
Microarray-Based Analyses of Rhinovirus Species-Specific Antibody Responses in Exacerbated Pediatric Asthma in a German Pediatric Cohort

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient Characteristics and Biomaterial Collection
2.2. Microarray-Based Determination of Virus-Specific IgG Levels
2.3. Statistical Analyses
3. Results
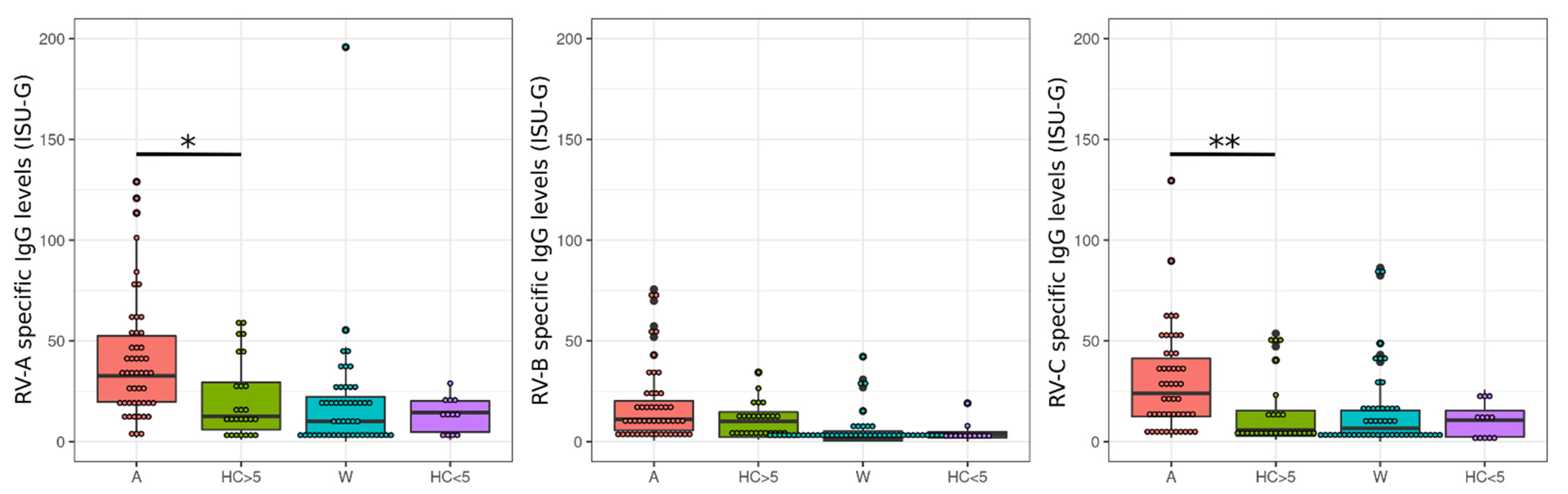
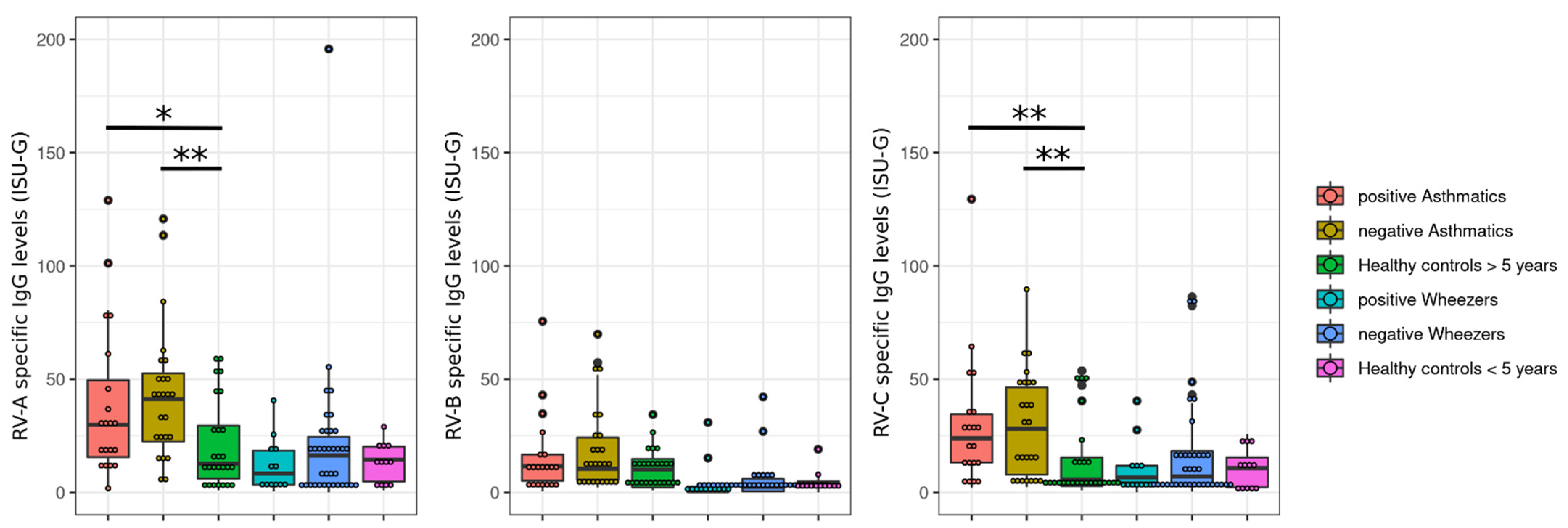
3.1. Rhinovirus Antibody Levels Are Increased in Exacerbated Asthmatics
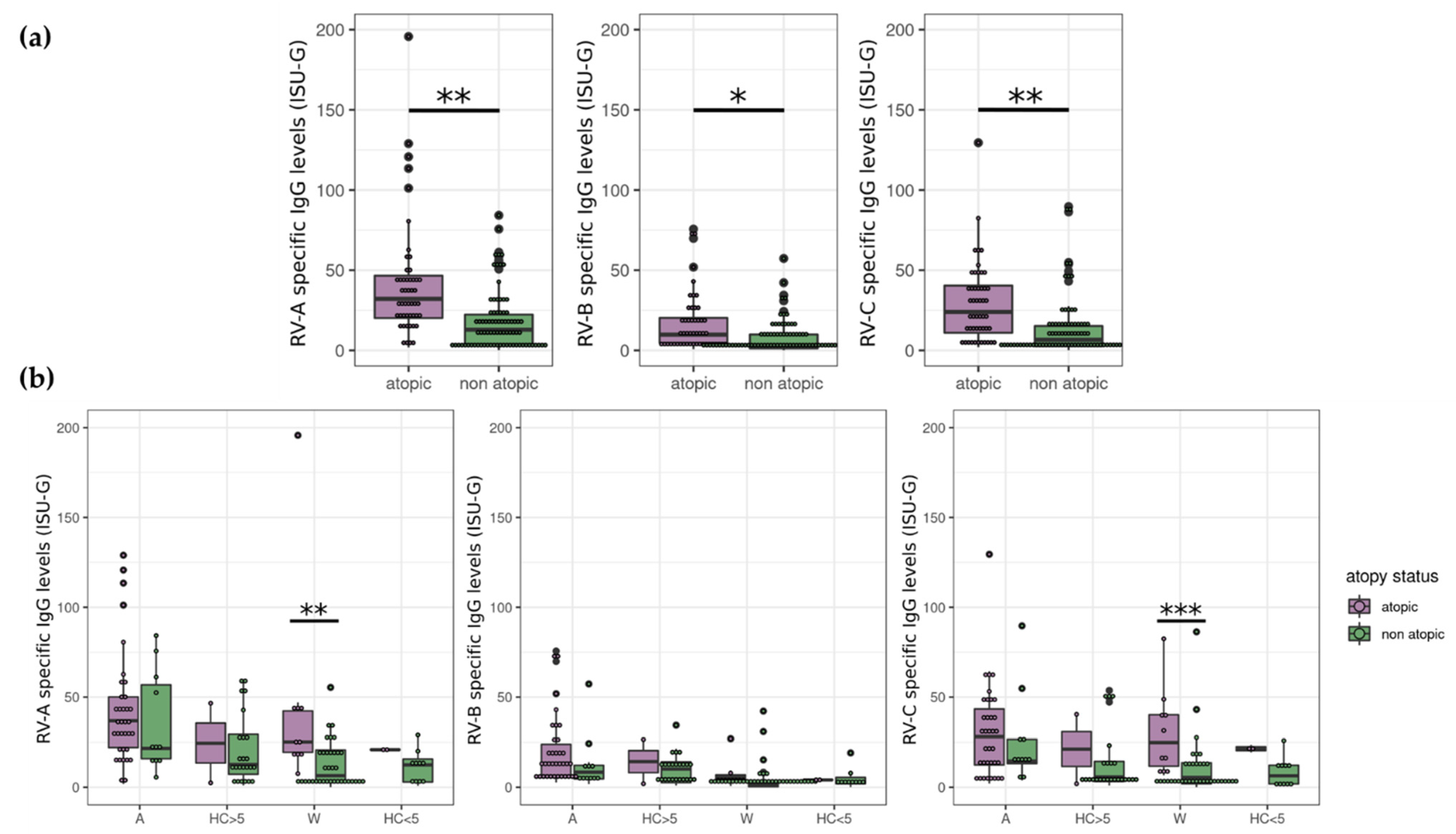
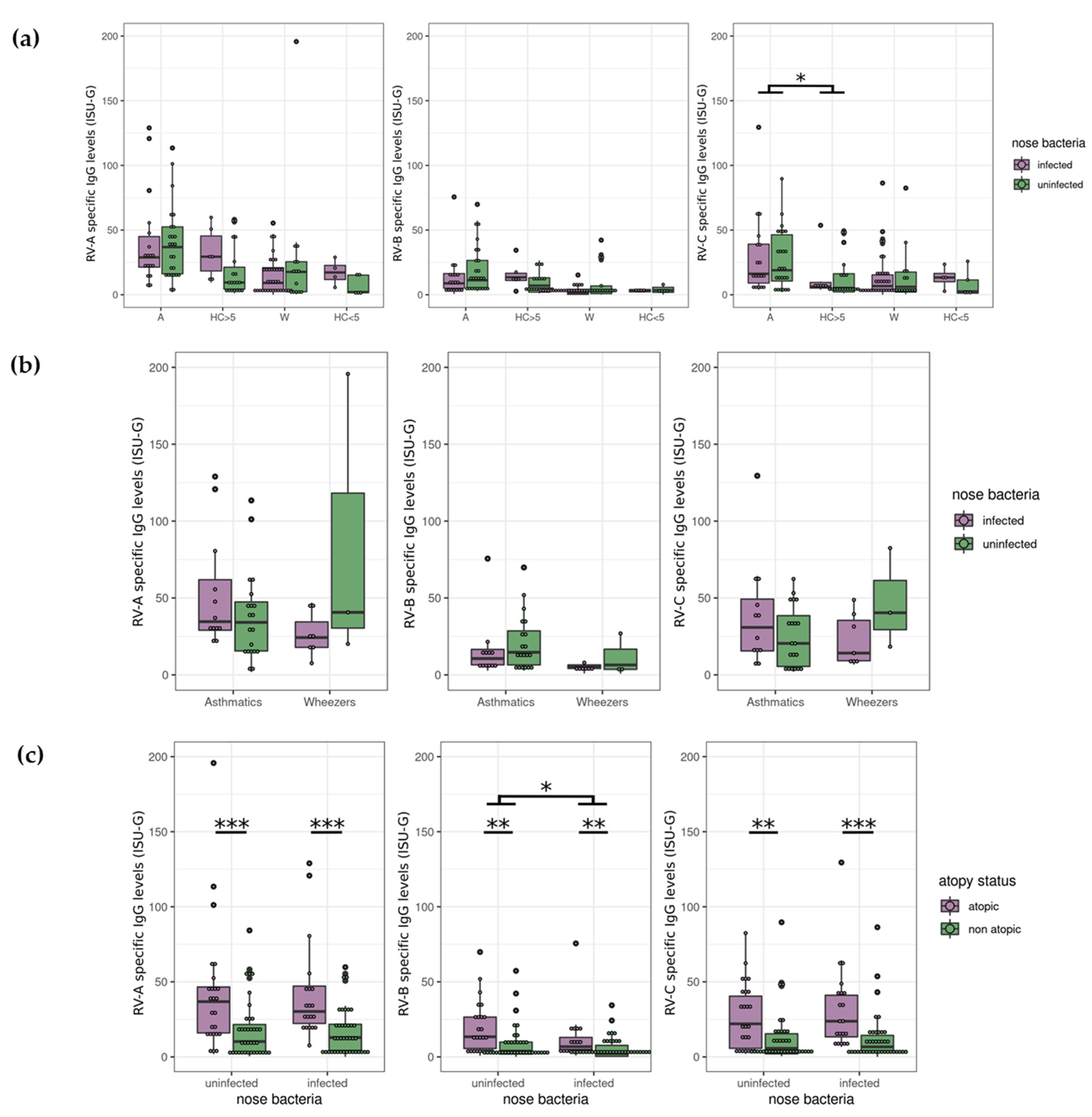
3.2. Atopy Strongly Influences RV Antibody Levels
3.3. The Steroid Status of Exacerbated Patients Does Not Significantly Correlate with RV-Specific IgG Levels
3.4. Nasopharyngeal Test Results Revealed no Correlation between Bacteria and RV
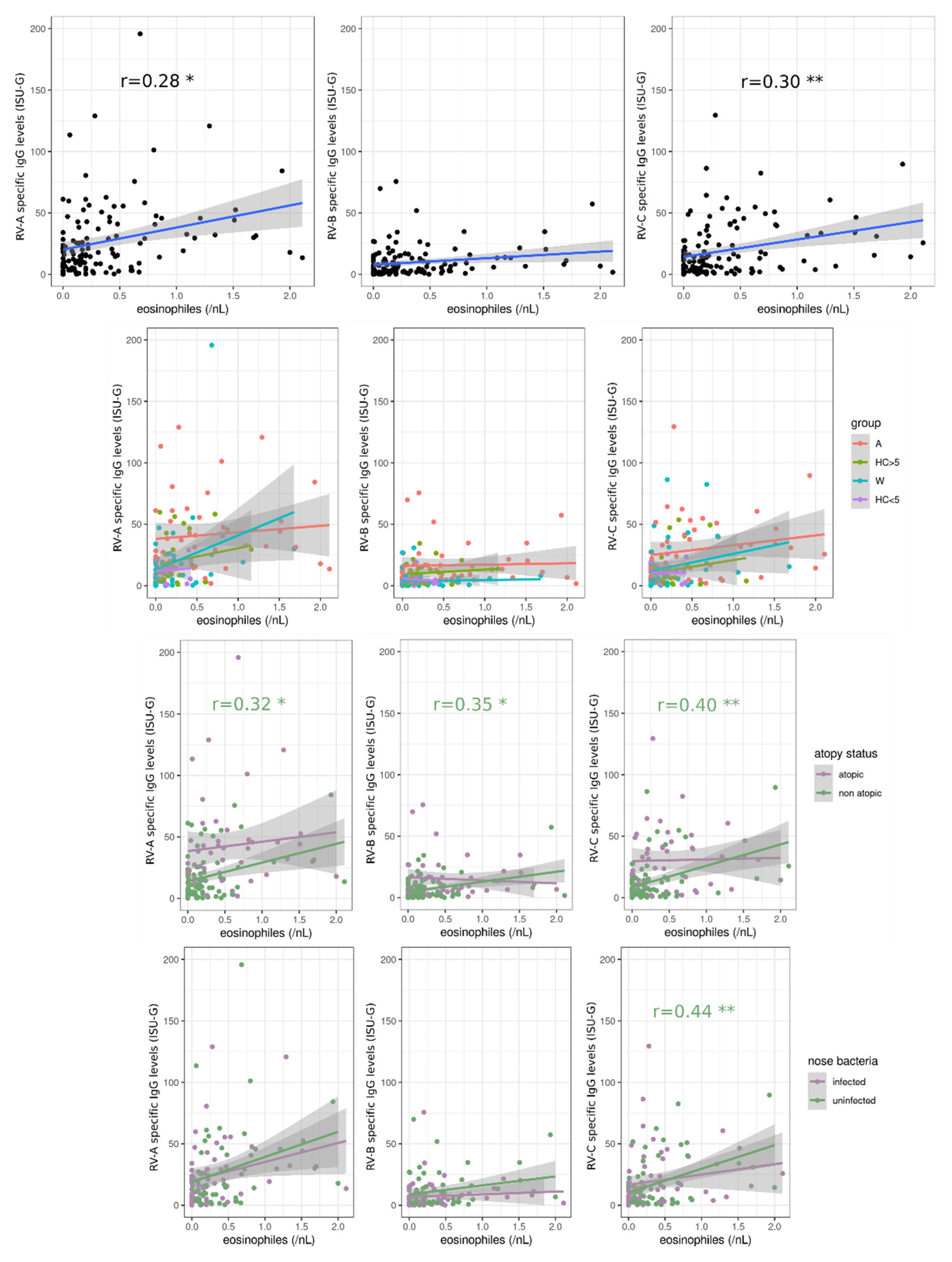
3.5. Blood Eosinophils Are Positively Correlated with RV-Specific IgG Levels
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hopkin, J.M. The diagnosis of asthma, a clinical syndrome. Thorax 2012, 67, 660–662. [Google Scholar] [CrossRef] [Green Version]
- Rothe, T.; Spagnolo, P.; Bridevaux, P.O.; Clarenbach, C.; Eich-Wanger, C.; Meyer, F.; Miedinger, D.; Moller, A.; Nicod, L.P.; Nicolet-Chatelain, G.; et al. Diagnosis and Management of Asthma-The Swiss Guidelines. Respiration 2018, 95, 364–380. [Google Scholar] [CrossRef]
- Wenzel, S.E. Complex phenotypes in asthma: Current definitions. Pulm. Pharmacol. Ther. 2013, 26, 710–715. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.F.; Wenzel, S.E.; Brozek, J.L.; Bush, A.; Castro, M.; Sterk, P.J.; Adcock, I.M.; Bateman, E.D.; Bel, E.H.; Bleecker, E.R.; et al. International ERS/ATS guidelines on definition, evaluation and treatment of severe asthma. Eur. Respir. J. 2014, 43, 343–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heikkinen, T.; Jarvinen, A. The common cold. Lancet 2003, 361, 51–59. [Google Scholar] [CrossRef]
- Landes, M.B.; Neil, R.B.; McCool, S.S.; Mason, B.P.; Woron, A.M.; Garman, R.L.; Smalley, D.L. The frequency and seasonality of influenza and other respiratory viruses in Tennessee: Two influenza seasons of surveillance data, 2010–2012. Influenza Other Respir. Viruses 2013, 7, 1122–1127. [Google Scholar] [CrossRef] [Green Version]
- Midgley, C.M.; Haynes, A.K.; Baumgardner, J.L.; Chommanard, C.; Demas, S.W.; Prill, M.M.; Abedi, G.R.; Curns, A.T.; Watson, J.T.; Gerber, S.I. Determining the Seasonality of Respiratory Syncytial Virus in the United States: The Impact of Increased Molecular Testing. J. Infect. Dis. 2017, 216, 345–355. [Google Scholar] [CrossRef]
- Morikawa, S.; Kohdera, U.; Hosaka, T.; Ishii, K.; Akagawa, S.; Hiroi, S.; Kase, T. Seasonal variations of respiratory viruses and etiology of human rhinovirus infection in children. J. Clin. Virol. 2015, 73, 14–19. [Google Scholar] [CrossRef]
- Moriyama, M.; Hugentobler, W.J.; Iwasaki, A. Seasonality of Respiratory Viral Infections. Annu. Rev. Virol. 2020, 7, 83–101. [Google Scholar] [CrossRef]
- Busse, W.W.; Lemanske, R.F., Jr.; Gern, J.E. Role of viral respiratory infections in asthma and asthma exacerbations. Lancet 2010, 376, 826–834. [Google Scholar] [CrossRef]
- Johnston, S.L.; Pattemore, P.K.; Sanderson, G.; Smith, S.; Lampe, F.; Josephs, L.; Symington, P.; O’Toole, S.; Myint, S.H.; Tyrrell, D.A.; et al. Community study of role of viral infections in exacerbations of asthma in 9-11 year old children. BMJ 1995, 310, 1225–1229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikhail, I.; Grayson, M.H. Asthma and viral infections: An intricate relationship. Ann. Allergy Asthma Immunol. 2019, 123, 352–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, S.Y.; Wang, L.L.; Ren, L.; Luo, J.; Liao, W.; Liu, E.M. Epidemiological analysis and follow-up of human rhinovirus infection in children with asthma exacerbation. J. Med. Virol. 2018, 90, 219–228. [Google Scholar] [CrossRef]
- Palmenberg, A.C.; Gern, J.E. Classification and evolution of human rhinoviruses. Methods Mol. Biol. 2015, 1221, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Zlateva, K.T.; de Vries, J.J.; Coenjaerts, F.E.; van Loon, A.M.; Verheij, T.; Little, P.; Butler, C.C.; Goossens, H.; Ieven, M.; Claas, E.C.; et al. Prolonged shedding of rhinovirus and re-infection in adults with respiratory tract illness. Eur. Respir. J. 2014, 44, 169–177. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, S.E.; Lamson, D.M.; St George, K.; Walsh, T.J. Human rhinoviruses. Clin. Microbiol. Rev. 2013, 26, 135–162. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, J.L.; Turner, R.B.; Braciale, T.; Heymann, P.W.; Borish, L. Pathogenesis of rhinovirus infection. Curr. Opin. Virol. 2012, 2, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Basnet, S.; Palmenberg, A.C.; Gern, J.E. Rhinoviruses and Their Receptors. Chest 2019, 155, 1018–1025. [Google Scholar] [CrossRef]
- Lee, W.M.; Lemanske, R.F., Jr.; Evans, M.D.; Vang, F.; Pappas, T.; Gangnon, R.; Jackson, D.J.; Gern, J.E. Human rhinovirus species and season of infection determine illness severity. Am. J. Respir. Crit. Care Med. 2012, 186, 886–891. [Google Scholar] [CrossRef] [Green Version]
- Michi, A.N.; Love, M.E.; Proud, D. Rhinovirus-Induced Modulation of Epithelial Phenotype: Role in Asthma. Viruses 2020, 12, 1328. [Google Scholar] [CrossRef]
- Marguet, C.; Lubrano, M.; Gueudin, M.; Le Roux, P.; Deschildre, A.; Forget, C.; Couderc, L.; Siret, D.; Donnou, M.D.; Bubenheim, M.; et al. In very young infants severity of acute bronchiolitis depends on carried viruses. PLoS ONE 2009, 4, e4596. [Google Scholar] [CrossRef] [PubMed]
- Ruuskanen, O.; Lahti, E.; Jennings, L.C.; Murdoch, D.R. Viral pneumonia. Lancet 2011, 377, 1264–1275. [Google Scholar] [CrossRef]
- Vandini, S.; Biagi, C.; Fischer, M.; Lanari, M. Impact of Rhinovirus Infections in Children. Viruses 2019, 11, 521. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, K.; Mansbach, J.M.; Bochkov, Y.A.; Gern, J.E.; Piedra, P.A.; Bauer, C.S.; Teach, S.J.; Wu, S.; Sullivan, A.F.; Camargo, C.A., Jr. Association of Rhinovirus C Bronchiolitis and Immunoglobulin E Sensitization during Infancy with Development of Recurrent Wheeze. JAMA Pediatr. 2019, 173, 544–552. [Google Scholar] [CrossRef]
- Lambert, K.A.; Prendergast, L.A.; Dharmage, S.C.; Tang, M.; O’Sullivan, M.; Tran, T.; Druce, J.; Bardin, P.; Abramson, M.J.; Erbas, B. The role of human rhinovirus (HRV) species on asthma exacerbation severity in children and adolescents. J. Asthma 2018, 55, 596–602. [Google Scholar] [CrossRef]
- Linsuwanon, P.; Payungporn, S.; Samransamruajkit, R.; Posuwan, N.; Makkoch, J.; Theanboonlers, A.; Poovorawan, Y. High prevalence of human rhinovirus C infection in Thai children with acute lower respiratory tract disease. J. Infect. 2009, 59, 115–121. [Google Scholar] [CrossRef]
- Niespodziana, K.; Stenberg-Hammar, K.; Papadopoulos, N.G.; Focke-Tejkl, M.; Errhalt, P.; Konradsen, J.R.; Soderhall, C.; van Hage, M.; Hedlin, G.; Valenta, R. Microarray Technology May Reveal the Contribution of Allergen Exposure and Rhinovirus Infections as Possible Triggers for Acute Wheezing Attacks in Preschool Children. Viruses 2021, 13, 915. [Google Scholar] [CrossRef]
- Lauinger, I.L.; Bible, J.M.; Halligan, E.P.; Bangalore, H.; Tosas, O.; Aarons, E.J.; MacMahon, E.; Tong, C.Y. Patient characteristics and severity of human rhinovirus infections in children. J. Clin. Virol. 2013, 58, 216–220. [Google Scholar] [CrossRef]
- Aydin, M.; Naumova, E.A.; Lutz, S.; Meyer-Bahlburg, A.; Arnold, W.H.; Kreppel, F.; Ehrhardt, A.; Postberg, J.; Wirth, S. Do Current Asthma-Preventive Measures Appropriately Face the World Health Organization’s Concerns: A Study Presentation of a New Clinical, Prospective, Multicentric Pediatric Asthma Exacerbation Cohort in Germany. Front. Pediatr. 2020, 8, 574462. [Google Scholar] [CrossRef]
- Aydin, M.; Naumova, E.A.; Paulsen, F.; Zhang, W.; Gopon, F.; Theis, C.; Lutz, S.; Ehrke-Schulz, E.; Arnold, W.H.; Wirth, S.; et al. House Dust Mite Exposure Causes Increased Susceptibility of Nasal Epithelial Cells to Adenovirus Infection. Viruses 2020, 12, 1151. [Google Scholar] [CrossRef]
- Aydin, M.; Weisser, C.; Rue, O.; Mariadassou, M.; Maass, S.; Behrendt, A.K.; Jaszczyszyn, Y.; Heilker, T.; Spaeth, M.; Vogel, S.; et al. The Rhinobiome of Exacerbated Wheezers and Asthmatics: Insights From a German Pediatric Exacerbation Network. Front. Allergy 2021, 2, 667562. [Google Scholar] [CrossRef] [PubMed]
- Sallard, E.; Schult, F.; Baehren, C.; Buedding, E.; Mboma, O.; Ahmad-Nejad, P.; Ghebremedhin, B.; Ehrhardt, A.; Wirth, S.; Aydin, M. Viral Infection and Respiratory Exacerbation in Children: Results from a Local German Pediatric Exacerbation Cohort. Viruses 2022, 14, 491. [Google Scholar] [CrossRef] [PubMed]
- Dobano, C.; Santano, R.; Jimenez, A.; Vidal, M.; Chi, J.; Rodrigo Melero, N.; Popovic, M.; Lopez-Aladid, R.; Fernandez-Barat, L.; Tortajada, M.; et al. Immunogenicity and crossreactivity of antibodies to the nucleocapsid protein of SARS-CoV-2: Utility and limitations in seroprevalence and immunity studies. Transl. Res. 2021, 232, 60–74. [Google Scholar] [CrossRef]
- Doherty, J.; Morain, N.O.; Stack, R.; Girod, P.; Tosetto, M.; Inzitiari, R.; Sheridan, J.; Cullen, G.; Mc Dermott, E.; Buckley, M.; et al. Reduced serological response to COVID-19 vaccines in patients with IBD is further diminished by TNF inhibitor therapy; Early results of the VARIATION study (VAriability in Response in IBD Against SARS-CoV-2 ImmunisatiON). J. Crohns Colitis 2022, jjac029. [Google Scholar] [CrossRef]
- Gallais, F.; Velay, A.; Nazon, C.; Wendling, M.J.; Partisani, M.; Sibilia, J.; Candon, S.; Fafi-Kremer, S. Intrafamilial Exposure to SARS-CoV-2 Associated with Cellular Immune Response without Seroconversion, France. Emerg. Infect. Dis. 2021, 27, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Kellam, P.; Barclay, W. The dynamics of humoral immune responses following SARS-CoV-2 infection and the potential for reinfection. J. Gen. Virol. 2020, 101, 791–797. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Russell, R.M.; Bibollet-Ruche, F.; Skelly, A.N.; Sherrill-Mix, S.; Freeman, D.A.; Stoltz, R.; Lindemuth, E.; Lee, F.H.; Sterrett, S.; et al. Predictors of Nonseroconversion after SARS-CoV-2 Infection. Emerg. Infect. Dis. 2021, 27, 2454–2458. [Google Scholar] [CrossRef] [PubMed]
- Niespodziana, K.; Cabauatan, C.R.; Jackson, D.J.; Gallerano, D.; Trujillo-Torralbo, B.; Del Rosario, A.; Mallia, P.; Valenta, R.; Johnston, S.L. Rhinovirus-induced VP1-specific Antibodies are Group-specific and Associated with Severity of Respiratory Symptoms. EBioMedicine 2015, 2, 64–70. [Google Scholar] [CrossRef] [Green Version]
- Saco, T.V.; Strauss, A.T.; Ledford, D.K. Hepatitis B vaccine nonresponders: Possible mechanisms and solutions. Ann. Allergy Asthma Immunol. 2018, 121, 320–327. [Google Scholar] [CrossRef]
- Aydin, M.; Schellhorn, S.; Wirth, S.; Zhang, W.; Ehrhardt, A. Human Species D Adenoviruses Isolated from Diarrheal Feces Show Low Infection Rates in Primary Nasal Epithelial Cells. Children 2021, 8, 563. [Google Scholar] [CrossRef]
- Huang, H.J.; Campana, R.; Akinfenwa, O.; Curin, M.; Sarzsinszky, E.; Karsonova, A.; Riabova, K.; Karaulov, A.; Niespodziana, K.; Elisyutina, O.; et al. Microarray-Based Allergy Diagnosis: Quo Vadis? Front. Immunol. 2020, 11, 594978. [Google Scholar] [CrossRef] [PubMed]
- Borochova, K.; Niespodziana, K.; Stenberg Hammar, K.; van Hage, M.; Hedlin, G.; Soderhall, C.; Focke-Tejkl, M.; Valenta, R. Features of the Human Antibody Response against the Respiratory Syncytial Virus Surface Glycoprotein G. Vaccines 2020, 8, 337. [Google Scholar] [CrossRef] [PubMed]
- Niespodziana, K.; Stenberg-Hammar, K.; Megremis, S.; Cabauatan, C.R.; Napora-Wijata, K.; Vacal, P.C.; Gallerano, D.; Lupinek, C.; Ebner, D.; Schlederer, T.; et al. PreDicta chip-based high resolution diagnosis of rhinovirus-induced wheeze. Nat. Commun. 2018, 9, 2382. [Google Scholar] [CrossRef]
- Gallerano, D.; Wollmann, E.; Lupinek, C.; Schlederer, T.; Ebner, D.; Harwanegg, C.; Niespodziana, K.; Schmetterer, K.; Pickl, W.; Puchhammer-Stockl, E.; et al. HIV microarray for the mapping and characterization of HIV-specific antibody responses. Lab Chip 2015, 15, 1574–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Hadley Wickham, R.F. Lionel Henry and Kirill Müller Dplyr: A Grammar of Data Manipulation; R Package Version 1.0.7; 2021. Available online: https://CRAN.R-project.org/package=dplyr (accessed on 10 June 2022).
- Elkin, L.; Kay, M.; Higgins, J.; Wobbrock, J. An Aligned Rank Transform Procedure for Multifactor Contrast Tests. In Proceedings of the The 34th Annual ACM Symposium on User Interface Software and Technology, New York, NY, USA, 10–14 October 2021. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Megremis, S.; Niespodziana, K.; Cabauatan, C.; Xepapadaki, P.; Kowalski, M.L.; Jartti, T.; Bachert, C.; Finotto, S.; West, P.; Stamataki, S.; et al. Rhinovirus Species-Specific Antibodies Differentially Reflect Clinical Outcomes in Health and Asthma. Am. J. Respir. Crit. Care Med. 2018, 198, 1490–1499. [Google Scholar] [CrossRef] [Green Version]
- Erkkola, R.; Turunen, R.; Raisanen, K.; Waris, M.; Vuorinen, T.; Laine, M.; Tahtinen, P.; Gern, J.E.; Bochkov, Y.A.; Ruohola, A.; et al. Rhinovirus C Is Associated with Severe Wheezing and Febrile Respiratory Illness in Young Children. Pediatr. Infect. Dis. J. 2020, 39, 283–286. [Google Scholar] [CrossRef]
- Turunen, R.; Jartti, T.; Bochkov, Y.A.; Gern, J.E.; Vuorinen, T. Rhinovirus species and clinical characteristics in the first wheezing episode in children. J. Med. Virol. 2016, 88, 2059–2068. [Google Scholar] [CrossRef]
- Kozik, A.J.; Huang, Y.J. The microbiome in asthma: Role in pathogenesis, phenotype, and response to treatment. Ann. Allergy Asthma Immunol. 2019, 122, 270–275. [Google Scholar] [CrossRef] [Green Version]
- Moffatt, M.F.; Cookson, W.O. The lung microbiome in health and disease. Clin. Med. 2017, 17, 525–529. [Google Scholar] [CrossRef]
- Sullivan, A.; Hunt, E.; MacSharry, J.; Murphy, D.M. The Microbiome and the Pathophysiology of Asthma. Respir. Res. 2016, 17, 163. [Google Scholar] [CrossRef] [Green Version]
- Ver Heul, A.; Planer, J.; Kau, A.L. The Human Microbiota and Asthma. Clin. Rev. Allergy Immunol. 2019, 57, 350–363. [Google Scholar] [CrossRef]
- O’Sullivan, J.A.; Bochner, B.S. Eosinophils and eosinophil-associated diseases: An update. J. Allergy Clin. Immunol. 2018, 141, 505–517. [Google Scholar] [CrossRef] [Green Version]
- Gould, H.J.; Sutton, B.J. IgE in allergy and asthma today. Nat. Rev. Immunol. 2008, 8, 205–217. [Google Scholar] [CrossRef]
- Hasegawa, K.; Craig, S.S.; Teach, S.J.; Camargo, C.A., Jr. Management of Asthma Exacerbations in the Emergency Department. J. Allergy Clin. Immunol. Pract. 2021, 9, 2599–2610. [Google Scholar] [CrossRef]
- Para, O.; Montagnani, A.; Guidi, S.; Bertu, L.; Manfellotto, D.; Campanini, M.; Fontanella, A.; Dentali, F.; Group, F.A.-E.S. Hospitalization and mortality for acute exacerbation of asthma: An Italian population-based study. Intern. Emerg. Med. 2022, 17, 1107–1113. [Google Scholar] [CrossRef]
- Jackson, D.J.; Gangnon, R.E.; Evans, M.D.; Roberg, K.A.; Anderson, E.L.; Pappas, T.E.; Printz, M.C.; Lee, W.M.; Shult, P.A.; Reisdorf, E.; et al. Wheezing rhinovirus illnesses in early life predict asthma development in high-risk children. Am. J. Respir. Crit. Care Med. 2008, 178, 667–672. [Google Scholar] [CrossRef] [Green Version]
- Ling, K.M.; Garratt, L.W.; Gill, E.E.; Lee, A.H.Y.; Agudelo-Romero, P.; Sutanto, E.N.; Iosifidis, T.; Rosenow, T.; Turvey, S.E.; Lassmann, T.; et al. Rhinovirus Infection Drives Complex Host Airway Molecular Responses in Children With Cystic Fibrosis. Front. Immunol. 2020, 11, 1327. [Google Scholar] [CrossRef]
- Michi, A.N.; Yipp, B.G.; Dufour, A.; Lopes, F.; Proud, D. PGC-1alpha mediates a metabolic host defense response in human airway epithelium during rhinovirus infections. Nat. Commun. 2021, 12, 3669. [Google Scholar] [CrossRef]
- Rubner, F.J.; Jackson, D.J.; Evans, M.D.; Gangnon, R.E.; Tisler, C.J.; Pappas, T.E.; Gern, J.E.; Lemanske, R.F., Jr. Early life rhinovirus wheezing, allergic sensitization, and asthma risk at adolescence. J. Allergy Clin. Immunol. 2017, 139, 501–507. [Google Scholar] [CrossRef] [Green Version]
- Tozzi, A.E.; Del Chierico, F.; Pandolfi, E.; Reddel, S.; Gesualdo, F.; Gardini, S.; Guarrasi, V.; Russo, L.; Croci, I.; Campagna, I.; et al. Nasopharyngeal microbiota in hospitalized children with Bordetella pertussis and Rhinovirus infection. Sci. Rep. 2021, 11, 22858. [Google Scholar] [CrossRef]
- Bizzintino, J.; Lee, W.M.; Laing, I.A.; Vang, F.; Pappas, T.; Zhang, G.; Martin, A.C.; Khoo, S.K.; Cox, D.W.; Geelhoed, G.C.; et al. Association between human rhinovirus C and severity of acute asthma in children. Eur. Respir J. 2011, 37, 1037–1042. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.T.; Lin, Y.T.; Yang, C.C.; Tsai, S.S.; Wang, J.Y.; Huang, Y.L.; Lin, T.I.; Lin, T.M.; Tsai, Y.C.; Yu, H.R.; et al. High correlation between human rhinovirus type C and children with asthma exacerbations in Taiwan. J. Microbiol. Immunol. Infect. 2020, 53, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.K.; Khuri-Bulos, N.; Williams, J.V.; Shehabi, A.A.; Faouri, S.; Al Jundi, I.; Chen, Q.; Heil, L.; Mohamed, Y.; Morin, L.L.; et al. Human rhinovirus C associated with wheezing in hospitalised children in the Middle East. J. Clin. Virol. 2009, 46, 85–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.K.; Hegele, R.G.; Manfreda, J.; Wooldrage, K.; Becker, A.B.; Ferguson, A.C.; Dimich-Ward, H.; Watson, W.T.; Chan-Yeung, M. Relationship of early childhood viral exposures to respiratory symptoms, onset of possible asthma and atopy in high risk children: The Canadian Asthma Primary Prevention Study. Pediatr. Pulmonol. 2007, 42, 290–297. [Google Scholar] [CrossRef]
- Lemanske, R.F., Jr.; Jackson, D.J.; Gangnon, R.E.; Evans, M.D.; Li, Z.; Shult, P.A.; Kirk, C.J.; Reisdorf, E.; Roberg, K.A.; Anderson, E.L.; et al. Rhinovirus illnesses during infancy predict subsequent childhood wheezing. J. Allergy Clin. Immunol. 2005, 116, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Jartti, T.; Kuusipalo, H.; Vuorinen, T.; Soderlund-Venermo, M.; Allander, T.; Waris, M.; Hartiala, J.; Ruuskanen, O. Allergic sensitization is associated with rhinovirus-, but not other virus-, induced wheezing in children. Pediatr. Allergy Immunol. 2010, 21, 1008–1014. [Google Scholar] [CrossRef]
- Mack, I.; Kieninger, E.; Cangiano, G.; Tapparel, C.; Kuehni, C.; Spycher, B.; Kaiser, L.; Frey, U.; Regamey, N.; Latzin, P. Rhinovirus Infections and Associated Respiratory Morbidity in Infants: A Prospective Cohort Study. Pediatr. Infect. Dis. J. 2016, 35, 1069–1074. [Google Scholar] [CrossRef] [Green Version]
- Miller, E.K.; Williams, J.V.; Gebretsadik, T.; Carroll, K.N.; Dupont, W.D.; Mohamed, Y.A.; Morin, L.L.; Heil, L.; Minton, P.A.; Woodward, K.; et al. Host and viral factors associated with severity of human rhinovirus-associated infant respiratory tract illness. J. Allergy Clin. Immunol. 2011, 127, 883–891. [Google Scholar] [CrossRef]
- Jackson, D.J.; Evans, M.D.; Gangnon, R.E.; Tisler, C.J.; Pappas, T.E.; Lee, W.M.; Gern, J.E.; Lemanske, R.F., Jr. Evidence for a causal relationship between allergic sensitization and rhinovirus wheezing in early life. Am. J. Respir. Crit. Care Med. 2012, 185, 281–285. [Google Scholar] [CrossRef] [Green Version]
- Martinez, F.D.; Wright, A.L.; Taussig, L.M.; Holberg, C.J.; Halonen, M.; Morgan, W.J. Asthma and wheezing in the first six years of life. The Group Health Medical Associates. N. Engl. J. Med. 1995, 332, 133–138. [Google Scholar] [CrossRef] [Green Version]
- Cabrera, T.C.; Mendez, S.A.; Gonzalez, M.M.L.; Garcia, D.G.; Garcia-Marcos, L.; Galindo, P.V.; Belinchon, J.P. Childhood wheezing phenotypes and their association to perinatal factors: A longitudinal study. Allergol. Immunopathol. 2021, 49, 8–16. [Google Scholar] [CrossRef]
- De Sario, M.; Di Domenicantonio, R.; Corbo, G.; Forastiere, F.; Pistelli, R.; Rusconi, F.; Sammarro, S.; Serra, M.G.; Compagnucci, P.; Perucci, C.A. Characteristics of early transient, persistent, and late onset wheezers at 9 to 11 years of age. J. Asthma 2006, 43, 633–638. [Google Scholar] [CrossRef]
- Caudri, D.; Savenije, O.E.; Smit, H.A.; Postma, D.S.; Koppelman, G.H.; Wijga, A.H.; Kerkhof, M.; Gehring, U.; Hoekstra, M.O.; Brunekreef, B.; et al. Perinatal risk factors for wheezing phenotypes in the first 8 years of life. Clin. Exp. Allergy 2013, 43, 1395–1405. [Google Scholar] [CrossRef]
- Hallit, S.; Leynaert, B.; Delmas, M.C.; Rocchi, S.; De Blic, J.; Marguet, C.; Scherer, E.; Dufourg, M.N.; Bois, C.; Reboux, G.; et al. Wheezing phenotypes and risk factors in early life: The ELFE cohort. PLoS ONE 2018, 13, e0196711. [Google Scholar] [CrossRef] [Green Version]
- Severcan, E.U.; Bal, C.M.; Gulen, F.; Tanac, R.; Demir, E. Identifying wheezing phenotypes in a pediatric Turkish cohort. J. Asthma 2022, 59, 1298–1304. [Google Scholar] [CrossRef]
- Justiz Vaillant, A.A.; Wilson, A.M. Transient Hypogammaglobulinemia of Infancy; StatPearls: Treasure Island, FL, USA, 2022. [Google Scholar]
- Stenberg-Hammar, K.; Niespodziana, K.; Soderhall, C.; James, A.; Cabauatan, C.R.; Konradsen, J.R.; Melen, E.; van Hage, M.; Valenta, R.; Hedlin, G. Rhinovirus-specific antibody responses in preschool children with acute wheeze reflect severity of respiratory symptoms. Allergy 2016, 71, 1728–1735. [Google Scholar] [CrossRef]
- Allen, E.K.; Koeppel, A.F.; Hendley, J.O.; Turner, S.D.; Winther, B.; Sale, M.M. Characterization of the nasopharyngeal microbiota in health and during rhinovirus challenge. Microbiome 2014, 2, 22. [Google Scholar] [CrossRef] [Green Version]
- Bosch, A.A.; Biesbroek, G.; Trzcinski, K.; Sanders, E.A.; Bogaert, D. Viral and bacterial interactions in the upper respiratory tract. PLoS Pathog. 2013, 9, e1003057. [Google Scholar] [CrossRef] [Green Version]
- Hofstra, J.J.; Matamoros, S.; van de Pol, M.A.; de Wever, B.; Tanck, M.W.; Wendt-Knol, H.; Deijs, M.; van der Hoek, L.; Wolthers, K.C.; Molenkamp, R.; et al. Changes in microbiota during experimental human Rhinovirus infection. BMC Infect. Dis. 2015, 15, 336. [Google Scholar] [CrossRef] [Green Version]
- Korten, I.; Mika, M.; Klenja, S.; Kieninger, E.; Mack, I.; Barbani, M.T.; Gorgievski, M.; Frey, U.; Hilty, M.; Latzin, P. Interactions of Respiratory Viruses and the Nasal Microbiota during the First Year of Life in Healthy Infants. mSphere 2016, 1, e00312-16. [Google Scholar] [CrossRef] [Green Version]
- Lehtinen, M.J.; Hibberd, A.A.; Mannikko, S.; Yeung, N.; Kauko, T.; Forssten, S.; Lehtoranta, L.; Lahtinen, S.J.; Stahl, B.; Lyra, A.; et al. Nasal microbiota clusters associate with inflammatory response, viral load, and symptom severity in experimental rhinovirus challenge. Sci. Rep. 2018, 8, 11411. [Google Scholar] [CrossRef] [Green Version]
- McCauley, K.; Durack, J.; Valladares, R.; Fadrosh, D.W.; Lin, D.L.; Calatroni, A.; LeBeau, P.K.; Tran, H.T.; Fujimura, K.E.; LaMere, B.; et al. Distinct nasal airway bacterial microbiotas differentially relate to exacerbation in pediatric patients with asthma. J. Allergy Clin. Immunol. 2019, 144, 1187–1197. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sallard, E.; Niespodziana, K.; Bajic, M.; Schlederer, T.; Errhalt, P.; Behrendt, A.-K.; Wirth, S.; Meyer-Bahlburg, A.; Ehrhardt, A.; Valenta, R.; et al. Microarray-Based Analyses of Rhinovirus Species-Specific Antibody Responses in Exacerbated Pediatric Asthma in a German Pediatric Cohort. Viruses 2022, 14, 1857. https://doi.org/10.3390/v14091857
Sallard E, Niespodziana K, Bajic M, Schlederer T, Errhalt P, Behrendt A-K, Wirth S, Meyer-Bahlburg A, Ehrhardt A, Valenta R, et al. Microarray-Based Analyses of Rhinovirus Species-Specific Antibody Responses in Exacerbated Pediatric Asthma in a German Pediatric Cohort. Viruses. 2022; 14(9):1857. https://doi.org/10.3390/v14091857
Chicago/Turabian StyleSallard, Erwan, Katarzyna Niespodziana, Maja Bajic, Thomas Schlederer, Peter Errhalt, Ann-Kathrin Behrendt, Stefan Wirth, Almut Meyer-Bahlburg, Anja Ehrhardt, Rudolf Valenta, and et al. 2022. "Microarray-Based Analyses of Rhinovirus Species-Specific Antibody Responses in Exacerbated Pediatric Asthma in a German Pediatric Cohort" Viruses 14, no. 9: 1857. https://doi.org/10.3390/v14091857