
Two Consecutive Prolines in the Fusion Peptide of Murine β-Coronavirus Spike Protein Predominantly Determine Fusogenicity and May Be Essential but Not Sufficient to Cause Demyelination

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses
2.2. Isolation and Enrichment of Primary Neuron from Neonatal Day 0 Mouse Brain
2.3. Maintenance of Secondary Cells in Culture
2.4. Viral Infection in Primary Neuronal Cells, Lung Epithelial Cells L2, Neuroblastoma Cells Neuro2A, and Astrocytoma Cells DBT
2.5. Immunofluorescence on Cultured Cells
2.6. Quantification of RSA59 (PP), RSMHV2 (P), and RSMHV2 (PP) Induced Cell-to-Cell Fusion In Vitro
2.7. Inoculation of Mice
2.8. Histopathological Analyses
2.9. Immunohistochemical Analysis and Quantification of Viral Antigen
2.10. Immunofluorescence on Spinal Cord Cryosections

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibody | Dilution | Secondary Antibody | Dilution |
|---|---|---|---|
| Rabbit polyclonal Anti-MAP2 (Sigma) | 1:200 | Alexa fluor 568 Donkey Anti Rabbit (Invitrogen) | 1:800 |
| Mouse monoclonal Anti-NFM (Sigma) | 1:200 | FITC Goat Anti Mouse IgG (Jackson immunoresearch, West Grove, PA, USA) | 1:250 |
| Mouse monoclonal Anti-GFAP (Sigma) | 1:200 | FITC Goat Anti Mouse IgG (Jackson immunoresearch, West Grove, PA, USA) | 1:250 |
| Anti-H8H9 (same as anti-Gal C) (mouse monoclonal antibody against mature Oligodendrocytes) [34] | 1:50 | FITC Goat Anti Mouse IgG (Jackson immunoresearch, West Grove, PA, USA) | 1:250 |
| Anti-N (anti-nucleocapsid of MHV) (Gift from Dr. Julian Leibowitz of Texas A&M, College Station, TX) | 1:25 | FITC Goat Anti Mouse IgG (Jackson immunoresearch, West Grove, PA, USA) | 1:250 |
2.11. Detection of Demyelination and Quantification
2.12. Structure Modeling and Molecular Dynamics Simulation of RSMHV2 (PP)
2.13. Statistical Analysis
3. Results
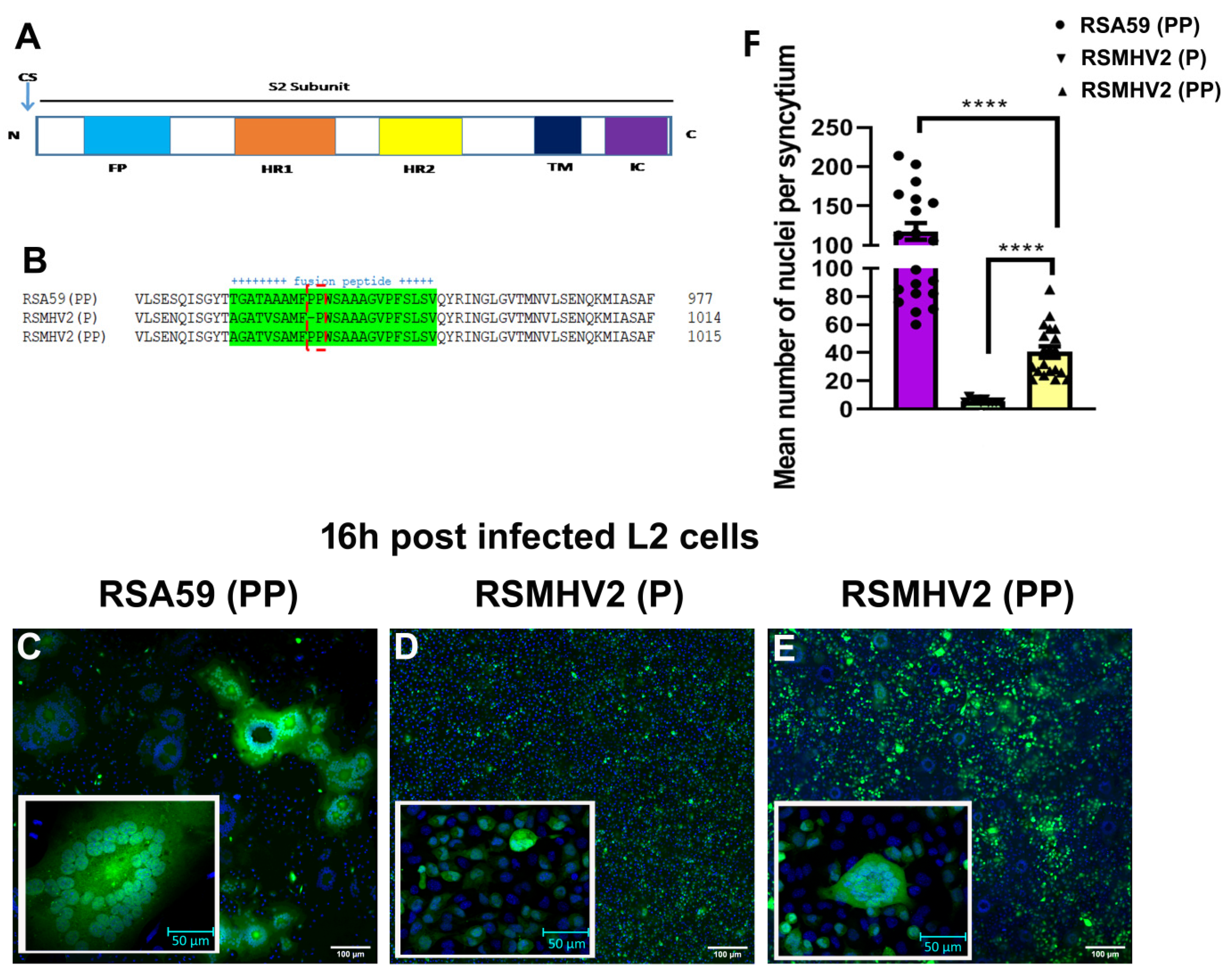
3.1. Sequence Comparison between RSA59 (PP), RSMHV2 (P), RSMHV2 (PP) Fusion Peptide
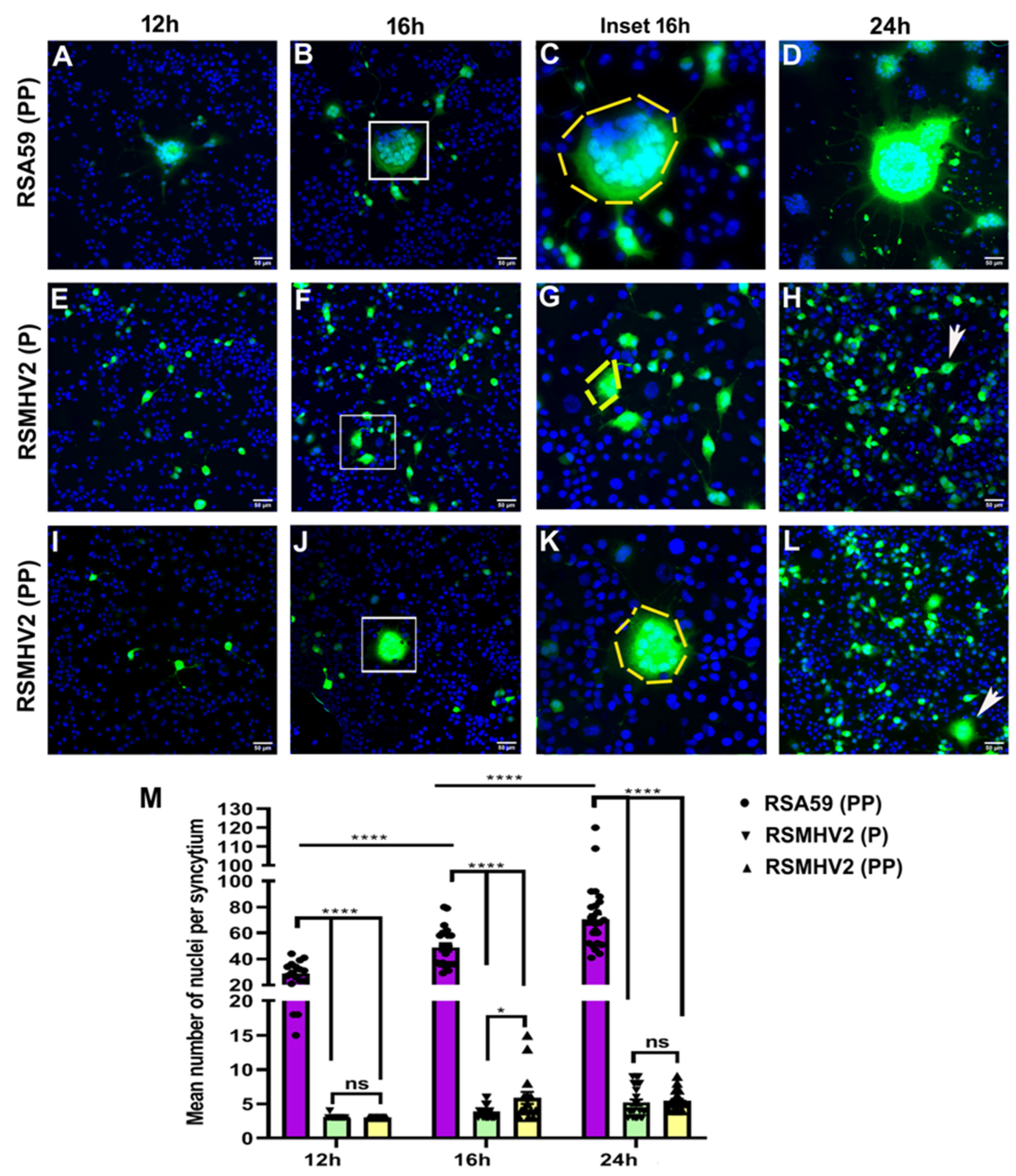
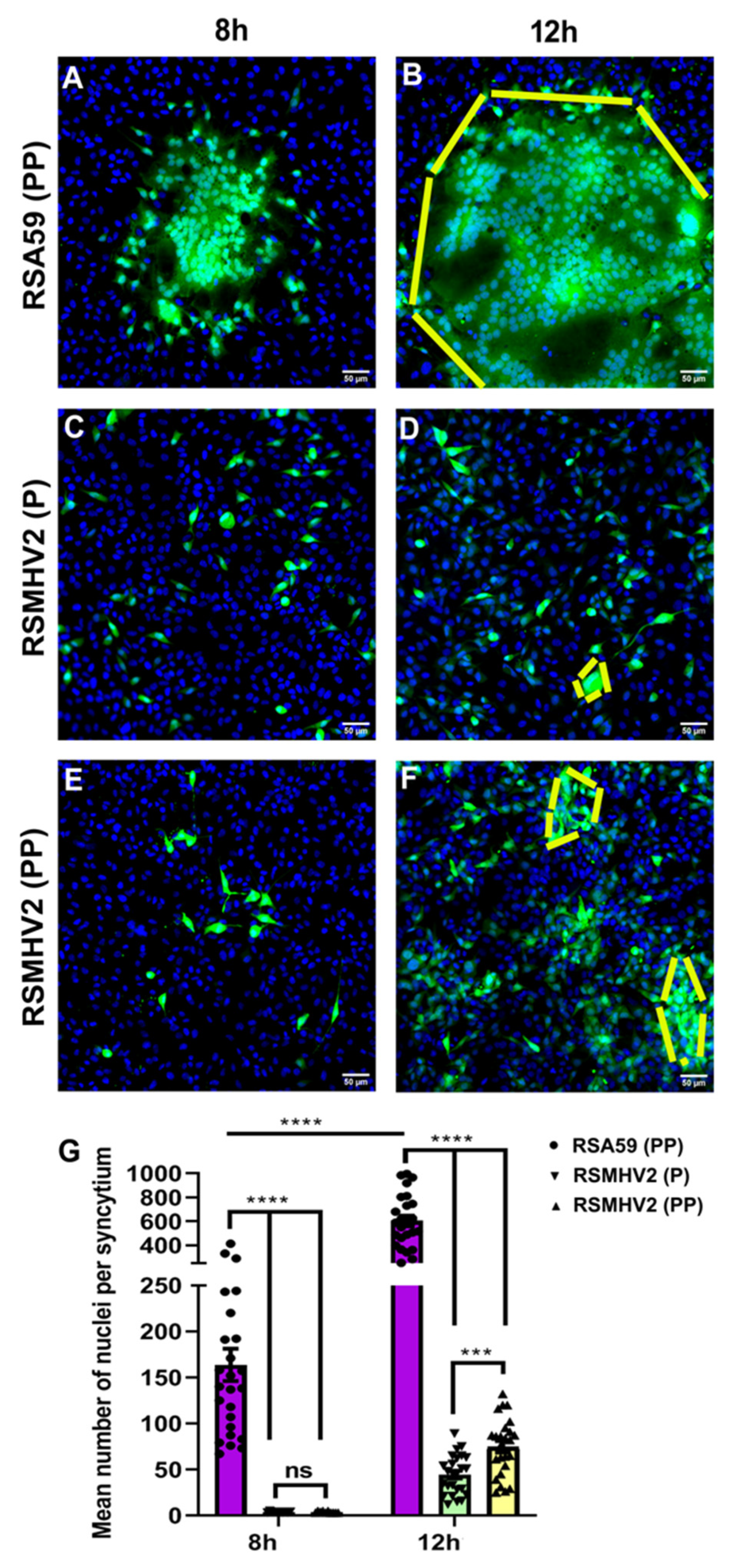
3.2. Divergent Fusogenicity of RSA59 (PP), RSMHV2 (P), and RSMHV2 (PP) in L2 Cells
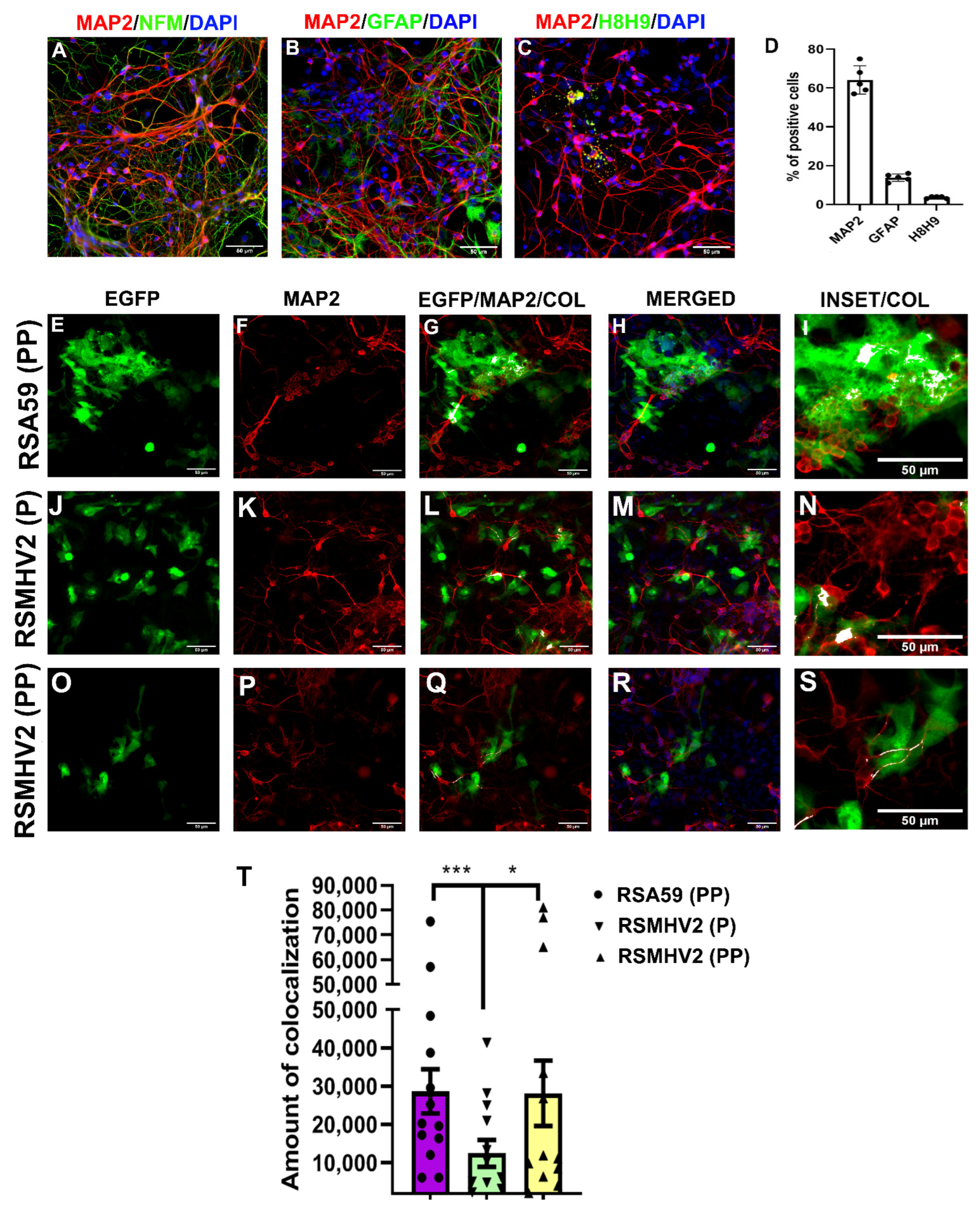
3.3. RSA59 (PP), RSMHV2 (P), and RSMHV2 (PP) Differ in Their Neuronal Tropism, Spread through Neuron and Syncytia Formation in Primary Neuronal Culture, Neuroblastoma Cell Line, Neuro2a, and Delayed Brain Tumor (DBT), Astrocytoma Cell Lines
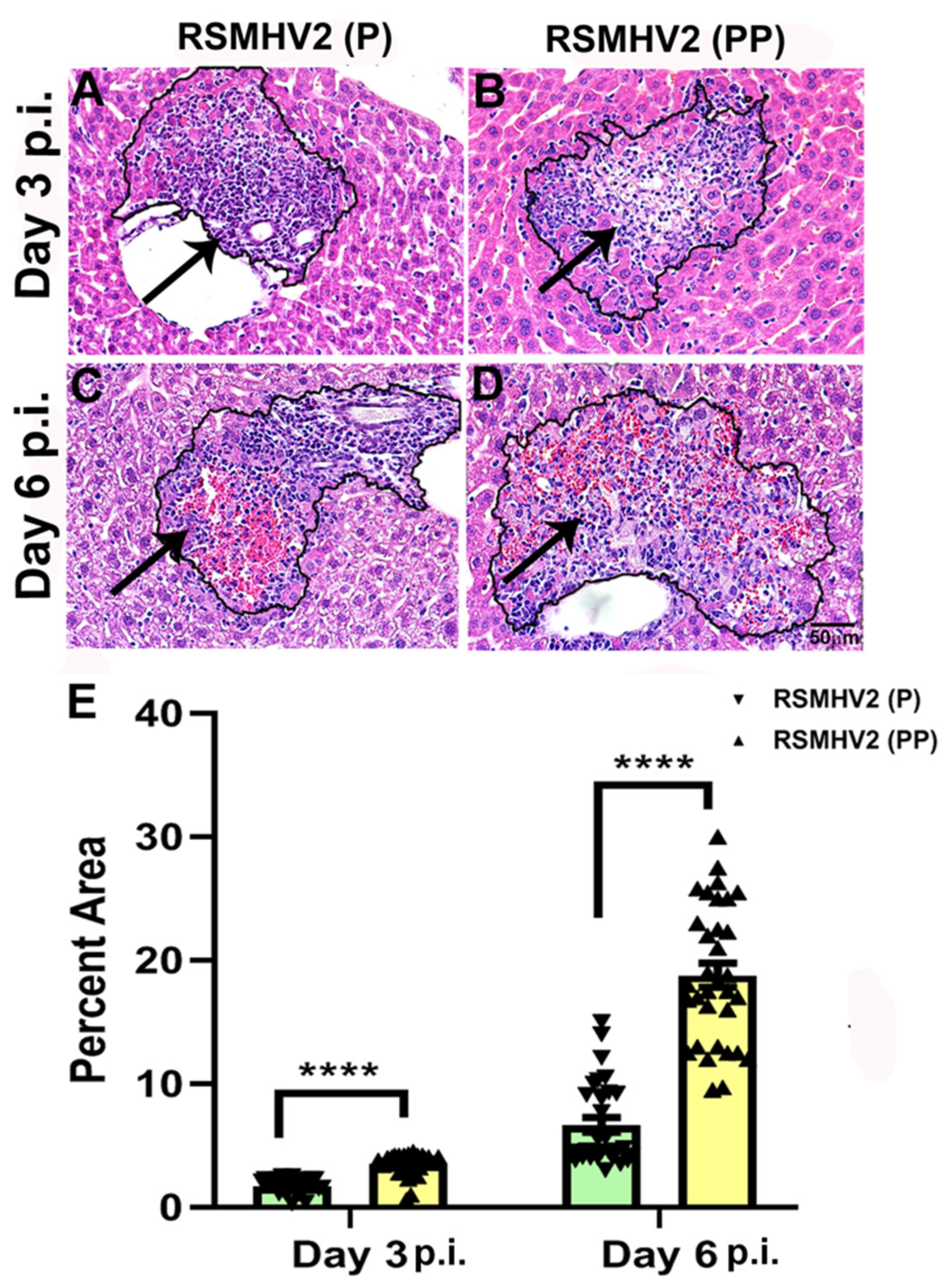
3.4. RSMHV2 (P) and RSMHV2 (PP) Differ in Their Ability to Induce Necrotizing Hepatitis at Day 3 and Day 6 p.i.
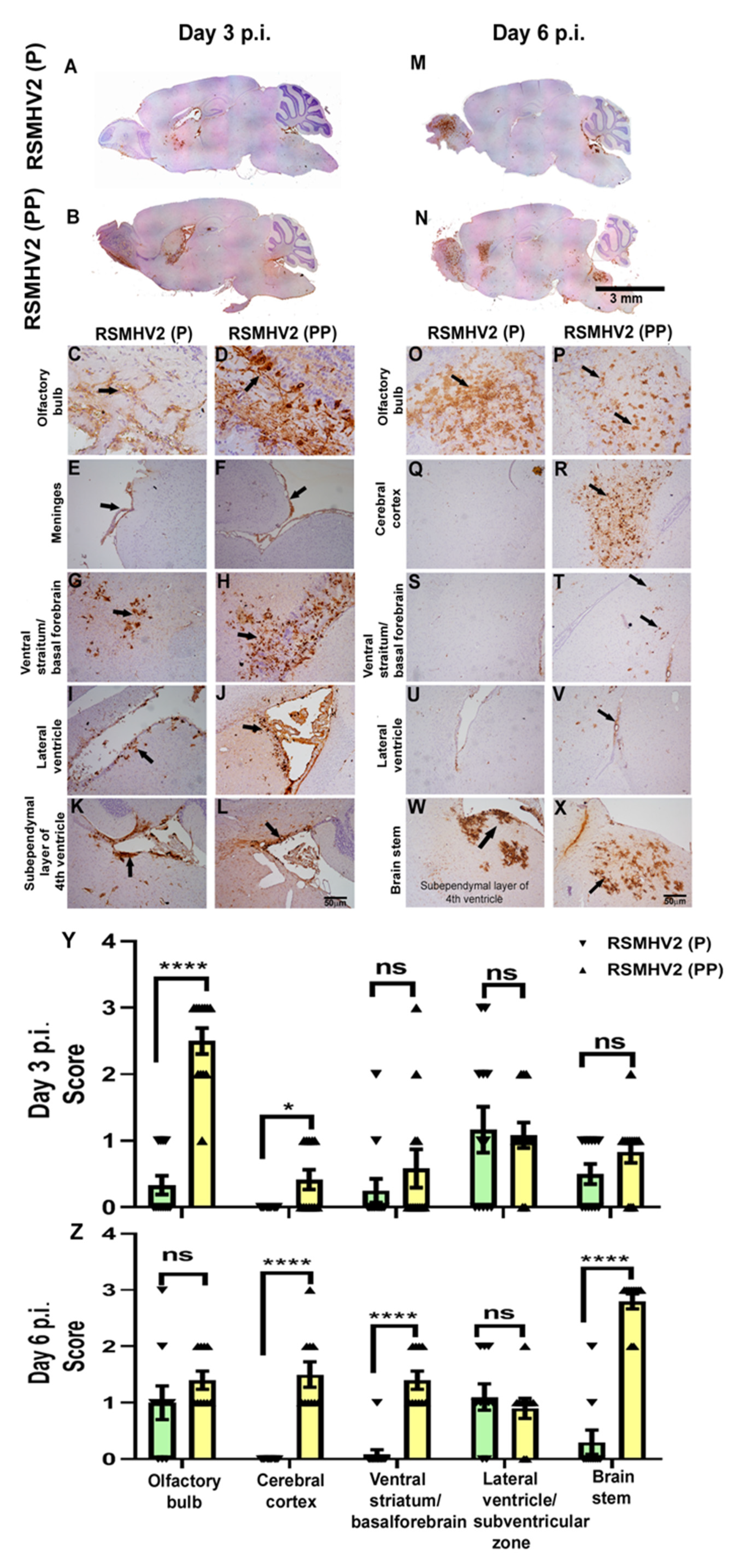
3.5. RSMHV2 (P), and RSMHV2 (PP) Differ in Their Ability to Distribute Viral Antigen in Different Neuroanatomic Regions on Day 3 and Day 6 p.i.
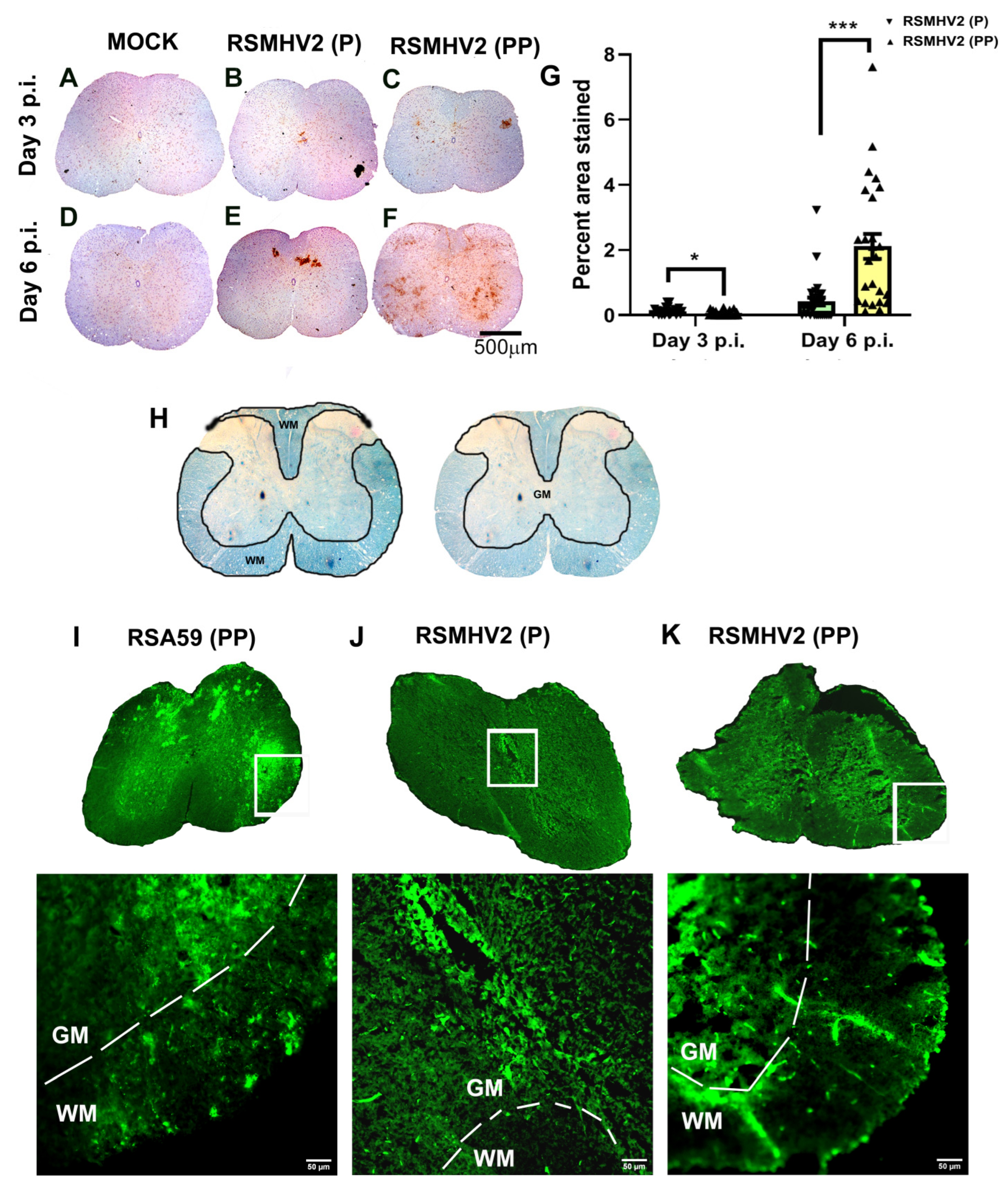
3.6. RSMHV2 (P) and RSMHV2 (PP) Differ in Their Ability to Neuronal Spread from Gray Matter to White Matter and Invade into the White Matter in Spinal Cord
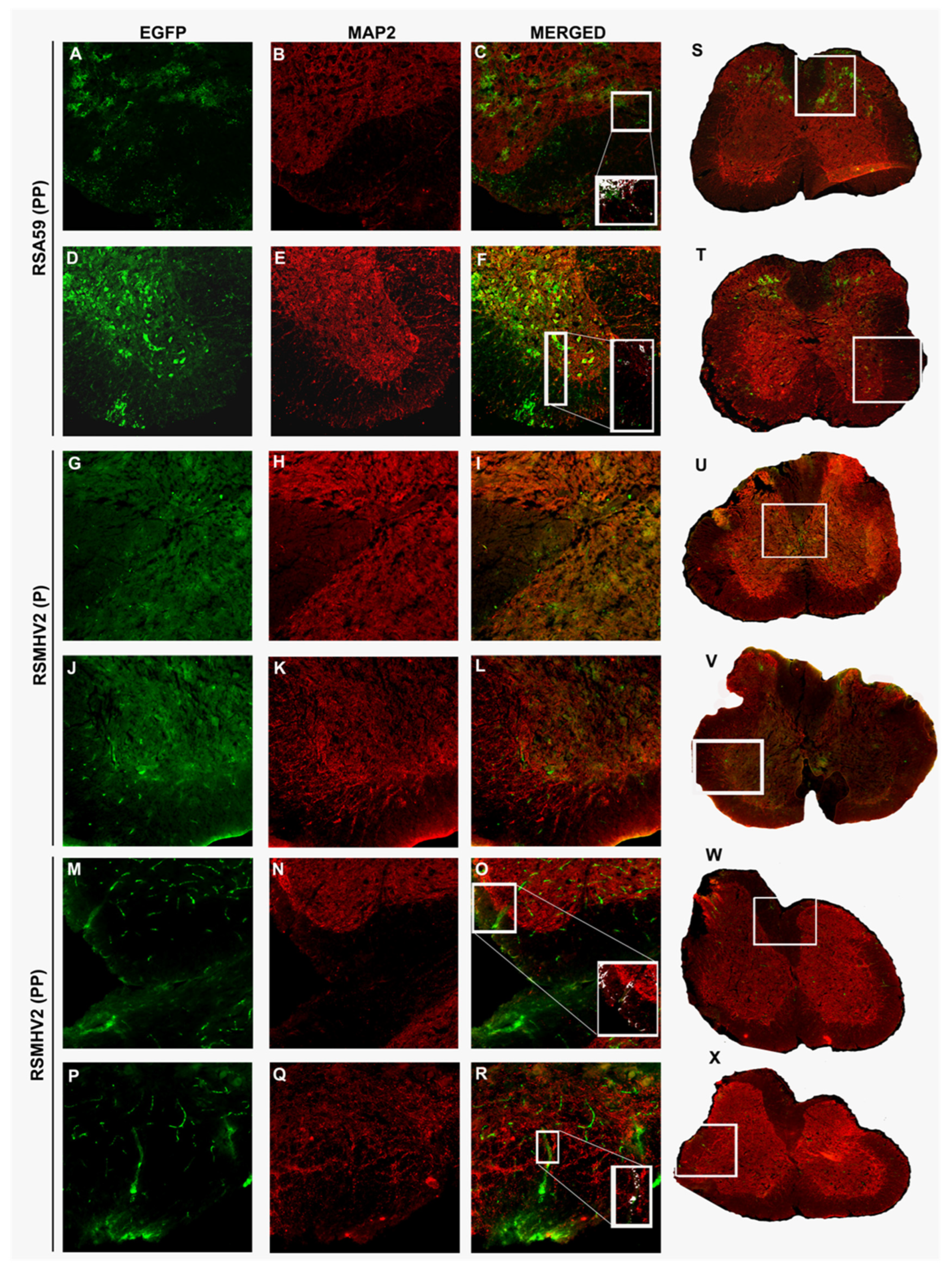
3.7. RSMHV2 (PP) and RSMHV2 (P) Differ in Their Axonal Transport Contributing to Viral Antigen Spread from Gray Matter to White Matter
3.8. RSMHV2 (PP) Induced Mild to Moderate Myelin Damage Compared to No Demyelination of RSMHV2 (P), but the Intensity of the Demyelination Was Much Less Compared to Parental Demyelinating Strain RSA59 (PP) as Studied on Day 30 p.i.
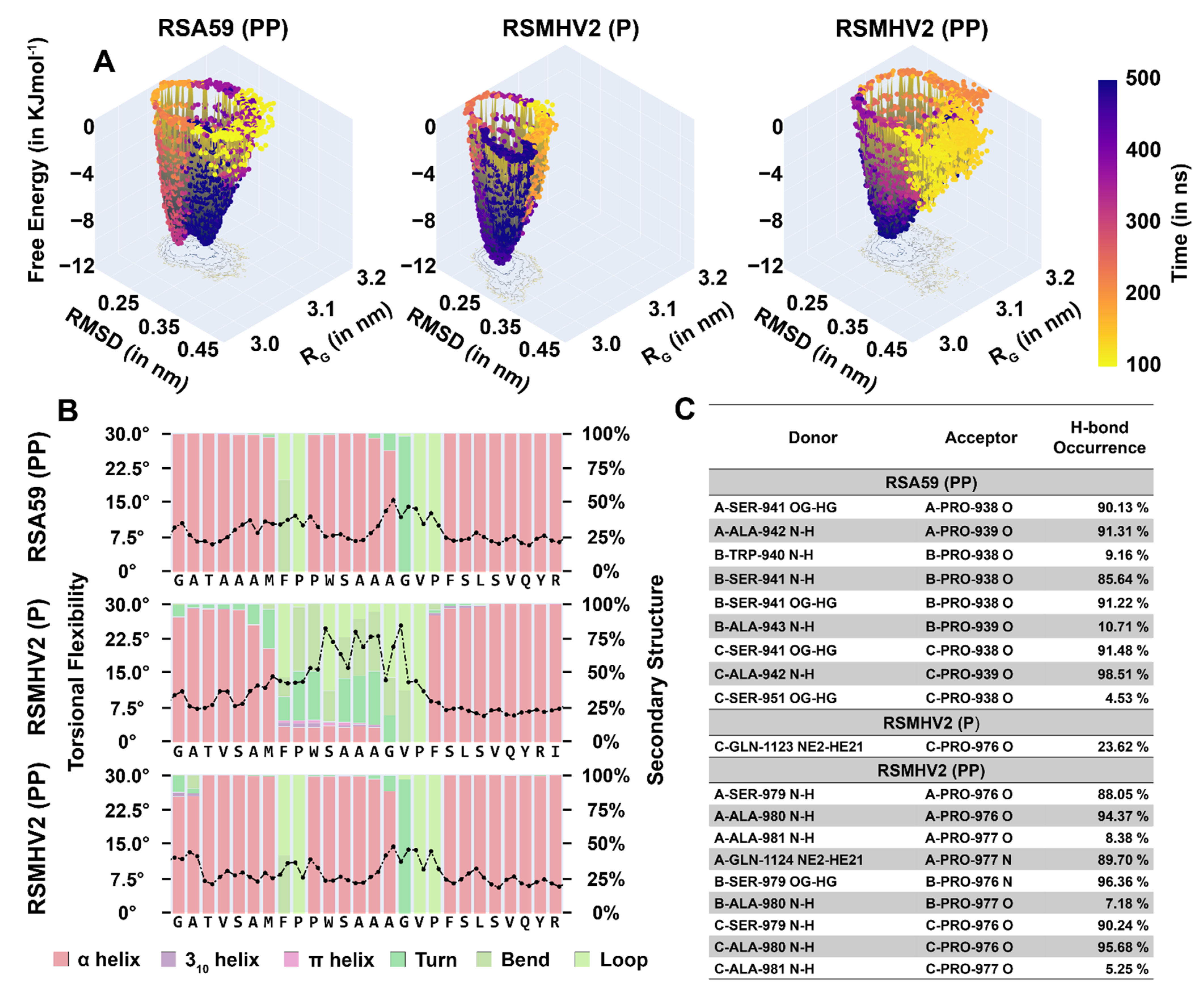
3.9. Rigidity in the Fusion Peptide from the Addition of Proline to Spike Protein from RSMHV2 (P)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Earp, L.J.; Delos, S.E.; Park, H.E.; White, J.M. The many mechanisms of viral membrane fusion proteins. Curr. Top. Microbiol. Immunol. 2005, 285, 25–66. [Google Scholar] [CrossRef] [PubMed]
- Durell, S.R.; Martin, I.; Ruysschaert, J.M.; Shai, Y.; Blumenthal, R. What studies of fusion peptides tell us about viral envelope glycoprotein-mediated membrane fusion (review). Mol. Membr. Biol. 1997, 14, 97–112. [Google Scholar] [CrossRef] [PubMed]
- Lamb, R.A.; Jardetzky, T.S. Structural basis of viral invasion: Lessons from paramyxovirus F. Curr. Opin. Struct. Biol. 2007, 17, 427–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, J.M.; Whittaker, G.R. Fusion of Enveloped Viruses in Endosomes. Traffic 2016, 17, 593–614. [Google Scholar] [CrossRef] [Green Version]
- Gallagher, T.M.; Buchmeier, M.J. Coronavirus spike proteins in viral entry and pathogenesis. Virology 2001, 279, 371–374. [Google Scholar] [CrossRef] [Green Version]
- Heald-Sargent, T.; Gallagher, T. Ready, set, fuse! The coronavirus spike protein and acquisition of fusion competence. Viruses 2012, 4, 557–580. [Google Scholar] [CrossRef] [Green Version]
- Xia, S.; Yan, L.; Xu, W.; Agrawal, A.S.; Algaissi, A.; Tseng, C.-T.K.; Wang, Q.; Du, L.; Tan, W.; Wilson, I.A.; et al. A pan-coronavirus fusion inhibitor targeting the HR1 domain of human coronavirus spike. Sci. Adv. 2019, 5, eaav4580. [Google Scholar] [CrossRef] [Green Version]
- Bosch, B.J.; van der Zee, R.; de Haan, C.A.; Rottier, P.J. The coronavirus spike protein is a class I virus fusion protein: Structural and functional characterization of the fusion core complex. J. Virol. 2003, 77, 8801–8811. [Google Scholar] [CrossRef] [Green Version]
- Kirchdoerfer, R.N.; Cottrell, C.A.; Wang, N.; Pallesen, J.; Yassine, H.M.; Turner, H.L.; Corbett, K.S.; Graham, B.S.; McLellan, J.S.; Ward, A.B. Pre-fusion structure of a human coronavirus spike protein. Nature 2016, 531, 118–121. [Google Scholar] [CrossRef] [Green Version]
- Li, F. Structure, Function, and Evolution of Coronavirus Spike Proteins. Annu. Rev. Virol. 2016, 3, 237–261. [Google Scholar] [CrossRef] [Green Version]
- Sainz, B., Jr.; Rausch, J.M.; Gallaher, W.R.; Garry, R.F.; Wimley, W.C. Identification and characterization of the putative fusion peptide of the severe acute respiratory syndrome-associated coronavirus spike protein. J. Virol. 2005, 79, 7195–7206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Z.; Hingley, S.T.; Simmons, G.; Yu, C.; Das Sarma, J.; Bates, P.; Weiss, S.R. Endosomal proteolysis by cathepsins is necessary for murine coronavirus mouse hepatitis virus type 2 spike-mediated entry. J. Virol. 2006, 80, 5768–5776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, S.Q.; Peng, H.J. Characteristics of and Public Health Responses to the Coronavirus Disease 2019 Outbreak in China. J. Clin. Med. 2020, 9, 575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hui, D.S.; Azhar, E.I.; Madani, T.A.; Ntoumi, F.; Kock, R.; Dar, O.; Ippolito, G.; McHugh, T.D.; Memish, Z.A.; Drosten, C.; et al. The continuing 2019-nCoV epidemic threat of novel coronaviruses to global health—The latest 2019 novel coronavirus outbreak in Wuhan, China. Int. J. Infect. Dis. 2020, 91, 264–266. [Google Scholar] [CrossRef] [Green Version]
- Tang, D.; Comish, P.; Kang, R. The hallmarks of COVID-19 disease. PLoS Pathog. 2020, 16, e1008536. [Google Scholar] [CrossRef]
- Rout, S.S.; Singh, M.; Shindler, K.S.; Das Sarma, J. One proline deletion in the fusion peptide of neurotropic mouse hepatitis virus (MHV) restricts retrograde axonal transport and neurodegeneration. J. Biol. Chem. 2020, 295, 6926–6935. [Google Scholar] [CrossRef] [Green Version]
- Singh, M.; Kishore, A.; Maity, D.; Sunanda, P.; Krishnarjuna, B.; Vappala, S.; Raghothama, S.; Kenyon, L.C.; Pal, D.; Das Sarma, J. A proline insertion-deletion in the spike glycoprotein fusion peptide of mouse hepatitis virus strongly alters neuropathology. J. Biol. Chem. 2019, 294, 8064–8087. [Google Scholar] [CrossRef] [Green Version]
- Das Sarma, J.; Fu, L.; Hingley, S.T.; Lai, M.M.; Lavi, E. Sequence analysis of the S gene of recombinant MHV-2/A59 coronaviruses reveals three candidate mutations associated with demyelination and hepatitis. J. Neurovirol. 2001, 7, 432–436. [Google Scholar] [CrossRef]
- Das Sarma, J.; Fu, L.; Hingley, S.T.; Lavi, E. Mouse hepatitis virus type-2 infection in mice: An experimental model system of acute meningitis and hepatitis. Exp. Mol. Pathol. 2001, 71, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Navas, S.; Seo, S.H.; Chua, M.M.; Das Sarma, J.; Lavi, E.; Hingley, S.T.; Weiss, S.R. Murine coronavirus spike protein determines the ability of the virus to replicate in the liver and cause hepatitis. J. Virol. 2001, 75, 2452–2457. [Google Scholar] [CrossRef] [Green Version]
- Das Sarma, J.; Fu, L.; Tsai, J.C.; Weiss, S.R.; Lavi, E. Demyelination determinants map to the spike glycoprotein gene of coronavirus mouse hepatitis virus. J. Virol. 2000, 74, 9206–9213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavi, E.; Gilden, D.H.; Wroblewska, Z.; Rorke, L.B.; Weiss, S.R. Experimental demyelination produced by the A59 strain of mouse hepatitis virus. Neurology 1984, 34, 597. [Google Scholar] [CrossRef] [PubMed]
- Perlman, S.; Jacobsen, G.; Olson, A.L.; Afifi, A. Identification of the spinal cord as a major site of persistence during chronic infection with a murine coronavirus. Virology 1990, 175, 418–426. [Google Scholar] [CrossRef]
- Shindler, K.S.; Kenyon, L.C.; Dutt, M.; Hingley, S.T.; Das Sarma, J. Experimental optic neuritis induced by a demyelinating strain of mouse hepatitis virus. J. Virol. 2008, 82, 8882–8886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das Sarma, J.; Scheen, E.; Seo, S.H.; Koval, M.; Weiss, S.R. Enhanced green fluorescent protein expression may be used to monitor murine coronavirus spread in vitro and in the mouse central nervous system. J. Neurovirol. 2002, 8, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Das Sarma, J.; Kenyon, L.C.; Hingley, S.T.; Shindler, K.S. Mechanisms of primary axonal damage in a viral model of multiple sclerosis. J. Neurosci. 2009, 29, 10272–10280. [Google Scholar] [CrossRef]
- Kenyon, L.C.; Biswas, K.; Shindler, K.S.; Nabar, M.; Stout, M.; Hingley, S.T.; Grinspan, J.B.; Das Sarma, J. Gliopathy of Demyelinating and Non-Demyelinating Strains of Mouse Hepatitis Virus. Front. Cell Neurosci. 2015, 9, 488. [Google Scholar] [CrossRef] [Green Version]
- Shindler, K.S.; Chatterjee, D.; Biswas, K.; Goyal, A.; Dutt, M.; Nassrallah, M.; Khan, R.S.; Das Sarma, J. Macrophage-mediated optic neuritis induced by retrograde axonal transport of spike gene recombinant mouse hepatitis virus. J. Neuropathol. Exp. Neurol. 2011, 70, 470–480. [Google Scholar] [CrossRef]
- Sadasivan, J.; Singh, M.; Sarma, J.D. Cytoplasmic tail of coronavirus spike protein has intracellular targeting signals. J. Biosci. 2017, 42, 231–244. [Google Scholar] [CrossRef]
- Das Sarma, J.; Iacono, K.; Gard, L.; Marek, R.; Kenyon, L.C.; Koval, M.; Weiss, S.R. Demyelinating and nondemyelinating strains of mouse hepatitis virus differ in their neural cell tropism. J. Virol. 2008, 82, 5519–5526. [Google Scholar] [CrossRef] [Green Version]
- Kuo, L.; Godeke, G.J.; Raamsman, M.J.; Masters, P.S.; Rottier, P.J. Retargeting of coronavirus by substitution of the spike glycoprotein ectodomain: Crossing the host cell species barrier. J. Virol. 2000, 74, 1393–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donnelly, D.J.; Gensel, J.C.; Ankeny, D.P.; van Rooijen, N.; Popovich, P.G. An efficient and reproducible method for quantifying macrophages in different experimental models of central nervous system pathology. J. Neurosci. Methods 2009, 181, 36–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bose, A.; Basu, R.; Maulik, M.; Das Sarma, J. Loss of Cx43-Mediated Functional Gap Junction Communication in Meningeal Fibroblasts Following Mouse Hepatitis Virus Infection. Mol. Neurobiol. 2018, 55, 6558–6571. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, P.; Pal, D. The interrelationships of side-chain and main-chain conformations in proteins. Prog. Biophys. Mol. Biol. 2001, 76, 1–102. [Google Scholar] [CrossRef]
- Sali, A.; Blundell, T.L. Comparative protein modelling by satisfaction of spatial restraints. J. Mol. Biol. 1993, 234, 779–815. [Google Scholar] [CrossRef] [PubMed]
- Parrinello, M.; Rahman, A. Polymorphic transitions in single crystals: A new molecular dynamics method. J. Appl. Phys. 1981, 52, 7182–7190. [Google Scholar] [CrossRef]
- Berendsen, H.J.C.; Postma, J.P.M.; Van Gunsteren, W.F.; DiNola, A.; Haak, J.R. Molecular dynamics with coupling to an external bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar] [CrossRef] [Green Version]
- Tavernelli, I.; Cotesta, S.; Di Iorio, E.E. Protein Dynamics, Thermal Stability, and Free-Energy Landscapes: A Molecular Dynamics Investigation. Biophys. J. 2003, 85, 2641–2649. [Google Scholar] [CrossRef] [Green Version]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Safiriyu, A.A.; Singh, M.; Kishore, A.; Mulchandani, V.; Maity, D.; Behera, A.; Sinha, B.; Pal, D.; Das Sarma, J. Two Consecutive Prolines in the Fusion Peptide of Murine β-Coronavirus Spike Protein Predominantly Determine Fusogenicity and May Be Essential but Not Sufficient to Cause Demyelination. Viruses 2022, 14, 834. https://doi.org/10.3390/v14040834
Safiriyu AA, Singh M, Kishore A, Mulchandani V, Maity D, Behera A, Sinha B, Pal D, Das Sarma J. Two Consecutive Prolines in the Fusion Peptide of Murine β-Coronavirus Spike Protein Predominantly Determine Fusogenicity and May Be Essential but Not Sufficient to Cause Demyelination. Viruses. 2022; 14(4):834. https://doi.org/10.3390/v14040834
Chicago/Turabian StyleSafiriyu, Abass Alao, Manmeet Singh, Abhinoy Kishore, Vaishali Mulchandani, Dibyajyoti Maity, Amrutamaya Behera, Bidisha Sinha, Debnath Pal, and Jayasri Das Sarma. 2022. "Two Consecutive Prolines in the Fusion Peptide of Murine β-Coronavirus Spike Protein Predominantly Determine Fusogenicity and May Be Essential but Not Sufficient to Cause Demyelination" Viruses 14, no. 4: 834. https://doi.org/10.3390/v14040834