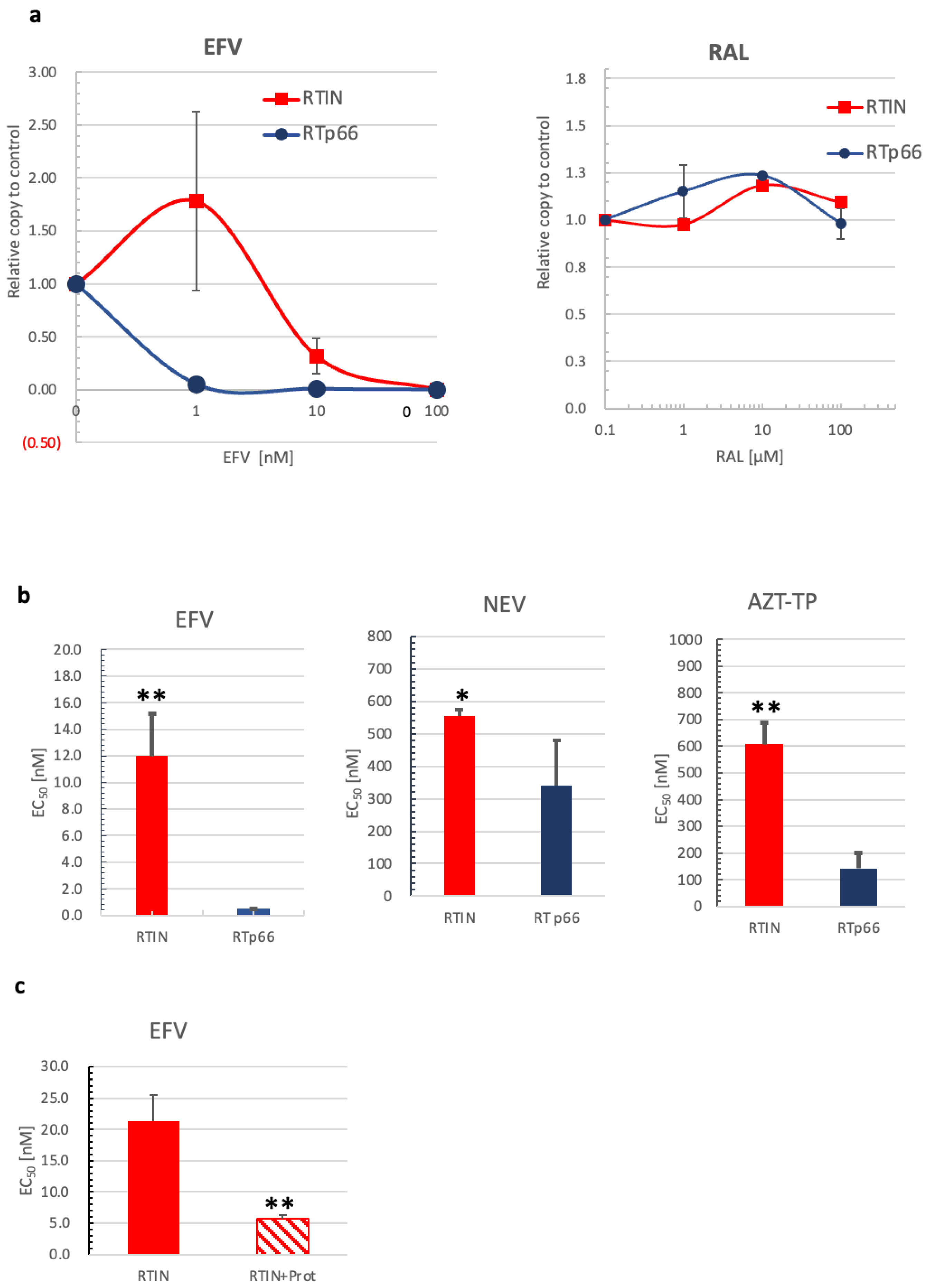
3.5. Effects of RTIN on RT and IN Inhibitors
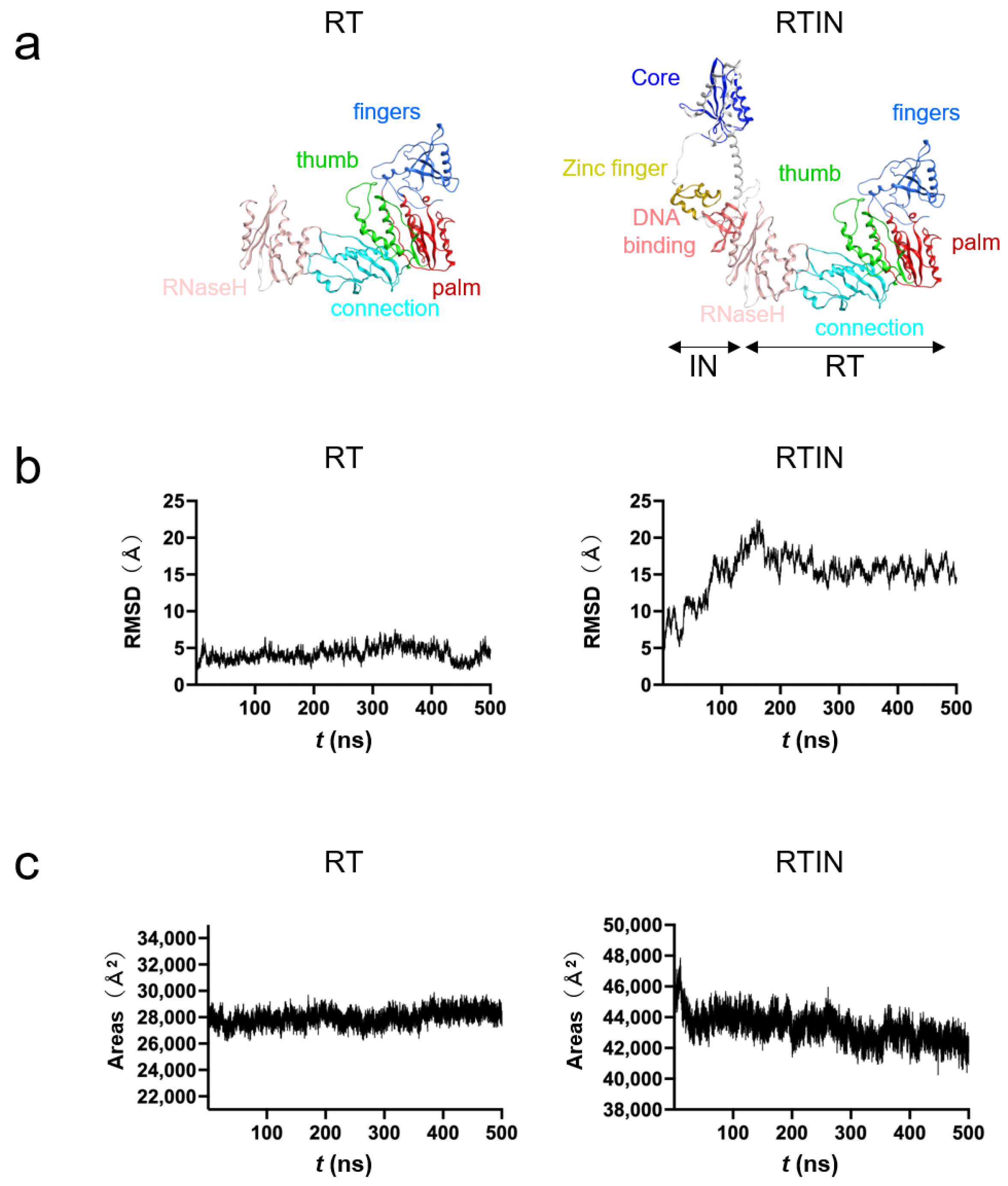
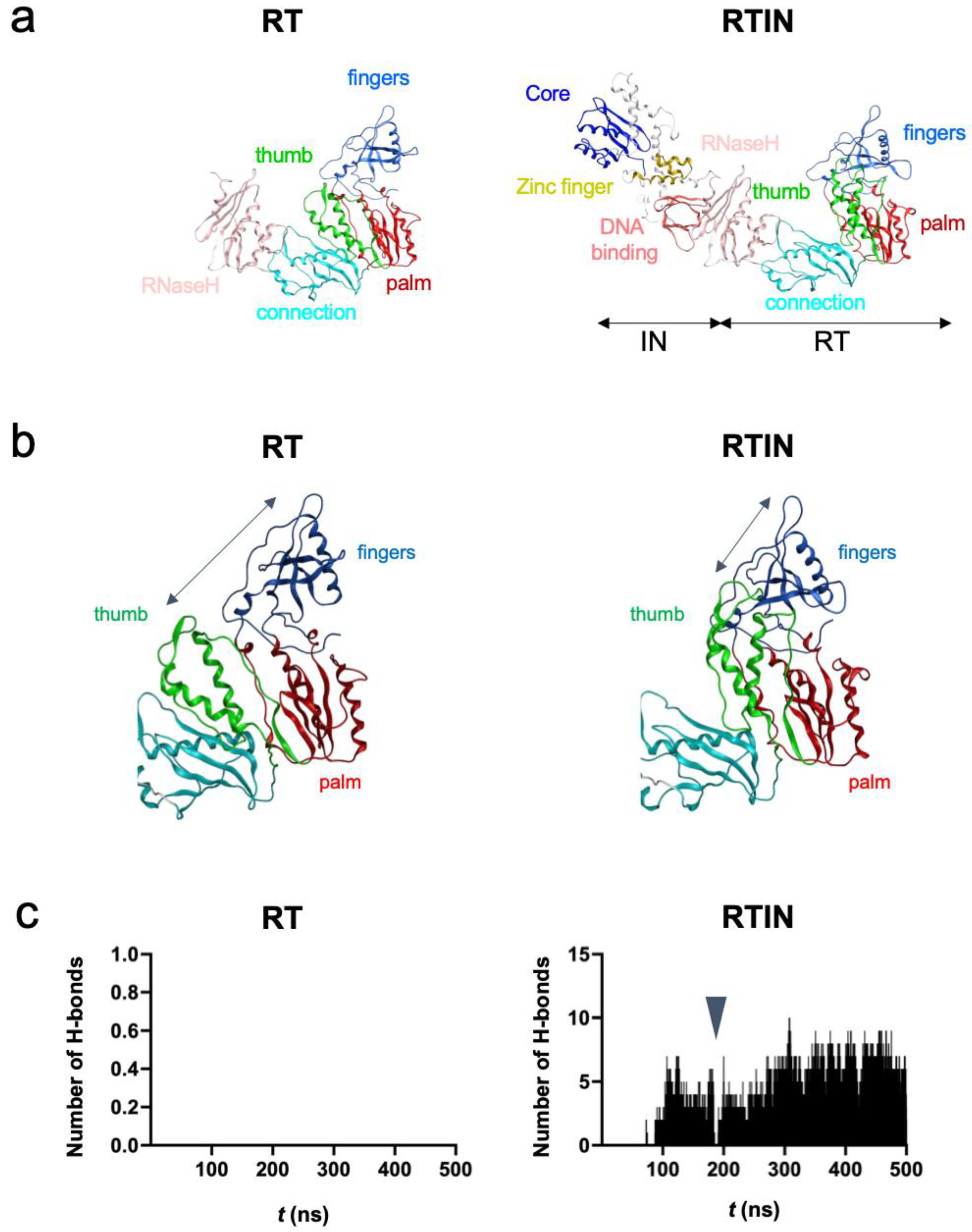
3.6. MD Simulations of RT and RTIN Proteins
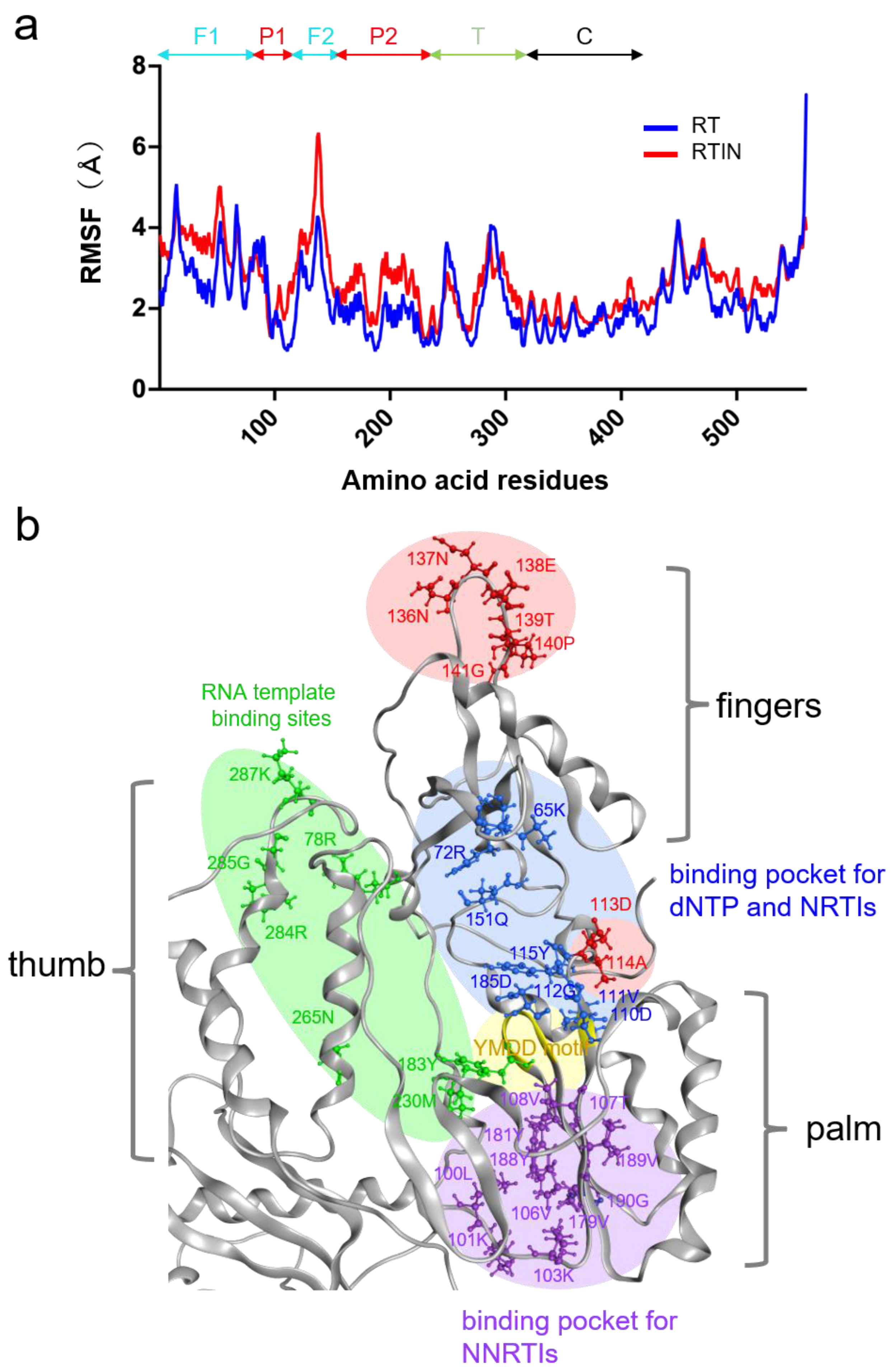
3.7. Structural Impacts of IN Fusion on RT Active Center
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hu, W.-S.; Hughes, S.H. HIV-1 Reverse Transcription. Cold Spring Harb. Perspect. Med. 2012, 2, a006882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, P.O. Integration. In Retroviruses; Coffin, J.M., Hughes, S.H., Varmus, H.E., Eds.; Cold Spring Harbor Laboratory Press: Long Island, NY, USA, 1997; pp. 161–203. [Google Scholar]
- Stevenson, M.; Stanwick, T.; Dempsey, M.; Lamonica, C. HIV-1 replication is controlled at the level of T cell activation and proviral integration. EMBO J. 1990, 9, 1551–1560. [Google Scholar] [CrossRef] [PubMed]
- Tsurutani, N.; Kubo, M.; Maeda, Y.; Ohashi, T.; Yamamoto, N.; Kannagi, M.; Masuda, T. Identification of Critical Amino Acid Residues in Human Immunodeficiency Virus Type 1 IN Required for Efficient Proviral DNA Formation at Steps prior to Integration in Dividing and Nondividing Cells. J. Virol. 2000, 74, 4795–4806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldstone, D.C.; Ennis-Adeniran, V.; Hedden, J.J.; Groom, H.C.T.; Rice, G.I.; Christodoulou, E.; Walker, P.A.; Kelly, G.; Haire, L.F.; Yap, M.W.; et al. HIV-1 restriction factor SAMHD1 is a deoxynucleoside triphosphate triphosphohydrolase. Nature 2011, 480, 379–382. [Google Scholar] [CrossRef]
- Schatz, O.; Mous, J.; Le Grice, S. HIV-1 RT-associated ribonuclease H displays both endonuclease and 3′-5′ exonuclease activity. EMBO J. 1990, 9, 1171–1176. [Google Scholar] [CrossRef]
- Nakamuraa, T.; Masuda, T.; Gotob, T.; Sanob, K.; Nakaib, M.; Haradaa, S. Lack of Infectivity of HIV-1 Integrase Zinc Finger-like Domain Mutant with Morphologically Normal Maturation. Biochem. Biophys. Res. Commun. 1997, 239, 715–722. [Google Scholar] [CrossRef]
- Masuda, T.; Planelles, V.; Krogstad, P.; Chen, I.S. Genetic analysis of human immunodeficiency virus type 1 integrase and the U3 att site: Unusual phenotype of mutants in the zinc finger-like domain. J. Virol. 1995, 69, 6687–6696. [Google Scholar] [CrossRef] [Green Version]
- Engelman, A.; Englund, G.; Orenstein, J.M.; Martin, M.A.; Craigie, R. Multiple effects of mutations in human immunodeficiency virus type 1 integrase on viral replication. J. Virol. 1995, 69, 2729–2736. [Google Scholar] [CrossRef] [Green Version]
- Briones, M.S.; Dobard, C.W.; Chow, S.A. Role of Human Immunodeficiency Virus Type 1 Integrase in Uncoating of the Viral Core. J. Virol. 2010, 84, 5181–5190. [Google Scholar] [CrossRef] [Green Version]
- Masuda, T. Non-Enzymatic Functions of Retroviral Integrase: The Next Target for Novel Anti-HIV Drug Development. Front. Microbiol. 2011, 2, 210. [Google Scholar] [CrossRef]
- Elliott, J.L.; Kutluay, S.B. Going beyond Integration: The Emerging Role of HIV-1 Integrase in Virion Morphogenesis. Viruses 2020, 12, 1005. [Google Scholar] [CrossRef] [PubMed]
- Desimmie, B.A.; Schrijvers, R.; Demeulemeester, J.; Borrenberghs, D.; Weydert, C.; Thys, W.; Vets, S.; Van Remoortel, B.; Hofkens, J.; De Rijck, J.; et al. LEDGINs inhibit late stage HIV-1 replication by modulating integrase multimerization in the virions. Retrovirology 2013, 10, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelman, A.; Kessl, J.J.; Kvaratskhelia, M. Allosteric inhibition of HIV-1 integrase activity. Curr. Opin. Chem. Biol. 2013, 17, 339–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kessl, J.J.; Kutluay, S.B.; Townsend, D.; Rebensburg, S.; Slaughter, A.; Larue, R.; Shkriabai, N.; Bakouche, N.; Fuchs, J.R.; Bieniasz, P.D.; et al. HIV-1 Integrase Binds the Viral RNA Genome and Is Essential during Virion Morphogenesis. Cell 2016, 166, 1257–1268.e12. [Google Scholar] [CrossRef] [Green Version]
- Elliott, J.L.; E Eschbach, J.; Koneru, P.C.; Li, W.; Puray-Chavez, M.; Townsend, D.; Lawson, D.Q.; Engelman, A.N.; Kvaratskhelia, M.; Kutluay, S.B. Integrase-RNA interactions underscore the critical role of integrase in HIV-1 virion morphogenesis. Elife 2020, 9, e54311. [Google Scholar] [CrossRef]
- Liu, S.; Koneru, P.C.; Li, W.; Pathirage, C.; Engelman, A.N.; Kvaratskhelia, M.; Musier-Forsyth, K. HIV-1 integrase binding to genomic RNA 5′-UTR induces local structural changes in vitro and in virio. Retrovirology 2021, 18, 1–16. [Google Scholar] [CrossRef]
- Swanstrom, R.; Wills, J.W. Synthesis, Assembly, and Processing of Viral Proteins. In Retroviruses; Coffin, J.M., Hughes, S.H., Varmus, H.E., Eds.; Cold Spring Harbor Laboratory Press: Long Island, NY, USA, 1997. [Google Scholar]
- Freed, E.O. HIV-1 assembly, release and maturation. Nat. Rev. Genet. 2015, 13, 484–496. [Google Scholar] [CrossRef]
- Takahata, T.; Takeda, E.; Tobiume, M.; Tokunaga, K.; Yokoyama, M.; Huang, Y.-L.; Hasegawa, A.; Shioda, T.; Sato, H.; Kannagi, M.; et al. Critical Contribution of Tyr15 in the HIV-1 Integrase (IN) in Facilitating IN Assembly and Nonenzymatic Function through the IN Precursor Form with Reverse Transcriptase. J. Virol. 2017, 91, e02003-16. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.-L.; Kawai, G.; Hasegawa, A.; Kannagi, M.; Masuda, T. Impact of 5′-end nucleotide modifications of HIV-1 genomic RNA on reverse transcription. Biochem. Biophys. Res. Commun. 2019, 516, 1145–1151. [Google Scholar] [CrossRef]
- Masuda, T.; Sato, Y.; Huang, Y.-L.; Koi, S.; Takahata, T.; Hasegawa, A.; Kawai, G.; Kannagi, M. Fate of HIV-1 cDNA intermediates during reverse transcription is dictated by transcription initiation site of virus genomic RNA. Sci. Rep. 2015, 5, 17680. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Adachi, A.; Gendelman, H.E.; Koenig, S.; Folks, T.; Willey, R.; Rabson, A.; Martin, M.A. Production of acquired immunodeficiency syndrome-associated retrovirus in human and nonhuman cells transfected with an infectious molecular clone. J. Virol. 1986, 59, 284–291. [Google Scholar] [CrossRef] [Green Version]
- Smerdon, S.J.; Jäger, J.; Wang, J.; Kohlstaedt, L.A.; Chirino, A.J.; Friedman, J.M.; Rice, P.A.; Steitz, T.A. Structure of the binding site for nonnucleoside inhibitors of the reverse transcriptase of human immunodeficiency virus type 1. Proc. Natl. Acad. Sci. USA 1994, 91, 3911–3915. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Ling, H.; Yang, W.; Craigie, R. Structure of a two-domain fragment of HIV-1 integrase: Implications for domain organization in the intact protein. EMBO J. 2001, 20, 7333–7343. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.C.-H.; Krucinski, J.; Miercke, L.J.W.; Finer-Moore, J.S.; Tang, A.H.; Leavitt, A.D.; Stroud, R.M. Crystal structure of the HIV-1 integrase catalytic core and C-terminal domains: A model for viral DNA binding. Proc. Natl. Acad. Sci. USA 2000, 97, 8233–8238. [Google Scholar] [CrossRef] [Green Version]
- Koma, T.; Kotani, O.; Miyakawa, K.; Ryo, A.; Yokoyama, M.; Doi, N.; Adachi, A.; Sato, H.; Nomaguchi, M. Allosteric Regulation of HIV-1 Capsid Structure for Gag Assembly, Virion Production, and Viral Infectivity by a Disordered Interdomain Linker. J. Virol. 2019, 93, e00381-19. [Google Scholar] [CrossRef] [Green Version]
- Izumida, M.; Kotani, O.; Hayashi, H.; Smith, C.; Fukuda, T.; Suga, K.; Iwao, M.; Ishibashi, F.; Sato, H.; Kubo, Y. Unique Mode of Antiviral Action of a Marine Alkaloid against Ebola Virus and SARS-CoV-2. Viruses 2022, 14, 816. [Google Scholar] [CrossRef]
- Koma, T.; Yokoyama, M.; Kotani, O.; Doi, N.; Nakanishi, N.; Okubo, H.; Adachi, S.; Adachi, A.; Sato, H.; Nomaguchi, M. Species-Specific Valid Ternary Interactions of HIV-1 Env-gp120, CD4, and CCR5 as Revealed by an Adaptive Single-Amino Acid Substitution at the V3 Loop Tip. J. Virol. 2021, 95, e02177-20. [Google Scholar] [CrossRef]
- Kotani, O.; Suzuki, Y.; Saito, S.; Ainai, A.; Ueno, A.; Hemmi, T.; Sano, K.; Tabata, K.; Yokoyama, M.; Suzuki, T.; et al. Structure-Guided Creation of an Anti-HA Stalk Antibody F11 Derivative That Neutralizes Both F11-Sensitive and -Resistant Influenza A(H1N1)pdm09 Viruses. Viruses 2021, 13, 1733. [Google Scholar] [CrossRef]
- Case, D.A.; Betz, R.M.; Cerutti, D.S.; Cheatham, T.E., III; Darden, T.A.; Duke, R.E.; Giese, T.J.; Gohlke, H.; Goetz, A.W.; Homeyer, N.; et al. AMBER 16; University of California: San Francisco, CA, USA, 2016. [Google Scholar]
- Maier, J.A.; Martinez, C.; Kasavajhala, K.; Wickstrom, L.; Hauser, K.E.; Simmerling, C. ff14SB: Improving the accuracy of protein side chain and backbone parameters from ff99SB. J. Chem. Theory Comput. 2015, 11, 3696–3713. [Google Scholar] [CrossRef]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Ryckaert, J.-P.; Ciccotti, G.; Berendsen, H.J.C. Numerical integration of the cartesian equations of motion of a system with constraints: Molecular dynamics of n-alkanes. J. Comput. Phys. 1977, 23, 327–341. [Google Scholar] [CrossRef] [Green Version]
- Weiser, J.; Shenkin, P.S.; Still, W.C. Approximate atomic surfaces from linear combinations of pairwise overlaps (LCPO). J. Comput. Chem. 1999, 20, 217–230. [Google Scholar] [CrossRef]
- Krishnan, L.; Li, X.; Naraharisetty, H.L.; Hare, S.; Cherepanov, P.; Engelman, A. Structure-based modeling of the functional HIV-1 intasome and its inhibition. Proc. Natl. Acad. Sci. USA 2010, 107, 15910–15915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollingsworth, S.A.; Dror, R.O. Molecular Dynamics Simulation for All. Neuron 2018, 99, 1129–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dodson, G.G.; Lane, D.; Verma, C.S. Molecular simulations of protein dynamics: New windows on mechanisms in biology. EMBO Rep. 2008, 9, 144–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karplus, M.; Kuriyan, J. Molecular dynamics and protein function. Proc. Natl. Acad. Sci. USA 2005, 102, 6679–6685. [Google Scholar] [CrossRef] [Green Version]
- Keul, N.D.; Oruganty, K.; Schaper Bergman, E.T.; Beattie, N.R.; McDonald, W.E.; Kadirvelraj, R.; Gross, M.L.; Phillips, R.S.; Harvey, S.C.; Wood, Z.A. The entropic force generated by intrinsically disordered segments tunes protein function. Nature 2018, 563, 584–588. [Google Scholar] [CrossRef]
- Kuwata, T.; Takaki, K.; Yoshimura, K.; Enomoto, I.; Wu, F.; Ourmanov, I.; Hirsch, V.M.; Yokoyama, M.; Sato, H.; Matsushita, S. Conformational Epitope Consisting of the V3 and V4 Loops as a Target for Potent and Broad Neutralization of Simian Immunodeficiency Viruses. J. Virol. 2013, 87, 5424–5436. [Google Scholar] [CrossRef] [Green Version]
- Sakuragi, S.; Kotani, O.; Yokoyama, M.; Shioda, T.; Sato, H.; Sakuragi, J.-I. Identification of a Novel Cis-Acting Regulator of HIV-1 Genome Packaging. Int. J. Mol. Sci. 2021, 22, 3435. [Google Scholar] [CrossRef]
- Eyokoyama, M.; Enomaguchi, M.; Edoi, N.; Ekanda, T.; Eadachi, A.; Esato, H. In silico Analysis of HIV-1 Env-gp120 Reveals Structural Bases for Viral Adaptation in Growth-Restrictive Cells. Front. Microbiol. 2016, 7, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarafianos, S.G.; Marchand, B.; Das, K.; Himmel, D.M.; Parniak, M.A.; Hughes, S.H.; Arnold, E. Structure and Function of HIV-1 Reverse Transcriptase: Molecular Mechanisms of Polymerization and Inhibition. J. Mol. Biol. 2009, 385, 693–713. [Google Scholar] [CrossRef] [Green Version]
- Das, K.; Martinez, S.E.; Bauman, J.D.; Arnold, E. HIV-1 reverse transcriptase complex with DNA and nevirapine reveals non-nucleoside inhibition mechanism. Nat. Struct. Mol. Biol. 2012, 19, 253–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nissley, D.V.; Radzio, J.; Ambrose, Z.; Sheen, C.-W.; Hamamouch, N.; Moore, K.L.; Tachedjian, G.; Sluis-Cremer, N. Characterization of novel non-nucleoside reverse transcriptase (RT) inhibitor resistance mutations at residues 132 and 135 in the 51 kDa subunit of HIV-1 RT. Biochem. J. 2007, 404, 151–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyer, P.L.; Gao, H.-Q.; Clark, P.K.; Sarafianos, S.G.; Arnold, E.; Hughes, S.H. YADD Mutants of Human Immunodeficiency Virus Type 1 and Moloney Murine Leukemia Virus Reverse Transcriptase Are Resistant to Lamivudine Triphosphate (3TCTP) In Vitro. J. Virol. 2001, 75, 6321–6328. [Google Scholar] [CrossRef] [Green Version]
- Dobard, C.W.; Briones, M.S.; Chow, S.A. Molecular Mechanisms by Which Human Immunodeficiency Virus Type 1 Integrase Stimulates the Early Steps of Reverse Transcription. J. Virol. 2007, 81, 10037–10046. [Google Scholar] [CrossRef] [Green Version]
- Zhu, K.; Dobard, C.; Chow, S.A. Requirement for Integrase during Reverse Transcription of Human Immunodeficiency Virus Type 1 and the Effect of Cysteine Mutations of Integrase on Its Interactions with Reverse Transcriptase. J. Virol. 2004, 78, 5045–5055. [Google Scholar] [CrossRef] [Green Version]
- Cohen, A.; Barankiewicz, J.; Lederman, H.M.; Gelfand, E.W. Purine and pyrimidine metabolism in human T lymphocytes. Regulation of deoxyribonucleotide metabolism. J. Biol. Chem. 1983, 258, 12334–12340. [Google Scholar] [CrossRef]
- Traut, T.W. Physiological concentrations of purines and pyrimidines. Mol. Cell. Biochem. 1994, 140, 1–22. [Google Scholar] [CrossRef]
- Bai, H.; Kath, J.E.; Zörgiebel, F.M.; Sun, M.; Ghosh, P.; Hatfull, G.F.; Grindley, N.D.F.; Marko, J.F. Remote control of DNA-acting enzymes by varying the Brownian dynamics of a distant DNA end. Proc. Natl. Acad. Sci. USA 2012, 109, 16546–16551. [Google Scholar] [CrossRef]
- Sano, K.; Saito, S.; Suzuki, T.; Kotani, O.; Ainai, A.; van Riet, E.; Tabata, K.; Saito, K.; Takahashi, Y.; Yokoyama, M.; et al. An influenza HA stalk reactive polymeric IgA antibody exhibits anti-viral function regulated by binary interaction between HA and the antibody. PLoS ONE 2021, 16, e0245244. [Google Scholar] [CrossRef] [PubMed]
- Herschhorn, A.; Oz-Gleenberg, I.; Hizi, A. Quantitative analysis of the interactions between HIV-1 integrase and retroviral reverse transcriptases. Biochem. J. 2008, 412, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Gleenberg, I.O.; Goldgur, Y.; Hizi, A. Ile178 of HIV-1 reverse transcriptase is critical for inhibiting the viral integrase. Biochem. Biophys. Res. Commun. 2007, 364, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Grandgenett, D.P.; Gerard, G.F.; Green, M. A Single Subunit from Avian Myeloblastosis Virus with Both RNA-Directed DNA Polymerase and Ribonuclease H Activity. Proc. Natl. Acad. Sci. USA 1973, 70, 230–234. [Google Scholar] [CrossRef] [Green Version]
- Engelman, A.N.; Kvaratskhelia, M. Multimodal Functionalities of HIV-1 Integrase. Viruses 2022, 14, 926. [Google Scholar] [CrossRef]
- Rankovic, S.; Deshpande, A.; Harel, S.; Aiken, C.; Rousso, I. HIV-1 Uncoating Occurs via a Series of Rapid Biomechanical Changes in the Core Related to Individual Stages of Reverse Transcription. J. Virol. 2021, 95, e00166-21. [Google Scholar] [CrossRef]
- Dharan, A.; Bachmann, N.; Talley, S.; Zwikelmaier, V.; Campbell, E.M. Nuclear pore blockade reveals that HIV-1 completes reverse transcription and uncoating in the nucleus. Nat. Microbiol. 2020, 5, 1088–1095. [Google Scholar] [CrossRef]
- Burdick, R.C.; Li, C.; Munshi, M.; Rawson, J.M.O.; Nagashima, K.; Hu, W.-S.; Pathak, V.K. HIV-1 uncoats in the nucleus near sites of integration. Proc. Natl. Acad. Sci. USA 2020, 117, 5486–5493. [Google Scholar] [CrossRef]
- Arhel, N.J. Early HIV replication revisited. Nat. Microbiol. 2020, 5, 1065–1066. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masuda, T.; Kotani, O.; Yokoyama, M.; Abe, Y.; Kawai, G.; Sato, H. Cis-Allosteric Regulation of HIV-1 Reverse Transcriptase by Integrase. Viruses 2023, 15, 31. https://doi.org/10.3390/v15010031
Masuda T, Kotani O, Yokoyama M, Abe Y, Kawai G, Sato H. Cis-Allosteric Regulation of HIV-1 Reverse Transcriptase by Integrase. Viruses. 2023; 15(1):31. https://doi.org/10.3390/v15010031
Chicago/Turabian StyleMasuda, Takao, Osamu Kotani, Masaru Yokoyama, Yuya Abe, Gota Kawai, and Hironori Sato. 2023. "Cis-Allosteric Regulation of HIV-1 Reverse Transcriptase by Integrase" Viruses 15, no. 1: 31. https://doi.org/10.3390/v15010031