
Human Rhinoviruses in Pediatric Patients in a Tertiary Care Hospital in Germany: Molecular Epidemiology and Clinical Significance


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Clinical Data
2.2. Nucleic Acid (NA) Extraction and RV Detection
2.3. Rhinovirus Genotyping and Phylogenetic Analysis
2.4. Statistical Analysis
3. Results
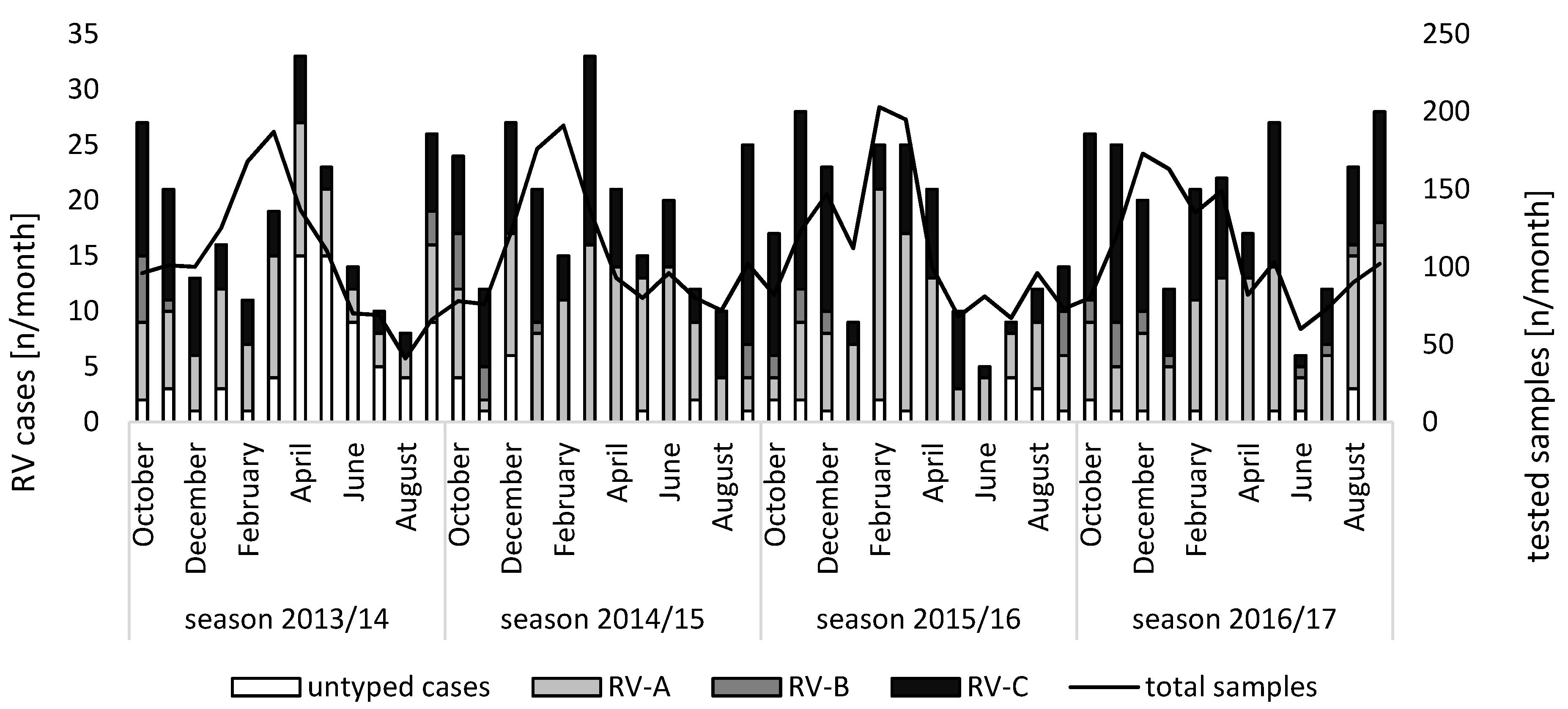
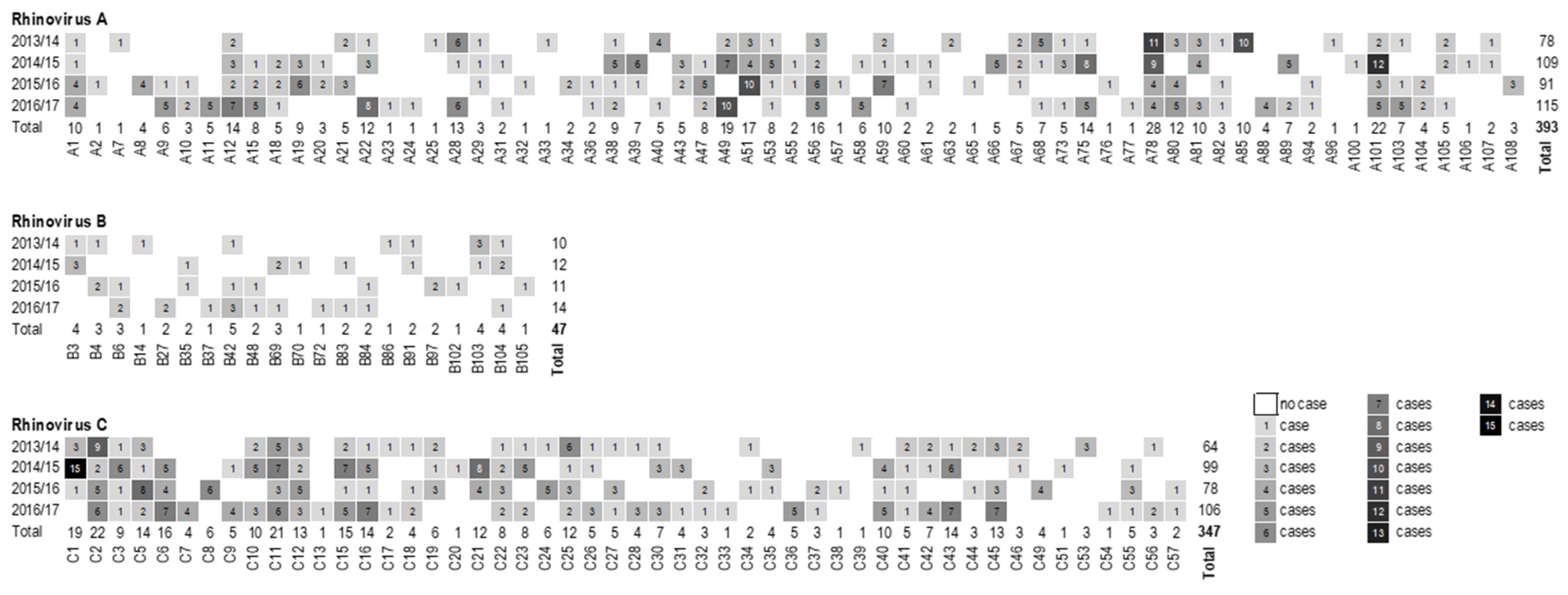
3.1. Species Distribution and Seasonality
3.2. Study Population and Clinical Features
3.3. Co-Infections
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hayden, F.G. Rhinovirus and the lower respiratory tract. Rev. Med. Virol. 2004, 14, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, S.E.; Lamson, D.M.; St George, K.; Walsh, T.J. Human rhinoviruses. Clin. Microbiol. Rev. 2013, 26, 135–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winther, B. Rhinovirus infections in the upper airway. Proc. Am. Thorac. Soc. 2011, 8, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Kiang, D.; Yagi, S.; Kantardjieff, K.A.; Kim, E.J.; Louie, J.K.; Schnurr, D.P. Molecular characterization of a variant rhinovirus from an outbreak associated with uncommonly high mortality. J. Clin. Virol. 2007, 38, 227–237. [Google Scholar] [CrossRef]
- Henquell, C.; Mirand, A.; Deusebis, A.-L.; Regagnon, C.; Archimbaud, C.; Chambon, M.; Bailly, J.-L.; Gourdon, F.; Hermet, E.; Dauphin, J.-B.; et al. Prospective genotyping of human rhinoviruses in children and adults during the winter of 2009–2010. J. Clin. Virol. 2012, 53, 280–284. [Google Scholar] [CrossRef]
- Ison, M.G.; Hayden, F.G.; Kaiser, L.; Corey, L.; Boeckh, M. Rhinovirus infections in hematopoietic stem cell transplant recipients with pneumonia. Clin. Infect. Dis. 2003, 36, 1139–1143. [Google Scholar] [CrossRef] [Green Version]
- Sallard, E.; Schult, F.; Baehren, C.; Buedding, E.; Mboma, O.; Ahmad-Nejad, P.; Ghebremedhin, B.; Ehrhardt, A.; Wirth, S.; Aydin, M. Viral Infection and Respiratory Exacerbation in Children: Results from a Local German Pediatric Exacerbation Cohort. Viruses 2022, 14, 491. [Google Scholar] [CrossRef]
- Biagi, C.; Rocca, A.; Poletti, G.; Fabi, M.; Lanari, M. Rhinovirus Infection in Children with Acute Bronchiolitis and Its Impact on Recurrent Wheezing and Asthma Development. Microorganisms 2020, 8, 1620. [Google Scholar] [CrossRef]
- Liu, L.; Pan, Y.; Zhu, Y.; Song, Y.; Su, X.; Yang, L.; Li, M. Association between rhinovirus wheezing illness and the development of childhood asthma: A meta-analysis. BMJ Open 2017, 7, e013034. [Google Scholar] [CrossRef] [Green Version]
- Friedlander, S.L.; Busse, W.W. The role of rhinovirus in asthma exacerbations. J. Allergy Clin. Immunol. 2005, 116, 267–273. [Google Scholar] [CrossRef]
- McManus, T.E.; Marley, A.-M.; Baxter, N.; Christie, S.N.; O’Neill, H.J.; Elborn, J.S.; Coyle, P.V.; Kidney, J.C. Respiratory viral infection in exacerbations of COPD. Respir. Med. 2008, 102, 1575–1580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leigh, R.; Proud, D. Virus-induced modulation of lower airway diseases: Pathogenesis and pharmacologic approaches to treatment. Pharmacol. Ther. 2015, 148, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Kurai, D.; Saraya, T.; Ishii, H.; Takizawa, H. Virus-induced exacerbations in asthma and COPD. Front. Microbiol. 2013, 4, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Picornavirus Pages. Picornavirus Classification. Available online: https://www.picornaviridae.com/ (accessed on 27 October 2021).
- Lamson, D.; Renwick, N.; Kapoor, V.; Liu, Z.; Palacios, G.; Ju, J.; Dean, A.; St George, K.; Briese, T.; Lipkin, W.I. MassTag polymerase-chain-reaction detection of respiratory pathogens, including a new rhinovirus genotype, that caused influenza-like illness in New York State during 2004–2005. J. Infect. Dis. 2006, 194, 1398–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McErlean, P.; Shackelton, L.A.; Lambert, S.B.; Nissen, M.D.; Sloots, T.P.; Mackay, I.M. Characterisation of a newly identified human rhinovirus, HRV-QPM, discovered in infants with bronchiolitis. J. Clin. Virol. 2007, 39, 67–75. [Google Scholar] [CrossRef]
- Lau, S.K.P.; Yip, C.C.Y.; Tsoi, H.-W.; Lee, R.A.; So, L.-Y.; Lau, Y.-L.; Chan, K.-H.; Woo, P.C.Y.; Yuen, K.-Y. Clinical features and complete genome characterization of a distinct human rhinovirus (HRV) genetic cluster, probably representing a previously undetected HRV species, HRV-C, associated with acute respiratory illness in children. J. Clin. Microbiol. 2007, 45, 3655–3664. [Google Scholar] [CrossRef] [Green Version]
- Stobart, C.C.; Nosek, J.M.; Moore, M.L. Rhinovirus Biology, Antigenic Diversity, and Advancements in the Design of a Human Rhinovirus Vaccine. Front. Microbiol. 2017, 8, 2412. [Google Scholar] [CrossRef]
- Bochkov, Y.A.; Watters, K.; Ashraf, S.; Griggs, T.F.; Devries, M.K.; Jackson, D.J.; Palmenberg, A.C.; Gern, J.E. Cadherin-related family member 3, a childhood asthma susceptibility gene product, mediates rhinovirus C binding and replication. Proc. Natl. Acad. Sci. USA 2015, 112, 5485–5490. [Google Scholar] [CrossRef] [Green Version]
- McIntyre, C.L.; McWilliam Leitch, E.C.; Savolainen-Kopra, C.; Hovi, T.; Simmonds, P. Analysis of genetic diversity and sites of recombination in human rhinovirus species C. J. Virol. 2010, 84, 10297–10310. [Google Scholar] [CrossRef] [Green Version]
- McIntyre, C.L.; Knowles, N.J.; Simmonds, P. Proposals for the classification of human rhinovirus species A, B and C into genotypically assigned types. J. Gen. Virol. 2013, 94, 1791–1806. [Google Scholar] [CrossRef]
- Davison, A.J.; Dolan, A.; Akter, P.; Addison, C.; Dargan, D.J.; Alcendor, D.J.; McGeoch, D.J.; Hayward, G.S. The human cytomegalovirus genome revisited: Comparison with the chimpanzee cytomegalovirus genome. J. Gen. Virol. 2003, 84, 17–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golke, P.; Hönemann, M.; Bergs, S.; Liebert, U.G. Human Rhinoviruses in Adult Patients in a Tertiary Care Hospital in Germany: Molecular Epidemiology and Clinical Significance. Viruses 2021, 13, 2027. [Google Scholar] [CrossRef] [PubMed]
- Laux, G.; Perricaudet, M.; Farrell, P.J. A spliced Epstein-Barr virus gene expressed in immortalized lymphocytes is created by circularization of the linear viral genome. EMBO J. 1988, 7, 769–774. [Google Scholar] [CrossRef]
- Gompels, U.A.; Nicholas, J.; Lawrence, G.; Jones, M.; Thomson, B.J.; Martin, M.E.; Efstathiou, S.; Craxton, M.; Macaulay, H.A. The DNA sequence of human herpesvirus-6: Structure, coding content, and genome evolution. Virology 1995, 209, 29–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espy, M.J.; Ross, T.K.; Teo, R.; Svien, K.A.; Wold, A.D.; Uhl, J.R.; Smith, T.F. Evaluation of LightCycler PCR for implementation of laboratory diagnosis of herpes simplex virus infections. J. Clin. Microbiol. 2000, 38, 3116–3118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; Holloway, B.; Dare, R.K.; Kuypers, J.; Yagi, S.; Williams, J.V.; Hall, C.B.; Erdman, D.D. Real-time reverse transcription-PCR assay for comprehensive detection of human rhinoviruses. J. Clin. Microbiol. 2008, 46, 533–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mubareka, S.; Louie, L.; Wong, H.; Granados, A.; Chong, S.; Luinstra, K.; Petrich, A.; Smieja, M.; Vearncombe, M.; Mahony, J.; et al. Co-circulation of multiple genotypes of human rhinovirus during a large outbreak of respiratory illness in a veterans’ long-term care home. J. Clin. Virol. 2013, 58, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Savolainen, C.; Blomqvist, S.; Mulders, M.N.; Hovi, T. Genetic clustering of all 102 human rhinovirus prototype strains: Serotype 87 is close to human enterovirus 70. J. Gen. Virol. 2002, 83, 333–340. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Barclay, W.S.; al-Nakib, W.; Higgins, P.G.; Tyrrell, D.A. The time course of the humoral immune response to rhinovirus infection. Epidemiol. Infect. 1989, 103, 659–669. [Google Scholar] [CrossRef] [Green Version]
- Sansone, M.; Andersson, M.; Brittain-Long, R.; Andersson, L.-M.; Olofsson, S.; Westin, J.; Lindh, M. Rhinovirus infections in western Sweden: A four-year molecular epidemiology study comparing local and globally appearing types. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 947–954. [Google Scholar] [CrossRef] [PubMed]
- Esneau, C.; Duff, A.C.; Bartlett, N.W. Understanding Rhinovirus Circulation and Impact on Illness. Viruses 2022, 14, 141. [Google Scholar] [CrossRef] [PubMed]
- Gwaltney, J.M.; Hendley, J.O.; Simon, G.; Jordan, W.S. Rhinovirus infections in an industrial population. I. The occurrence of illness. N. Engl. J. Med. 1966, 275, 1261–1268. [Google Scholar] [CrossRef]
- Van der Linden, L.; Bruning, A.H.L.; Thomas, X.V.; Minnaar, R.P.; Rebers, S.P.H.; Schinkel, J.; de Jong, M.D.; Pajkrt, D.; Wolthers, K.C. A molecular epidemiological perspective of rhinovirus types circulating in Amsterdam from 2007 to 2012. Clin. Microbiol. Infect. 2016, 22, 1002.e9–1002.e14. [Google Scholar] [CrossRef] [PubMed]
- Everman, J.L.; Sajuthi, S.; Saef, B.; Rios, C.; Stoner, A.M.; Numata, M.; Hu, D.; Eng, C.; Oh, S.; Rodriguez-Santana, J.; et al. Functional genomics of CDHR3 confirms its role in HRV-C infection and childhood asthma exacerbations. J. Allergy Clin. Immunol. 2019, 144, 962–971. [Google Scholar] [CrossRef] [Green Version]
- Basnet, S.; Bochkov, Y.A.; Brockman-Schneider, R.A.; Kuipers, I.; Aesif, S.W.; Jackson, D.J.; Lemanske, R.F.; Ober, C.; Palmenberg, A.C.; Gern, J.E. CDHR3 Asthma-Risk Genotype Affects Susceptibility of Airway Epithelium to Rhinovirus C Infections. Am. J. Respir. Cell Mol. Biol. 2019, 61, 450–458. [Google Scholar] [CrossRef]
- Calvo, C.; Casas, I.; García-García, M.L.; Pozo, F.; Reyes, N.; Cruz, N.; García-Cuenllas, L.; Pérez-Breña, P. Role of rhinovirus C respiratory infections in sick and healthy children in Spain. Pediatr. Infect. Dis. J. 2010, 29, 717–720. [Google Scholar] [CrossRef]
- Li, W.; Yu, B.; Zhou, J.; Wang, Y.; Xue, B.; Pan, J.; Ran, Y.; Yang, X.; Wang, X.; Yang, F.; et al. Genetic diversity and epidemiology of human rhinovirus among children with severe acute respiratory tract infection in Guangzhou, China. Virol. J. 2021, 18, 174. [Google Scholar] [CrossRef]
- Tran, D.N.; Trinh, Q.D.; Pham, N.T.K.; Pham, T.M.H.; Ha, M.T.; Nguyen, T.Q.N.; Okitsu, S.; Shimizu, H.; Hayakawa, S.; Mizuguchi, M.; et al. Human rhinovirus infections in hospitalized children: Clinical, epidemiological and virological features. Epidemiol. Infect. 2016, 144, 346–354. [Google Scholar] [CrossRef]
- L’Huillier, A.G.; Kaiser, L.; Petty, T.J.; Kilowoko, M.; Kyungu, E.; Hongoa, P.; Vieille, G.; Turin, L.; Genton, B.; D’Acremont, V.; et al. Molecular Epidemiology of Human Rhinoviruses and Enteroviruses Highlights Their Diversity in Sub-Saharan Africa. Viruses 2015, 7, 6412–6423. [Google Scholar] [CrossRef] [Green Version]
- Hung, H.-M.; Yang, S.-L.; Chen, C.-J.; Chiu, C.-H.; Kuo, C.-Y.; Huang, K.-Y.A.; Lin, T.-Y.; Hsieh, Y.-C.; Gong, Y.-N.; Tsao, K.-C.; et al. Molecular epidemiology and clinical features of rhinovirus infections among hospitalized patients in a medical center in Taiwan. J. Microbiol. Immunol. Infect. 2019, 52, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Bizzintino, J.; Lee, W.-M.; Laing, I.A.; Vang, F.; Pappas, T.; Zhang, G.; Martin, A.C.; Khoo, S.-K.; Cox, D.W.; Geelhoed, G.C.; et al. Association between human rhinovirus C and severity of acute asthma in children. Eur. Respir. J. 2011, 37, 1037–1042. [Google Scholar] [CrossRef]
- Khetsuriani, N.; Lu, X.; Teague, W.G.; Kazerouni, N.; Anderson, L.J.; Erdman, D.D. Novel human rhinoviruses and exacerbation of asthma in children. Emerg. Infect. Dis. 2008, 14, 1793–1796. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.K.; Edwards, K.M.; Weinberg, G.A.; Iwane, M.K.; Griffin, M.R.; Hall, C.B.; Zhu, Y.; Szilagyi, P.G.; Morin, L.-L.; Heil, L.H.; et al. A novel group of rhinoviruses is associated with asthma hospitalizations. J. Allergy Clin. Immunol. 2009, 123, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Adam, D.C.; Chen, X.; Scotch, M.; MacIntyre, C.R.; Dwyer, D.; Kok, J. The Molecular Epidemiology and Clinical Phylogenetics of Rhinoviruses Among Paediatric Cases in Sydney, Australia. Int. J. Infect. Dis. 2021, 110, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Choi, T.; Devries, M.; Bacharier, L.B.; Busse, W.; Camargo, C.A.; Cohen, R.; Demuri, G.P.; Evans, M.D.; Fitzpatrick, A.M.; Gergen, P.J.; et al. Enhanced Neutralizing Antibody Responses to Rhinovirus C and Age-Dependent Patterns of Infection. Am. J. Respir. Crit. Care Med. 2021, 203, 822–830. [Google Scholar] [CrossRef]
- Arden, K.E.; Greer, R.M.; Wang, C.Y.T.; Mackay, I.M. Genotypic diversity, circulation patterns and co-detections among rhinoviruses in Queensland, 2001. Access Microbiol. 2020, 2, acmi000075. [Google Scholar] [CrossRef]
- Lee, W.-M.; Lemanske, R.F.; Evans, M.D.; Vang, F.; Pappas, T.; Gangnon, R.; Jackson, D.J.; Gern, J.E. Human rhinovirus species and season of infection determine illness severity. Am. J. Respir. Crit. Care Med. 2012, 186, 886–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagome, K.; Bochkov, Y.A.; Ashraf, S.; Brockman-Schneider, R.A.; Evans, M.D.; Pasic, T.R.; Gern, J.E. Effects of rhinovirus species on viral replication and cytokine production. J. Allergy Clin. Immunol. 2014, 134, 332–341. [Google Scholar] [CrossRef] [Green Version]
- Hönemann, M.; Martin, D.; Pietsch, C.; Maier, M.; Bergs, S.; Bieck, E.; Liebert, U.G. Influenza B virus infections in Western Saxony, Germany in three consecutive seasons between 2015 and 2018: Analysis of molecular and clinical features. Vaccine 2019, 37, 6550–6557. [Google Scholar] [CrossRef]
- Martin, D.; Hönemann, M.; Liebert, U.G. Dynamics of nosocomial parainfluenza virus type 3 and influenza virus infections at a large German University Hospital between 2012 and 2019. Diagn. Microbiol. Infect. Dis. 2021, 99, 115244. [Google Scholar] [CrossRef] [PubMed]
- Baertl, S.; Pietsch, C.; Maier, M.; Hönemann, M.; Bergs, S.; Liebert, U.G. Enteroviruses in Respiratory Samples from Paediatric Patients of a Tertiary Care Hospital in Germany. Viruses 2021, 13, 882. [Google Scholar] [CrossRef] [PubMed]
- Honkinen, M.; Lahti, E.; Österback, R.; Ruuskanen, O.; Waris, M. Viruses and bacteria in sputum samples of children with community-acquired pneumonia. Clin. Microbiol. Infect. 2012, 18, 300–307. [Google Scholar] [CrossRef] [Green Version]
- Ishizuka, S.; Yamaya, M.; Suzuki, T.; Takahashi, H.; Ida, S.; Sasaki, T.; Inoue, D.; Sekizawa, K.; Nishimura, H.; Sasaki, H. Effects of rhinovirus infection on the adherence of Streptococcus pneumoniae to cultured human airway epithelial cells. J. Infect. Dis. 2003, 188, 1928–1939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.H.; Kwon, H.J.; Jang, Y.J. Rhinovirus enhances various bacterial adhesions to nasal epithelial cells simultaneously. Laryngoscope 2009, 119, 1406–1411. [Google Scholar] [CrossRef] [PubMed]
- Passariello, C.; Schippa, S.; Conti, C.; Russo, P.; Poggiali, F.; Garaci, E.; Palamara, A.T. Rhinoviruses promote internalisation of Staphylococcus aureus into non-fully permissive cultured pneumocytes. Microbes Infect. 2006, 8, 758–766. [Google Scholar] [CrossRef]
- Sajjan, U.; Wang, Q.; Zhao, Y.; Gruenert, D.C.; Hershenson, M.B. Rhinovirus disrupts the barrier function of polarized airway epithelial cells. Am. J. Respir. Crit. Care Med. 2008, 178, 1271–1281. [Google Scholar] [CrossRef] [Green Version]
- Oliver, B.G.G.; Lim, S.; Wark, P.; Laza-Stanca, V.; King, N.; Black, J.L.; Burgess, J.K.; Roth, M.; Johnston, S.L. Rhinovirus exposure impairs immune responses to bacterial products in human alveolar macrophages. Thorax 2008, 63, 519–525. [Google Scholar] [CrossRef] [Green Version]
- Jain, S.; Williams, D.J.; Arnold, S.R.; Ampofo, K.; Bramley, A.M.; Reed, C.; Stockmann, C.; Anderson, E.J.; Grijalva, C.G.; Self, W.H.; et al. Community-acquired pneumonia requiring hospitalization among U.S. children. N. Engl. J. Med. 2015, 372, 835–845. [Google Scholar] [CrossRef] [Green Version]
- Loeffelholz, M.J.; Trujillo, R.; Pyles, R.B.; Miller, A.L.; Alvarez-Fernandez, P.; Pong, D.L.; Chonmaitree, T. Duration of rhinovirus shedding in the upper respiratory tract in the first year of life. Pediatrics 2014, 134, 1144–1150. [Google Scholar] [CrossRef] [Green Version]
- Jartti, T.; Lehtinen, P.; Vuorinen, T.; Koskenvuo, M.; Ruuskanen, O. Persistence of rhinovirus and enterovirus RNA after acute respiratory illness in children. J. Med. Virol. 2004, 72, 695–699. [Google Scholar] [CrossRef] [PubMed]
- Flight, W.G.; Bright-Thomas, R.J.; Tilston, P.; Mutton, K.J.; Guiver, M.; Webb, A.K.; Jones, A.M. Chronic rhinovirus infection in an adult with cystic fibrosis. J. Clin. Microbiol. 2013, 51, 3893–3896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, L.G.; Powell, H.; Grissell, T.V.; Davies, B.; Shafren, D.R.; Whitehead, B.F.; Hensley, M.J.; Gibson, P.G. Persistence of rhinovirus RNA and IP-10 gene expression after acute asthma. Respirology 2011, 16, 291–299. [Google Scholar] [CrossRef]
- Jartti, T.; Lee, W.-M.; Pappas, T.; Evans, M.; Lemanske, R.F.; Gern, J.E. Serial viral infections in infants with recurrent respiratory illnesses. Eur. Respir. J. 2008, 32, 314–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLean, G.R. Vaccine strategies to induce broadly protective immunity to rhinoviruses. Hum. Vaccin. Immunother. 2019, 16, 684–686. [Google Scholar] [CrossRef]
- Touabi, L.; Aflatouni, F.; McLean, G.R. Mechanisms of Rhinovirus Neutralisation by Antibodies. Viruses 2021, 13, 360. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Season 2013/2014 | Season 2014/2015 | Season 2015/2016 | Season 2016/2017 | Total | p-Value | |||
|---|---|---|---|---|---|---|---|---|
| age (years) | [mean ± SD] | 1.99 ± 2.68 | 2.58 ± 3.31 | 2.07 ± 3.15 | 2.8 ± 3.9 | 2.42 ± 3.37 | n.s. | |
| male | [% (n/total)] | 61.5 (91/148) | 60.3 (132/219) | 58.3 (105/180) | 55.5 (127/229) | 58.6 (455/776) |  | n.s. |
| female | [% (n/total)] | 38.5 (57/148) | 39.7 (87/219) | 41.7 (75/180) | 44.5 (102/229) | 41.4 (321/776) | ||
| RV species | ||||||||
| RV-A | [% (n/total)] | 51.4 (76/148) | 49.3 (108/220) | 49.4 (89/180) | 47.2 (108/229) | 49.1 (381/776) |  | |
| RV-B | [% (n/total)] | 6.1 (9/148) | 5.5 (12/220) | 6.1 (11/180) | 6.1 (14/229) | 5.9 (46/776) | n.s. | |
| RV-C | [% (n/total)] | 39.9 (59/148) | 44.7 (98/220) | 41.7 (75/180) | 46.3 (106/229) | 43.6 (338/776) | ||
| multiple | [% (n/total)] | 2.7 (4/148) | 0.5 (1/220) | 2.8 (5/180) | 0.4 (1/229) | 1.4 (11/776) | ||
| RV-A 49.8% (381/765) | RV-B 6% (46/765) | RV-C 44.2% (338/765) | Total | p-Value | |||
|---|---|---|---|---|---|---|---|
| study population | |||||||
| female | [% (n/total)] | 39.4 (150/381) | 43.5 (20/46) | 42.3 (143/338) | 40.9 (313/765) |  | n.s. |
| male | [% (n/total)] | 60.6 (231/381) | 56.5 (26/46) | 57.7 (195/338) | 59.1 (452/765) | ||
| age (years) | [mean ± SD] | 2.62 ± 3,57 | 3.5 ± 5.17 | 2.07 ± 2.77 | 2.43 ± 3.38 | n.s. | |
| inpatients | [% (n/total)] | 97.6 (372/381) | 95.7 (44/46) | 98.8 (334/338) | 98 (750/765) | | n.s. |
| outpatients | [% (n/total)] | 2.4 (9/381) | 4.3 (2/46) | 1.2 (4/338) | 2 (15/765) | ||
| length of stay (days) | [median(range)] | 5 (1–85) | 6 (1–77) | 5 (1–87) | 5 (1–87) | 0.007 | |
| comorbidities and risk factors | |||||||
| asthma | [% (n/total)] | 7.4 (28/377) | 4.3 (2/46) | 14.3 (48/336) | 10.3 (78/759) | 0.004 | |
| COPD | [% (n/total)] | 0.5 (2/377) | 0 (0/46) | 0 (0/337) | 0.3 (2/760) | n.s. | |
| structural lung disease | [% (n/total)] | 7.4 (28/377) | 15.2 (7/46) | 5.3 (18/337) | 7.0 (53/760) | n.s. | |
| airway ass. allergy | [% (n/total)] | 9.1 (34/374) | 2.2 (1/45) | 10.1 (34/335) | 9.2 (69/754) | n.s. | |
| cardiovascular diseases | [% (n/total)] | 13.0 (49/328) | 2.2 (1/46) | 5.9 (20/337) | 9.2 (70/760) | 0.001 | |
| metabolic disease | [% (n/total)] | 4.0 (15/377) | 8.7 (4/46) | 3.3 (11/337) | 3.9 (30/760) | n.s. | |
| malignancy | [% (n/total)] | 7.7 (29/378) | 10.9 (5/46) | 3.6 (12/325) | 6.0 (46/761) | n.s. | |
| immunosuppression | [% (n/total)] | 8.6 (32/373) | 11.4 (5/44) | 6.5 (22/337) | 7.8 (59/754) | n.s. | |
| clinical presentation | |||||||
| symptomatic | [% (n/total)] | 95.5 (360/377) | 89.1 (41/46) | 96.7 (327/338) | 95.7 (728/761) | n.s. | |
| temperature | |||||||
| <37.5 °C | [% (n/total)] | 42.7 (159/372) | 39.5 (17/43) | 53.9 (181/336) | 47.5 (357/751) |  | |
| 37.5–37.9 °C | [% (n/total)] | 10.2 (38/372) | 9.3 (4/43) | 13.7 (46/336) | 11.7 (88/751) | 0.001 | |
| ≥38 °C | [% (n/total)] | 47.0 (175/372) | 51.2 (22/43) | 32.4 (109/336) | 40.7 (306/751) | ||
| dyspnea | [% (n/total)] | 54.4 (203/373) | 43.2 (19/44) | 65.7 (222/338) | 58.8 (444/755) | <0.001 | |
| bronchodilator usage | [% (n/total)] | 61.5 (230/374) | 37.2 (16/43) | 73.6 (248/337) | 65.5 (494/754) | <0.001 | |
| URTI | [% (n/total)] | 83.8 (315/376) | 75.6 (34/45) | 84.9 (286/337) | 83.8 (635/758) | n.s. | |
| pharyngitis | [% (n/total)] | 75.7 (284/375) | 68.9 (31/45) | 74.2 (250/337) | 74.6 (565/757) | n.s. | |
| rhinitis | [% (n/total)] | 55.2 (207/375) | 40.0 (18/27) | 56.1 (189/337) | 54.7 (414/757) | n.s. | |
| tonsillitis | [% (n/total)] | 22.9 (86/375) | 22.2 (10/45) | 22.6 (76/337) | 22.7 (172/757) | n.s. | |
| LRTI | [% (n/total)] | 57.7 (217/376) | 44.4 (20/45) | 64.5 (218/338) | 59.9 (455/759) | n.s. | |
| pneumonia | [% (n/total)] | 17.3 (65/375) | 13.3 (6/45) | 8.3 (28/310) | 13.1 (99/758) | 0.002 | |
| bronchiolitis | [% (n/total)] | 43.1 (162/376) | 35.6 (16/45) | 57.4 (19/338) | 49.0 (372/759) | <0.001 | |
| ICU stay | [% (n/total)] | 23.6 (89/377) | 28.3 (13/46) | 20.4 (69/338) | 22.5 (171/761) | n.s. | |
| ventilation | |||||||
| none | [% (n/total)] | 83.1 (309/372) | 86.4 (38/44) | 86.9 (291/335) | 85.0 (638/751) |  | |
| non-invasive | [% (n/total)] | 8.1 (31/372) | 4.5 (2/44) | 7.8 (26/335) | 7.9 (59/751) | n.s. | |
| invasive | [% (n/total)] | 8.4 (32/372) | 9.1 (4/44) | 5.4 (18/335) | 7.2 (54/751) | ||
| co-infection | [% (n/total)] | 42.0 (160/381) | 34.8 (16/46) | 27.8 (94/338) | 35.3 (270/765) | <0.001 | |
| bacterial | [% (n/total)] | 16.3 (62/381) | 15.2 (7/46) | 8.9 (30/338) | 12.9 (99/765) |  | |
| viral | [% (n/total)] | 20.7 (79/381) | 17.4 (8/46) | 16.6 (56/338) | 18.7 (143/765) | n.s. | |
| fungal | [% (n/total)] | 0.3 (1/381) | 0 (0/46) | 0 (0/338) | 0.1 (1/765) | ||
| combined | [% (n/total)] | 4.7 (18/381) | 2.2 (1/46) | 2.4 (8/338) | 3.5 (27/765) | ||
| Bacteria | n | Viruses | n | Fungi | n |
|---|---|---|---|---|---|
| Haemophilus influenzae | 26 | Adenovirus | 61 | Aspergillus spp. | 2 |
| Staphylococcus aureus | 19 | Bocavirus | 44 | Candida spp. | 1 |
| Streptococcus pneumoniae | 17 | RSV | 44 | ||
| Escherichia coli | 15 | Metapneumovirus | 15 | ||
| Moraxella catarrhalis | 15 | Coronavirus OC43 | 9 | ||
| Klebsiella pneumoniae | 14 | Enterovirus | 8 | ||
| Pseudomonas aeruginosa | 11 | Parainfluenzavirus type 3 | 5 | ||
| Enterobacter cloacae | 7 | Parainfluenzavirus type 4 | 5 | ||
| Klebsiella oxytoca | 7 | Coronavirus NL63 | 4 | ||
| Mycoplasma pneumoniae | 7 | Parainfluenzavirus type 1 | 4 | ||
| Chlamydophila pneumoniae | 6 | Parainfluenzavirus type 2 | 4 | ||
| Haemophilus parainfluenzae | 5 | Coronavirus HKU1 | 3 | ||
| Streptococcus pyogenes | 5 | CMV | 2 | ||
| Serratia marcescens | 4 | Influenza A H1N1 | 2 | ||
| Stenotrophomonas maltophilia | 4 | Influenza A H3N2 | 2 | ||
| Acinetobacter baumanii | 2 | EBV | 1 | ||
| Morganella morganii | 2 | HHV 6 | 1 | ||
| Bordetella pertussis | 1 | HSV 1 | 1 | ||
| Citrobacter freundii | 1 | ||||
| Enterobacter aerogenes | 1 | ||||
| Enterobacter hormaechei | 1 | ||||
| Enterobacter kobei | 1 | ||||
| Haemophilus haemolyticus | 1 | ||||
| Haemophilus parahaemolyticus | 1 | ||||
| Leclercia adecarboxylata | 1 | ||||
| Pantoea agglomerans | 1 | ||||
| Proteus mirabilis | 1 | ||||
| Pseudomonas putida | 1 |
| RV Only 64.7 (495/765) | RV + Co-Infection 35.3 (270/765) | Total | p-Value | |||
|---|---|---|---|---|---|---|
| season | ||||||
| season 2013/2014 | [% (n/total)] | 17.4 (86/495) | 21.5 (58/270) | 18.8 (144/765) |  | n.s. |
| season 2014/2015 | [% (n/total)] | 29.7 (147/495) | 26.3 (71/270) | 28.5 (218/765) | ||
| season 2015/2016 | [% (n/total)] | 21.0 (104/495) | 26.3 (71/270) | 22.9 (175/765) | ||
| season 2016/2017 | [% (n/total)] | 31.9 (158/49) | 25.9 (70/270) | 29.8 (228/765) | ||
| study population | ||||||
| female | [% (n/total)] | 42.2 (209/495) | 38.5 (104/270) | 40.9 (313/765) |  | n.s. |
| male | [% (n/total)] | 57.8 (286/495) | 61.5 (166/270) | 59.1 (452/765) | ||
| length of stay (days) | [median (range)] | 4 (1–87) | 6 (1–82) | 5 (1–87) | <0.001 | |
| RV species | ||||||
| RV-A | [% (n/total)] | 44.6 (221/495) | 59.3 (160/270) | 49.8 (381/765) |  | |
| RV-B | [% (n/total)] | 6.1 (30/495) | 5.9 (16/270) | 6.0 (46/765) | <0.001 | |
| RV-C | [% (n/total)] | 49.3 (244/495) | 34.8 (94/270) | 44.2 (338/765) | ||
| clinical presentation | ||||||
| symptomatic | [% (n/total)] | 93.9 (461/491) | 98.9 (267/270) | 95.7 (728/761) | 0.001 | |
| temperature | ||||||
| <37.5 °C | [% (n/total)] | 51.1 (246/481) | 41.1(111/270) | 47.5 (357/751) |  | |
| 37.5–37.9°C | [% (n/total)] | 11.4 (55/481) | 12.2 (33/270) | 11.7 (88/751) | 0.025 | |
| ≥38 °C | [% (n/total)] | 37.4 (180/481) | 46.7 (126/270) | 40.7 (306/751) | ||
| dyspnea | [% (n/total)] | 59.2 (287/485) | 58.1 (157/270) | 58.8 (444/755) | n.s. | |
| bronchodilator usage | [% (n/total)] | 65.1 (315/484) | 66.3 (179/270) | 65.5 (494/754) | n.s | |
| URTI | [% (n/total)] | 82.2 (401/488) | 86.7 (234/270) | 83.8 (635/758) | n.s. | |
| pharyngitis | [% (n/total)] | 71.9 (350/487) | 79.6 (215/270) | 74.6 (565/757) | 0.019 | |
| rhinitis | [% (n/total)] | 54.0 (263/487) | 55.9 (151/270) | 54.7 (414/757) | n.s. | |
| tonsillitis | [% (n/total)] | 19.9 (97/487) | 27.8 (75/270) | 22.7 (172/757) | 0.013 | |
| LRTI | [% (n/total)] | 56.6 (277/489) | 65.9 (178/270) | 59.9 (455/759) | 0.012 | |
| pneumonia | [% (n/total)] | 7.8 (38/488) | 22.6 (61/270) | 13.1 (99/758) | <0.001 | |
| bronchiolitis | [% (n/total)] | 51.1 (250/489) | 45.2 (122/270) | 49.0 (372/759) | n.s. | |
| ICU stay | [% (n/total)] | 17.7 (87/491) | 31.1 (84/270) | 22.5 (171/761) | <0.001 | |
| ventilation | ||||||
| none | [% (n/total)] | 90.4 (435/481) | 75.3 (203/270) | 85.0 (638/751) |  | |
| non-invasive | [% (n/total)] | 6.2 (30/481) | 10.7 (29/270) | 7.9 (59/751) | <0.001 | |
| invasive | [% (n/total)] | 3.3 (16/481) | 14.1 (38/270) | 7.2 (54/751) | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neugebauer, F.; Bergs, S.; Liebert, U.G.; Hönemann, M. Human Rhinoviruses in Pediatric Patients in a Tertiary Care Hospital in Germany: Molecular Epidemiology and Clinical Significance. Viruses 2022, 14, 1829. https://doi.org/10.3390/v14081829
Neugebauer F, Bergs S, Liebert UG, Hönemann M. Human Rhinoviruses in Pediatric Patients in a Tertiary Care Hospital in Germany: Molecular Epidemiology and Clinical Significance. Viruses. 2022; 14(8):1829. https://doi.org/10.3390/v14081829
Chicago/Turabian StyleNeugebauer, Franziska, Sandra Bergs, Uwe Gerd Liebert, and Mario Hönemann. 2022. "Human Rhinoviruses in Pediatric Patients in a Tertiary Care Hospital in Germany: Molecular Epidemiology and Clinical Significance" Viruses 14, no. 8: 1829. https://doi.org/10.3390/v14081829