
Development of Robust Varicella Zoster Virus Luciferase Reporter Viruses for In Vivo Monitoring of Virus Growth and Its Antiviral Inhibition in Culture, Skin, and Humanized Mice


 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.1.1. Generation of Luciferase Reporter Viruses
2.1.2. Correction of Spurious Mutations
2.1.3. Deletion of ORF13 and ORF36 in the BAC DXRR 57luczeo Background
2.2. Compounds
2.3. Virus Growth Kinetics in Cells
2.4. Total Flux Half-Life in Cells Infected with VZV-ORFx-Luc Viruses
2.5. Efficacy in Cultured Cells
2.6. Preparation of Skin
2.7. Skin Organ Culture
2.8. Animal Procedures
2.9. Bioluminescence Imaging
2.10. Statistical Analysis
3. Results
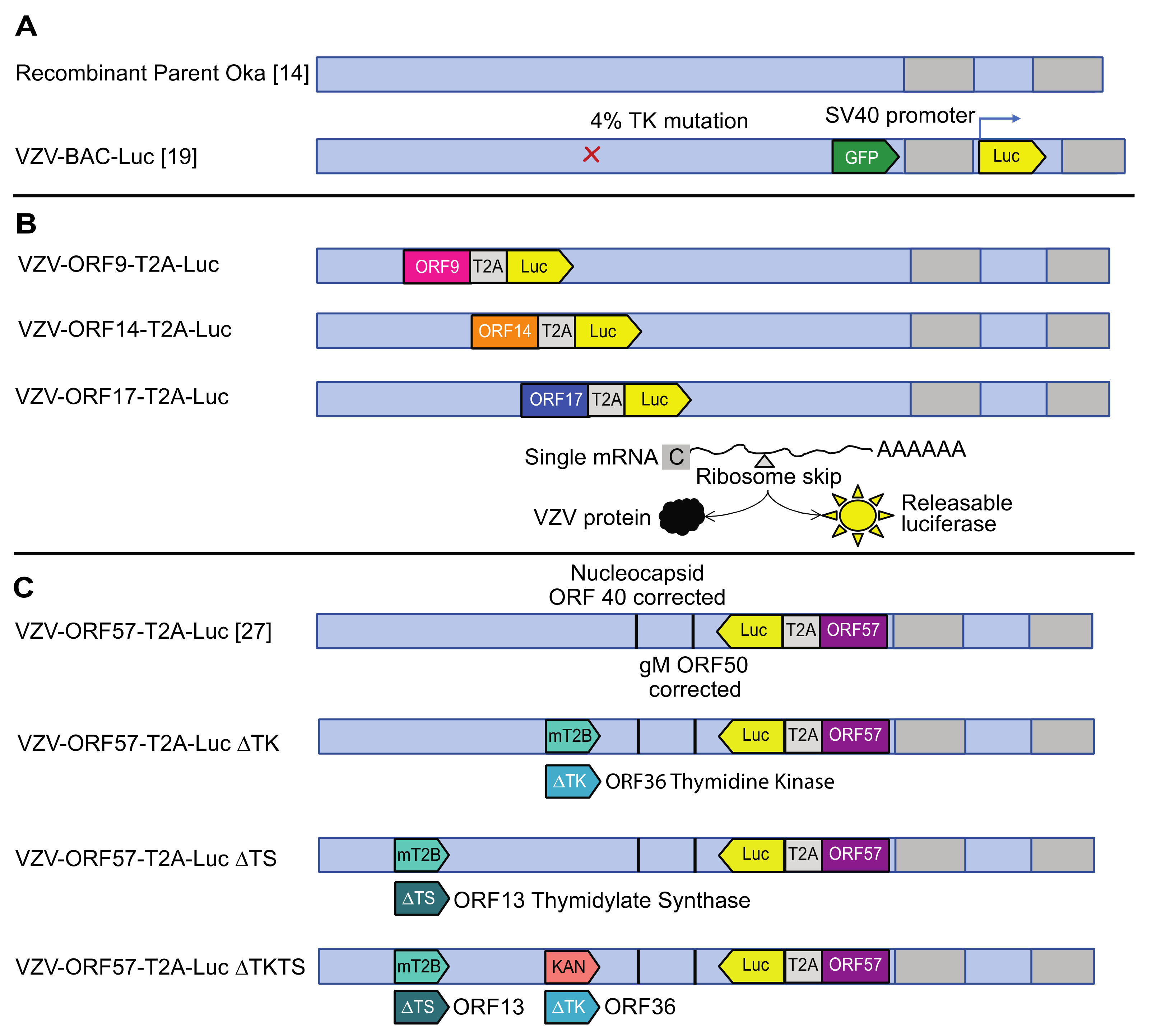
3.1. Construction of Robust Reporter VZV Viruses
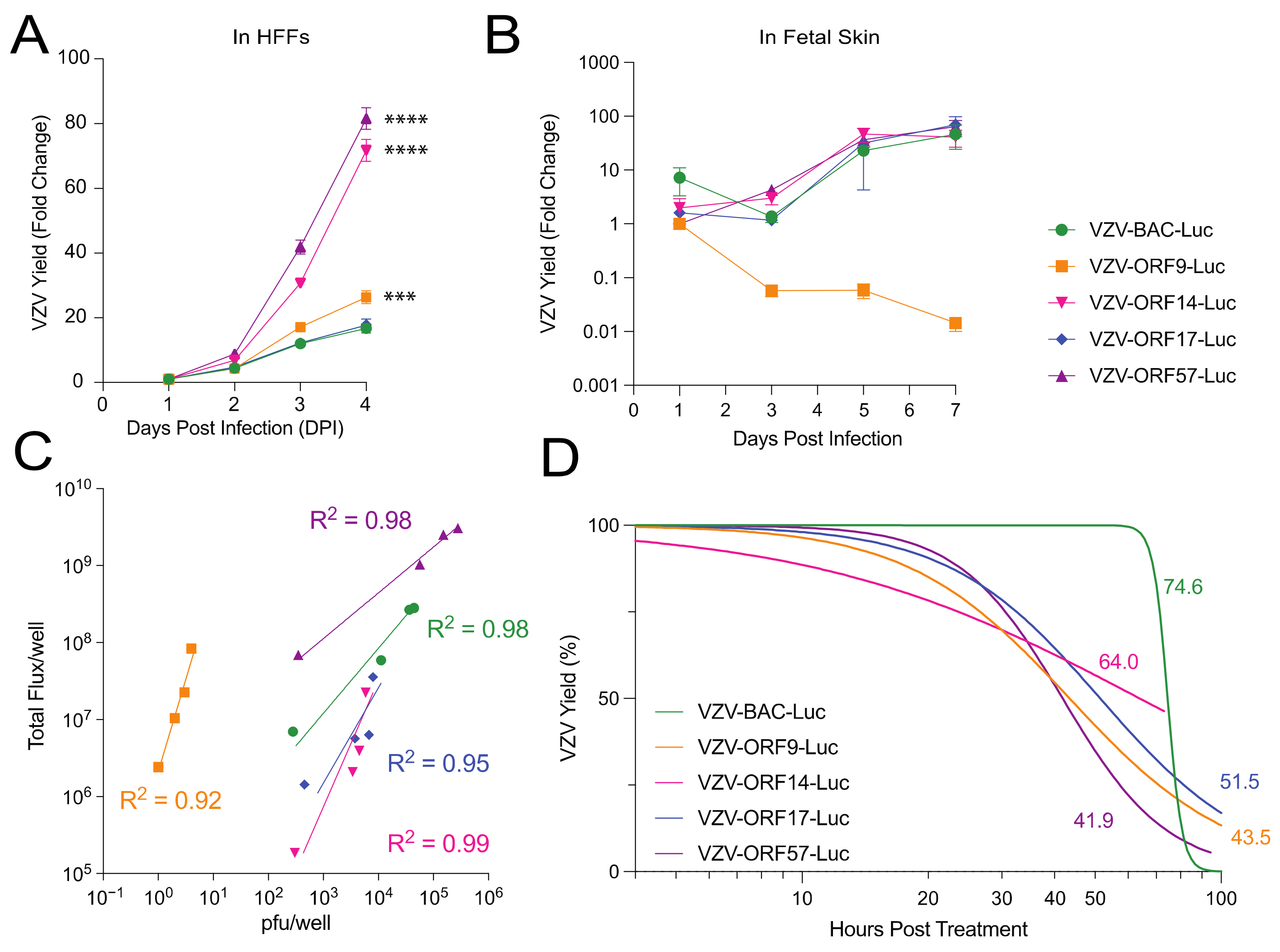
3.2. Comparison of VZV-BAC-Luc to VZV-ORFx-Luc Reporter Viruses in Cells and Skin
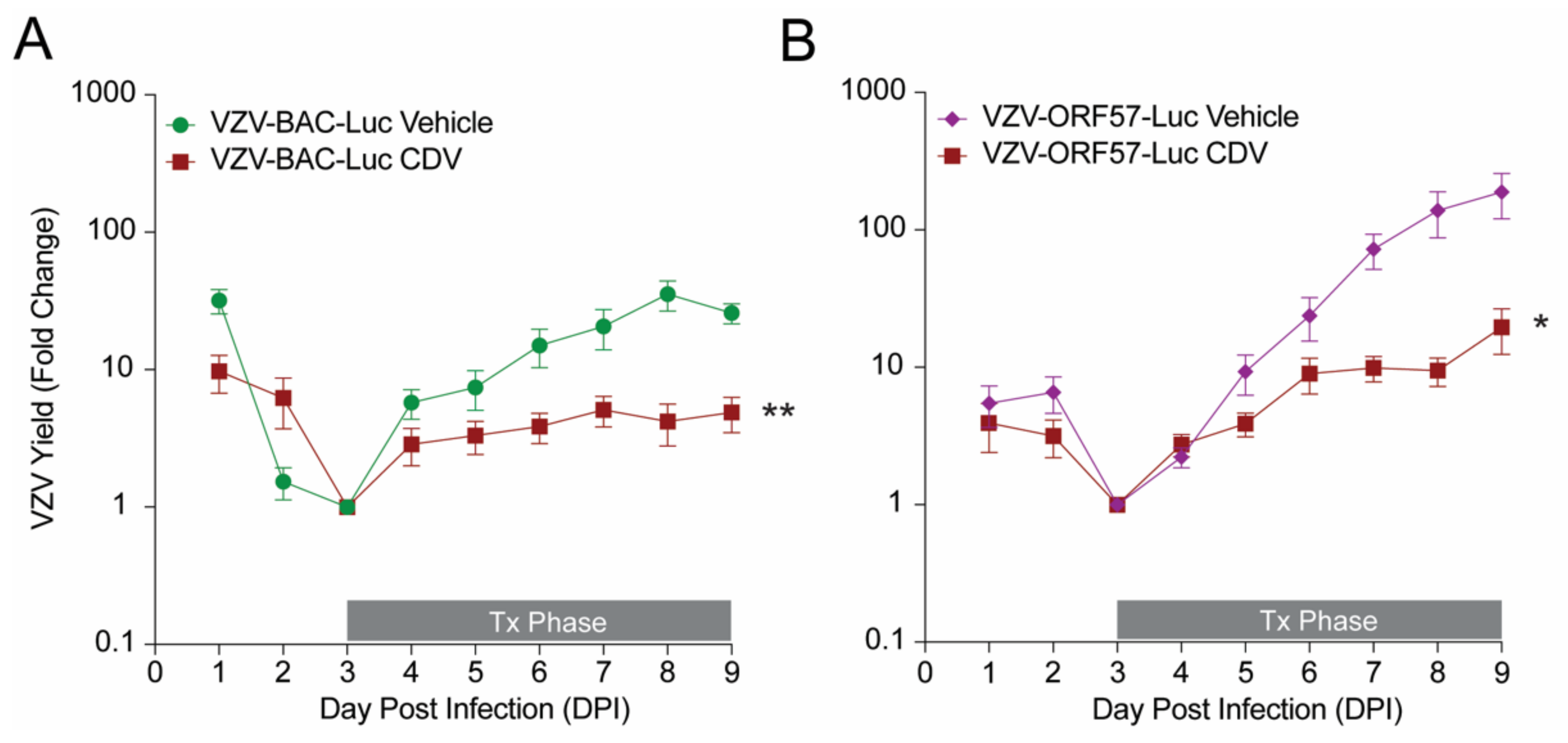
3.3. Comparison of VZV-BAC-Luc and VZV-ORF57-Luc in a SCIDhu Mouse Model
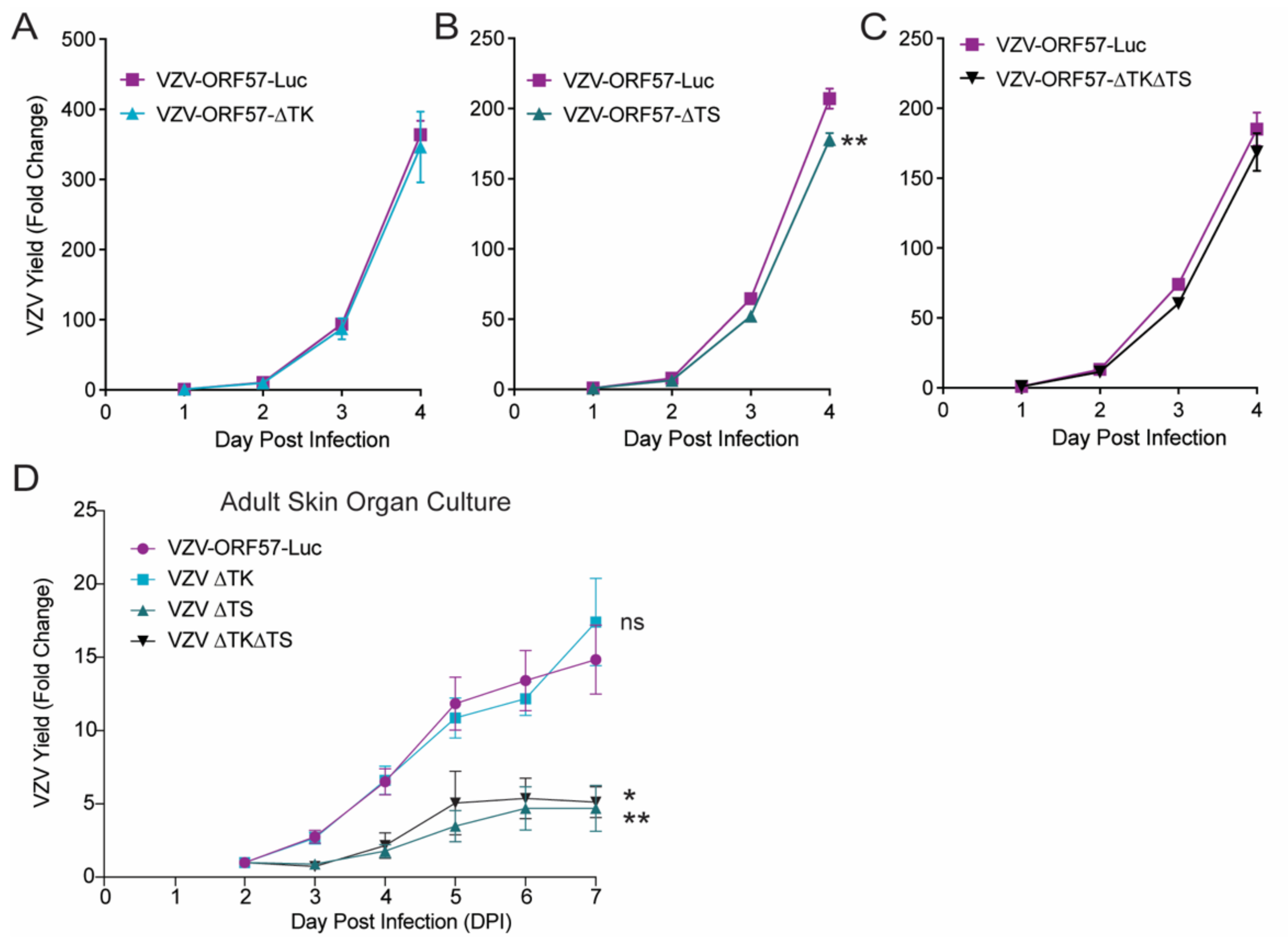
3.4. VZV Thymidylate Synthase (TS), but Not Thymidine Kinase (TK), Is Required for Virulence
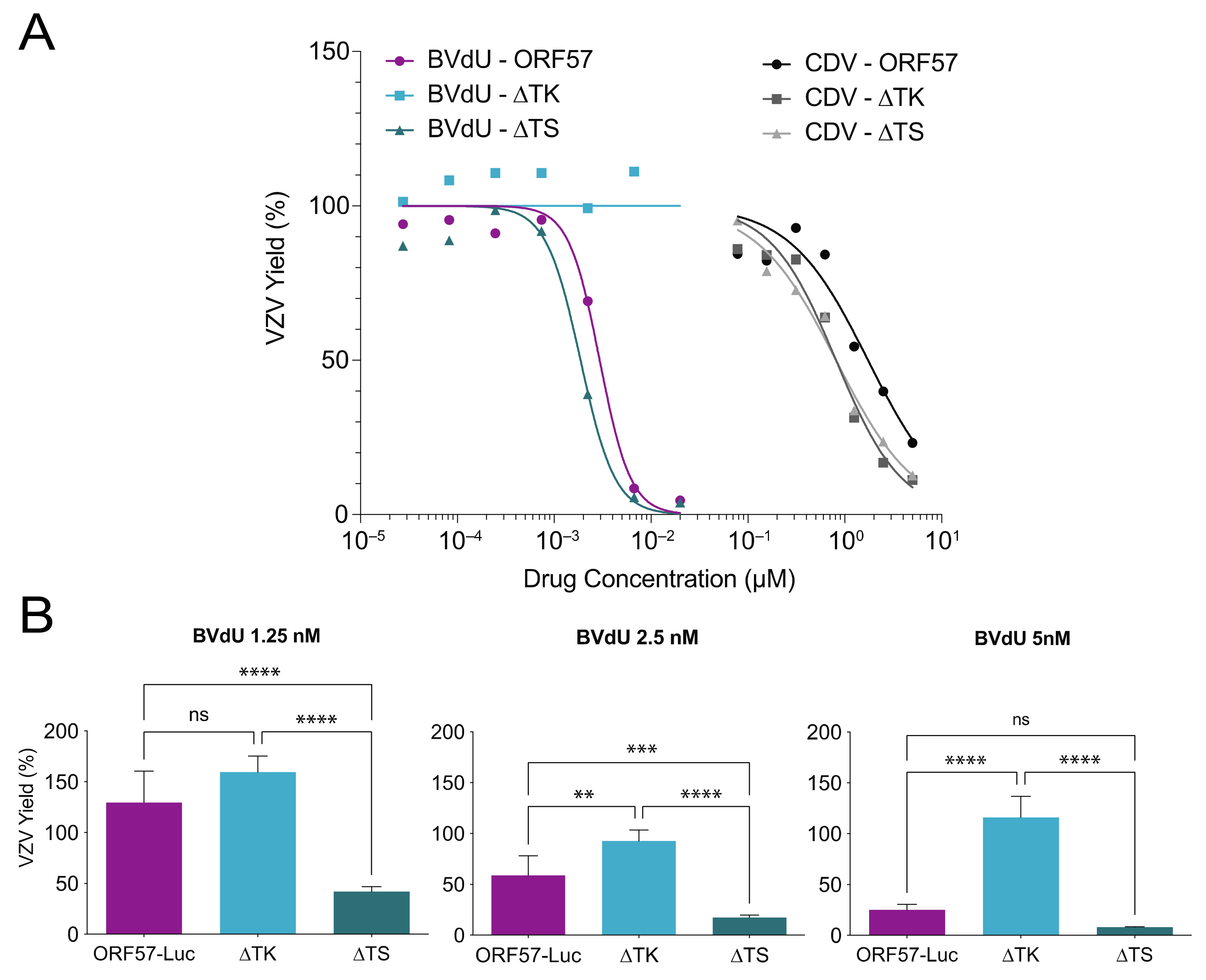
3.5. VZV TS Deletion Increases Sensitivity to Antiviral Drugs
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lopez, A.S.; Zhang, J.; Marin, M. Epidemiology of Varicella During the 2-Dose Varicella Vaccination Program—United States, 2005–2014. MMWR. Morb. Mortal. Wkly. Rep. 2016, 65, 902–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gershon, A.A.; Gershon, M.D. Pathogenesis and Current Approaches to Control of Varicella-Zoster Virus Infections. Clin. Microbiol. Rev. 2013, 26, 728–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Draper, M.; Stergiopoulos, S. Shingles vaccination uptake in Massachusetts adults aged 50 years and older. Vaccine 2021, 39, 6781–6786. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.-J.; Hung, M.-C.; Srivastav, A.; Grohskopf, L.A.; Kobayashi, M.; Harris, A.M.; Dooling, K.L.; Markowitz, L.E.; Rodriguez-Lainz, A.; Williams, W.W. Surveillance of Vaccination Coverage Among Adult Populations—United States. MMWR. Surveill. Summ. 2021, 70, 1–26. [Google Scholar] [CrossRef]
- Patterson, B.J.; Chen, C.-C.; McGuiness, C.B.; Glasser, L.I.; Sun, K.; Buck, P.O. Early examination of real-world uptake and second-dose completion of recombinant zoster vaccine in the United States from October 2017 to September. Hum. Vaccines Immunother. 2021, 17, 2482–2487. [Google Scholar] [CrossRef]
- De Clercq, E.; Li, G. Approved Antiviral Drugs over the Past 50 Years. Clin. Microbiol. Rev. 2016, 29, 695–747. [Google Scholar] [CrossRef] [Green Version]
- Poole, C.L.; James, S.H. Antiviral Therapies for Herpesviruses: Current Agents and New Directions. Clin. Ther. 2018, 40, 1282–1298. [Google Scholar] [CrossRef] [Green Version]
- De Clercq, E. Fifty Years in Search of Selective Antiviral Drugs. J. Med. Chem. 2019, 62, 7322–7339. [Google Scholar] [CrossRef]
- Mahalingam, R.; Gershon, A.; Gershon, M.; Cohen, J.I.; Arvin, A.; Zerboni, L.; Zhu, H.; Gray, W.; Messaoudi, I.; Traina-Dorge, V. Current In Vivo Models of Varicella-Zoster Virus Neurotropism. Viruses 2019, 11, 502. [Google Scholar] [CrossRef] [Green Version]
- Haberthur, K.; Messaoudi, I. Animal Models of Varicella Zoster Virus Infection. Pathogens 2013, 2, 364–382. [Google Scholar] [CrossRef]
- Roizman, B.; Knipe, D.M.; Whitley, R.J. (Eds.) Fields Virology, Chapter 67: Herpes Simplex Viruses, 5th ed.; Lippincott Williams & Wilkin: Philadelphia, PA, USA, 2007; pp. 2502–2601. [Google Scholar]
- Cohen, J.I.; Seidel, K.E. Generation of varicella-zoster virus (VZV) and viral mutants from cosmid DNAs: VZV thymidylate synthetase is not essential for replication in vitro. Proc. Natl. Acad. Sci. USA 1993, 90, 7376–7380. [Google Scholar] [CrossRef] [Green Version]
- Sawyer, M.H.; Inchauspe, G.; Biron, K.K.; Waters, D.J.; Straus, S.E.; Ostrove, J.M. Molecular analysis of the pyrimidine deoxyribonucleoside kinase gene of wild-type and acyclovir-resistant strains of varicella-zoster virus. J. Gen. Virol. 1988, 69, 2585–2593. [Google Scholar] [CrossRef]
- Niizuma, T.; Zerboni, L.; Sommer, M.H.; Ito, H.; Hinchliffe, S.; Arvin, A.M. Construction of varicella-zoster virus recombinants from parent Oka cosmids and demonstration that ORF65 protein is dispensable for infection of human skin and T cells in the SCID-hu mouse model. J. Virol. 2003, 77, 6062–6065. [Google Scholar] [CrossRef] [Green Version]
- Zerboni, L.; Hinchliffe, S.; Sommer, M.H.; Ito, H.; Besser, J.; Stamatis, S.; Cheng, J.; DiStefano, D.; Kraiouchkine, N.; Shaw, A.; et al. Analysis of varicella zoster virus attenuation by evaluation of chimeric parent Oka/vaccine Oka recombinant viruses in skin xenografts in the SCIDhu mouse model. Virology 2005, 332, 337–346. [Google Scholar] [CrossRef] [Green Version]
- Nagaike, K.; Mori, Y.; Gomi, Y.; Yoshii, H.; Takahashi, M.; Wagner, M.; Koszinowski, U.; Yamanishi, K. Cloning of the varicella-zoster virus genome as an infectious bacterial artificial chromosome in Escherichia coli. Vaccine 2004, 22, 4069–4074. [Google Scholar] [CrossRef]
- Yoshii, H.; Somboonthum, P.; Takahashi, M.; Yamanishi, K.; Mori, Y. Cloning of full length genome of varicella-zoster virus vaccine strain into a bacterial artificial chromosome and reconstitution of infectious virus. Vaccine 2007, 25, 5006–5012. [Google Scholar] [CrossRef]
- Tischer, B.K.; Kaufer, B.B.; Sommer, M.; Wussow, F.; Arvin, A.M.; Osterrieder, N. A Self-Excisable Infectious Bacterial Artificial Chromosome Clone of Varicella-Zoster Virus Allows Analysis of the Essential Tegument Protein Encoded by ORF9. J. Virol. 2007, 81, 13200–13208. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Rowe, J.; Wang, W.; Sommer, M.; Arvin, A.; Moffat, J.; Zhu, H. Genetic Analysis of Varicella-Zoster Virus ORF0 to ORF4 by Use of a Novel Luciferase Bacterial Artificial Chromosome System. J. Virol. 2007, 81, 9024–9033. [Google Scholar] [CrossRef] [Green Version]
- Markus, A.; Grigoryan, S.; Sloutskin, A.; Yee, M.B.; Zhu, H.; Yang, I.H.; Thakor, N.V.; Sarid, R.; Kinchington, P.R.; Goldstein, R.S. Varicella-Zoster Virus (VZV) Infection of Neurons Derived from Human Embryonic Stem Cells: Direct Demonstration of Axonal Infection, Transport of VZV, and Productive Neuronal Infection. J. Virol. 2011, 85, 6220–6233. [Google Scholar] [CrossRef] [Green Version]
- Tischer, B.K.; von Einem, J.; Kaufer, B.; Osterrieder, N. Two-step red-mediated recombination for versatile high-efficiency markerless DNA manipulation in Escherichia coli. Biotechniques 2006, 40, 191–197. [Google Scholar]
- Zhang, Z.; Huang, Y.; Zhu, H. An Efficient Protocol for VZV BAC-Based Mutagenesis. Adv. Struct. Saf. Stud. 2010, 634, 75–86. [Google Scholar] [CrossRef]
- Depledge, D.P.; Sadaoka, T.; Ouwendijk, W.J.D. Molecular Aspects of Varicella-Zoster Virus Latency. Viruses 2018, 10, 349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Selariu, A.; Warden, C.; Huang, G.; Huang, Y.; Zaccheus, O.; Cheng, T.; Xia, N.; Zhu, H. Genome-wide mutagenesis reveals that ORF7 is a novel VZV skin-tropic factor. PLoS Pathog. 2010, 6, e1000971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luker, G.D.; Prior, J.L.; Song, J.; Pica, C.M.; Leib, D.A. Bioluminescence Imaging Reveals Systemic Dissemination of Herpes Simplex Virus Type 1 in the Absence of Interferon Receptors. J. Virol. 2003, 77, 11082–11093. [Google Scholar] [CrossRef] [Green Version]
- Rowe, J.; Greenblatt, R.J.; Liu, D.; Moffat, J.F. Compounds that target host cell proteins prevent varicella-zoster virus replication in culture, ex vivo, and in SCID-Hu mice. Antivir. Res. 2010, 86, 276–285. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, M.G.; Smith, N.A.; Tighe, M.; Travis, K.L.; Liu, D.; Upadhyaya, P.K.; Kinchington, P.R.; Chan, G.C.; Moffat, J.F. A Novel Human Skin Tissue Model to Study Varicella-Zoster Virus and Human Cytomegalovirus. J. Virol. 2020, 94, 22. [Google Scholar] [CrossRef]
- Erazo, A.; Yee, M.B.; Osterrieder, N.; Kinchington, P.R. Varicella-Zoster Virus Open Reading Frame 66 Protein Kinase Is Required for Efficient Viral Growth in Primary Human Corneal Stromal Fibroblast Cells. J. Virol. 2008, 82, 7653–7665. [Google Scholar] [CrossRef] [Green Version]
- Warner, B.E.; Yee, M.B.; Zhang, M.; Hornung, R.S.; Kaufer, B.B.; Visalli, R.J.; Kramer, P.R.; Goins, W.F.; Kinchington, P.R. Varicella-zoster virus early infection but not complete replication is required for the induction of chronic hypersen-sitivity in rat models of postherpetic neuralgia. PLoS Pathog. 2021, 17, e1009689. [Google Scholar] [CrossRef]
- Leisenfelder, S.A.; Moffat, J.F. Varicella-Zoster Virus Infection of Human Foreskin Fibroblast Cells Results in Atypical Cyclin Expression and Cyclin-Dependent Kinase Activity. J. Virol. 2006, 80, 5577–5587. [Google Scholar] [CrossRef] [Green Version]
- De, C.; Liu, D.; Zheng, B.; Singh, U.S.; Chavre, S.; White, C.; Arnold, R.D.; Hagen, F.K.; Chu, C.K.; Moffat, J.F. β-L-1-[5-(E-2-bromovinyl)-2-(hydroxymethyl)-1,3-(dioxolan-4-yl)] uracil (l-BHDU) prevents varicella-zoster virus replication in a SCID-Hu mouse model and does not interfere with 5-fluorouracil catabolism. Antivir. Res. 2014, 110, 10–19. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, M.G.; Liu, D.; Legendre, M.; Markovitz, D.M.; Moffat, J.F. H84T BanLec has broad spectrum antiviral activity against human herpesviruses in cells, skin, and mice. Sci. Rep. 2022, 12, 1641. [Google Scholar] [CrossRef]
- Taylor, S.L.; Moffat, J.F. Replication of Varicella-Zoster Virus in Human Skin Organ Culture. J. Virol. 2005, 79, 11501–11506. [Google Scholar] [CrossRef] [Green Version]
- Moffat, J.F.; Zerboni, L.; Kinchington, P.R.; Grose, C.; Kaneshima, H.; Arvin, A.M. Attenuation of the vaccine Oka strain of varicella-zoster virus and role of glycoprotein C in alphaherpesvirus virulence demonstrated in the SCID-hu mouse. J. Virol. 1998, 72, 965–974. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, M.; Liu, D.; Lyu, J.; Fan, J.; Overhulse, J.; Kashemirov, B.; Prichard, M.; McKenna, C.; Moffat, J. An acyclic phosphonate prodrug of HPMPC is effective against VZV in skin organ culture and mice. Antivir. Res. 2022, 199, 105275. [Google Scholar] [CrossRef]
- Moffat, J.F.; Zerboni, L.; Sommer, M.H.; Heineman, T.C.; Cohen, J.I.; Kaneshima, H.; Arvin, A.M. The ORF47 and ORF66 putative protein kinases of varicella-zoster virus determine tropism for human T cells and skin in the SCID-hu mouse. Proc. Natl. Acad. Sci. USA 1998, 95, 11969–11974. [Google Scholar] [CrossRef] [Green Version]
- Besser, J.; Sommer, M.H.; Zerboni, L.; Bagowski, C.P.; Ito, H.; Moffat, J.; Ku, C.C.; Arvin, A.M. Differentiation of vari-cella-zoster virus ORF47 protein kinase and IE62 protein binding domains and their contributions to replication in human skin xenografts in the SCID-hu mouse. J. Virol. 2003, 77, 5964–5974. [Google Scholar] [CrossRef] [Green Version]
- Che, X.; Reichelt, M.; Sommer, M.H.; Rajamani, J.; Zerboni, L.; Arvin, A.M. Functions of the ORF9-to-ORF12 gene clus-ter in varicella-zoster virus replication and in the pathogenesis of skin infection. J. Virol. 2008, 82, 5825–5834. [Google Scholar] [CrossRef] [Green Version]
- De Clercq, E. Antiviral drugs in current clinical use. J. Clin. Virol. 2004, 30, 115–133. [Google Scholar] [CrossRef]
- Balzarini, J.; McGuigan, C. Chemotherapy of varicella-zoster virus by a novel class of highly specific anti-VZV bicyclic pyrimidine nucleosides. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2002, 1587, 287–295. [Google Scholar] [CrossRef] [Green Version]
- Hew, K.; Dahlroth, S.-L.; Veerappan, S.; Pan, L.X.; Cornvik, T.; Nordlund, P. Structure of the Varicella Zoster Virus Thymidylate Synthase Establishes Functional and Structural Similarities as the Human Enzyme and Potentiates Itself as a Target of Brivudine. PLoS ONE 2015, 10, e0143947. [Google Scholar] [CrossRef] [Green Version]
- Andrei, G.; Topalis, D.; Fiten, P.; McGuigan, C.; Balzarini, J.; Opdenakker, G.; Snoeck, R. In vitro-selected drug-resistant varicella-zoster virus mutants in the thymidine kinase and DNA polymerase genes yield novel phenotype-genotype asso-ciations and highlight differences between antiherpesvirus drugs. J. Virol. 2012, 86, 2641–2652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prichard, M.; Prichard, L.E.; Shipman, C. Inhibitors of thymidylate synthase and dihydrofolate reductase potentiate the antiviral effect of acyclovir. Antivir. Res. 1993, 20, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Oliver, S.L.; Zerboni, L.; Sommer, M.; Rajamani, J.; Arvin, A.M. Development of recombinant varicella-zoster viruses expressing luciferase fusion proteins for live in vivo imaging in human skin and dorsal root ganglia xenografts. J. Virol. Methods 2008, 154, 182–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debrus, S.; Sadzot-Delvaux, C.; Nikkels, A.F.; Piette, J.; Rentier, B. Varicella-zoster virus gene 63 encodes an immedi-ate-early protein that is abundantly expressed during latency. J. Virol. 1995, 69, 3240–3245. [Google Scholar] [CrossRef] [Green Version]
- Jones, M.; Dry, I.R.; Frampton, D.; Singh, M.; Kanda, R.; Yee, M.; Kellam, P.; Hollinshead, M.; Kinchington, P.R.; O’Toole, E.; et al. RNA-seq Analysis of Host and Viral Gene Expression Highlights Interaction between Varicella Zoster Virus and Keratinocyte Differentiation. PLoS Pathog. 2014, 10, e1003896. [Google Scholar] [CrossRef] [Green Version]
- Balliet, J.W.; Min, J.C.; Cabatingan, M.S.; Schaffer, P.A. Site-directed mutagenesis of large DNA palindromes: Construction and in vitro characterization of herpes simplex virus type 1 mutants containing point mutations that eliminate the oriL or oriS initiation function. J. Virol. 2005, 79, 12783–12797. [Google Scholar] [CrossRef] [Green Version]
- Weller, S.K.; Spadaro, A.; Schaffer, J.E.; Murray, A.W.; Maxam, A.M.; Schaffer, P.A. Cloning, sequencing, and func-tional analysis of oriL, a herpes simplex virus type 1 origin of DNA synthesis. Mol. Cell Biol. 1985, 5, 930–942. [Google Scholar]
- Polvino-Bodnar, M.; Orberg, P.K.; Schaffer, P.A. Herpes simplex virus type 1 oriL is not required for virus replication or for the establishment and reactivation of latent infection in mice. J. Virol. 1987, 61, 3528–3535. [Google Scholar] [CrossRef] [Green Version]
- Rostad, C.A.; Currier, M.C.; Moore, M.L. Fluorescent and Bioluminescent Reporter Myxoviruses. Viruses 2016, 8, 214. [Google Scholar] [CrossRef]
- Fulton, B.O.; Palese, P.; Heaton, N.S. Replication-Competent Influenza B Reporter Viruses as Tools for Screening Anti-virals and Antibodies. J. Virol. 2015, 89, 12226–12231. [Google Scholar] [CrossRef] [Green Version]
- Cilloniz, C.; Jackson, W.; Grose, C.; Czechowski, D.; Hay, J.; Ruyechan, W.T. The Varicella-Zoster Virus (VZV) ORF9 Protein Interacts with the IE62 Major VZV Transactivator. J. Virol. 2007, 81, 761–774. [Google Scholar] [CrossRef] [Green Version]
- Braspenning, S.E.; Sadaoka, T.; Breuer, J.; Verjans, G.M.G.M.; Ouwendijk, W.J.D.; Depledge, D.P. Decoding the Architecture of the Varicella-Zoster Virus Transcriptome. mBio 2020, 11, e01568-20. [Google Scholar] [CrossRef]
- Cai, M.; Wang, S.; Xing, J.; Zheng, C. Characterization of the nuclear import and export signals, and subcellular transport mechanism of varicella-zoster virus ORF. J. Gen. Virol. 2010, 92, 621–626. [Google Scholar] [CrossRef]
- Lebrun, M.; Thelen, N.; Thiry, M.; Riva, L.; Ote, I.; Condé, C.; Vandevenne, P.; Di Valentin, E.; Bontems, S.; Sadzot-Delvaux, C. Varicella-zoster virus induces the formation of dynamic nuclear capsid aggregates. Virology 2014, 454–455, 311–327. [Google Scholar] [CrossRef]
- Storlie, J.; Jackson, W.; Hutchinson, J.; Grose, C. Delayed biosynthesis of varicella-zoster virus glycoprotein C: Upregulation by hexamethylene bisacetamide and retinoic acid treatment of infected cells. J. Virol. 2006, 80, 9544–9556. [Google Scholar] [CrossRef] [Green Version]
- Sato, H.; Callanan, L.D.; Pesnicak, L.; Krogmann, T.; Cohen, J.I. Varicella-zoster virus (VZV) ORF17 protein induces RNA cleavage and is critical for replication of VZV at 37 degrees C but not 33 degrees C. J. Virol. 2002, 76, 11012–11023. [Google Scholar] [CrossRef] [Green Version]
- Desloges, N.; Rahaus, M.; Wolff, M.H. The varicella–zoster virus–mediated delayed host shutoff: Open reading frame 17 has no major function, whereas immediate–early 63 protein represses heterologous gene expression. Microbes Infect. 2005, 7, 1519–1529. [Google Scholar] [CrossRef]
- Roviš, T.L.; Bailer, S.M.; Pothineni, V.R.; Ouwendijk, W.J.; Šimić, H.; Babić, M.; Miklić, K.; Malić, S.; Verweij, M.C.; Baiker, A.; et al. Comprehensive Analysis of Varicella-Zoster Virus Proteins Using a New Monoclonal Antibody Collection. J. Virol. 2013, 87, 6943–6954. [Google Scholar] [CrossRef] [Green Version]
- Strasfeld, L.; Chou, S. Antiviral drug resistance: Mechanisms and clinical implications. Infect. Dis. Clin. 2010, 24, 809–833. [Google Scholar] [CrossRef]
- Morfin, F.; Thouvenot, D.; De Turenne-Tessier, M.; Lina, B.; Aymard, M.; Ooka, T. Phenotypic and genetic characteriza-tion of thymidine kinase from clinical strains of varicella-zoster virus resistant to acyclovir. Antimicrob. Agents Chemother. 1999, 43, 2412–2416. [Google Scholar] [CrossRef] [Green Version]
- Heineman, T.C.; Cohen, J.I. Deletion of the varicella-zoster virus large subunit of ribonucleotide reductase impairs growth of virus in vitro. J. Virol. 1994, 68, 3317–3323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, J.; Williams, M.; Cohen, J.I. Disruption of the varicella-zoster virus dUTPase and the adjacent ORF9A gene results in impaired growth and reduced syncytia formation in vitro. Virology 1997, 234, 186–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Target | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| fLuc Firefly (Photinus pyralis) Luciferase | GAGGGATCCGGTTCCGGAGAGGGCAGAGGAAGTCTGCTAACATGCGGTGACGTCGAGGAGAATCCTGGCCCAATGGAAGACGCC AAAAACATA | AATTCGAATTCGCGCGCAGATCTTTACACGGCGATCTTTCCGCCCTTCTTGGC |
| Zeo (Zeomycin resistance) | AGATCTAGATCTCGAGTAATGGAACGGACCGTGTTGAC | GCTGACGTCGACGAATTCTGATCACTCAAGTTTCGAGGTCGAGGTG |
| gCLuc (F2 and R3; ORF14) | CTTATCGCAGTTATCGCAACCCTATGCATCCGTTGCTGTTCAATGGACGAGCTGTACAAG | AATAAAATGATATACACAGACGCGTTTGGTTGGTTTCTGTCAGTCCTGCTCCTCAGCC |
| ORF9Luc (F2 and R3) | AGTAGGGCCCGTTCGGCATCAAGAACTGATGCGCGAAAATCAATGGACGAGCTGTACAAG | ACGTTTATTTATTATACATAATACCGGGTAAACCGTTACTTCAGTCCTGCTCCTCAGCC |
| ORF17Luc (F2 and R2) | CTCCACTCCCACTAAACACTGTATTAACAAAATATTGGAATTCAATGGACGAGCTGTACAAG | AGCAAAATAAAACAATGAACCATTAAGTCGCTCTTATGTGTGTCAGTCCTGCTCCTCAGCC |
| 57Luc (ORF57) | ACGTTGAGGAGCCTTGCAGGTTGGGTGCCGCGCTTCACCGTCAATGGACGAGCTGTACAAG | TTTATATTTAACGGCTTTTAATTTGAAGACACCTATCCTCTCAGTCCTGCTCCTC |
| 40REP (ORF40) | ATATAGATATTACGTTTATCATGCCAATGGGAGTGTTTCAGGCGAATTCCATGGACAGATATACACGACAGGATGACGACGATAAGTAGG | CAGTTGAAAAATCGCCGGCGTGTCGTGTATATCTGTCCATGGAATTCGCCTGAAACACTCCCATTGGCACAGGGTAATGCCAGTGTTAC |
| 50REP (ORF50) | TGAATTATCCAAATTCGCCAATTAAGCGTATCCATTTGATGATCTAAAGCTTCCACCTCGGGTGTCGTGGTGTCGTACGGGGATGACGACGATAAGTAGG | TCTGAAAAAGTCTCACCGTACGACACCACGACACCCGAGGTGGAAGCTTTAGATCATCAAATGGATACGCTTAATAGGGTAATGCCAGTGTTAC |
| TS (TS T2B-For and endR2) | TATCAAGTGGTCGTTTGTATTTAACGATTATTACCGGTACCGGTACCATGGTGAGCAAGGGCGAGGAG | ACATCTACTGTCTTGACAACATTTAAAAATCCATTAAAGATTATCTGGATCCTACCTTTC |
| TK (TK T2BFor and endR3) | CGCCGAAGAATTTTTACACCACTTTGCAATAACACCAAACGGTACCATGGTGAGCAAGGGCGAGGAG | TGTGTATCATCTTTTTACTGGTACATACGTAAATACTAGGTTATCTGGATCCTACCTTTC |
| TKkan-del (For and Rev) | CGCCGAAGAATTTTTACACCACTTTGCAATAACACCAAACGGATGACGACGATAAGTAGG | TGTGTATCATCTTTTTACTGGTACATACGTAAATACTAGGAGGGTAATGCCAGTGTTAC |
| Location of Mutation | Identification of Mutation and the Correction |
|---|---|
| ORF40 mutation in pOka DX | 72661 atg cca atg gga gtg ttt cag Aca aac tcc atg gac aga tat aca cga cac … M P M G V F Q T428 N S M E R Y T R H |
| ORF40 Correction in pOka DX-RR | 72661 atg cca atg gga gtg ttt cag GcG aaT tcc atg gac aga tat aca cga cac … M P M G V F Q A428 N S M E R Y T R H |
| ORF50 mutation in pOka DX | gaa aaa gtc tca ccg tac gac acc acg aca ccc gag gCg gaa gcg tta gat cat caa atg E11 K V S P Y D T T T P E A E A L D H Q M |
| ORF50 correction in pOka DX-RR | gaa aaa gtc tca ccg tac gac acc acg aca ccc gag gTg gaa gcT tta gat cat caa atg E11 K V S P Y D T T T P E V E A L D H Q M |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lloyd, M.G.; Yee, M.B.; Flot, J.S.; Liu, D.; Geiler, B.W.; Kinchington, P.R.; Moffat, J.F. Development of Robust Varicella Zoster Virus Luciferase Reporter Viruses for In Vivo Monitoring of Virus Growth and Its Antiviral Inhibition in Culture, Skin, and Humanized Mice. Viruses 2022, 14, 826. https://doi.org/10.3390/v14040826
Lloyd MG, Yee MB, Flot JS, Liu D, Geiler BW, Kinchington PR, Moffat JF. Development of Robust Varicella Zoster Virus Luciferase Reporter Viruses for In Vivo Monitoring of Virus Growth and Its Antiviral Inhibition in Culture, Skin, and Humanized Mice. Viruses. 2022; 14(4):826. https://doi.org/10.3390/v14040826
Chicago/Turabian StyleLloyd, Megan G., Michael B. Yee, Joseph S. Flot, Dongmei Liu, Brittany W. Geiler, Paul R. Kinchington, and Jennifer F. Moffat. 2022. "Development of Robust Varicella Zoster Virus Luciferase Reporter Viruses for In Vivo Monitoring of Virus Growth and Its Antiviral Inhibition in Culture, Skin, and Humanized Mice" Viruses 14, no. 4: 826. https://doi.org/10.3390/v14040826