
Intermediate Monocytes with PD-L1 and CD62L Expression as a Possible Player in Active SARS-CoV-2 Infection

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Materials
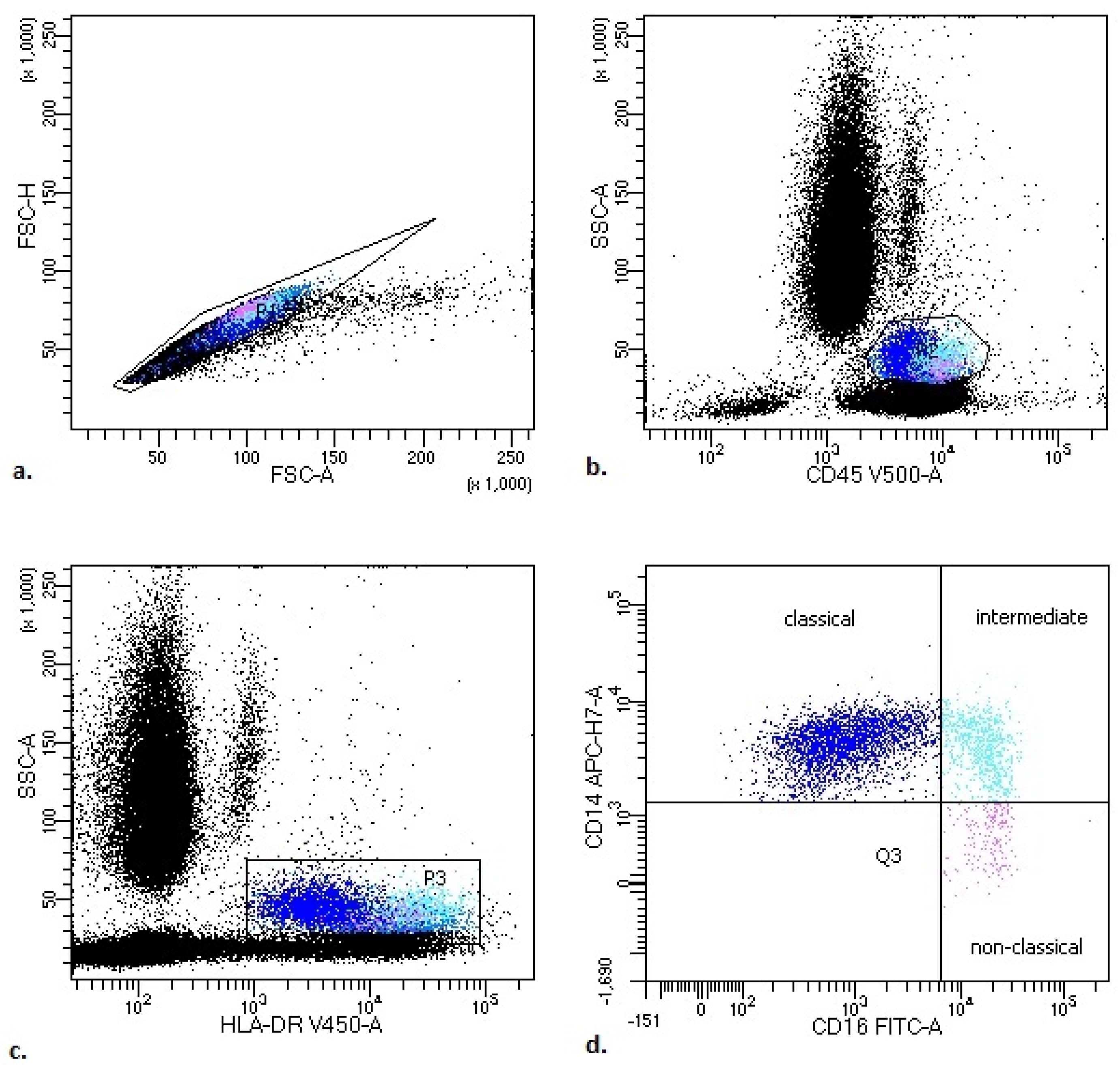
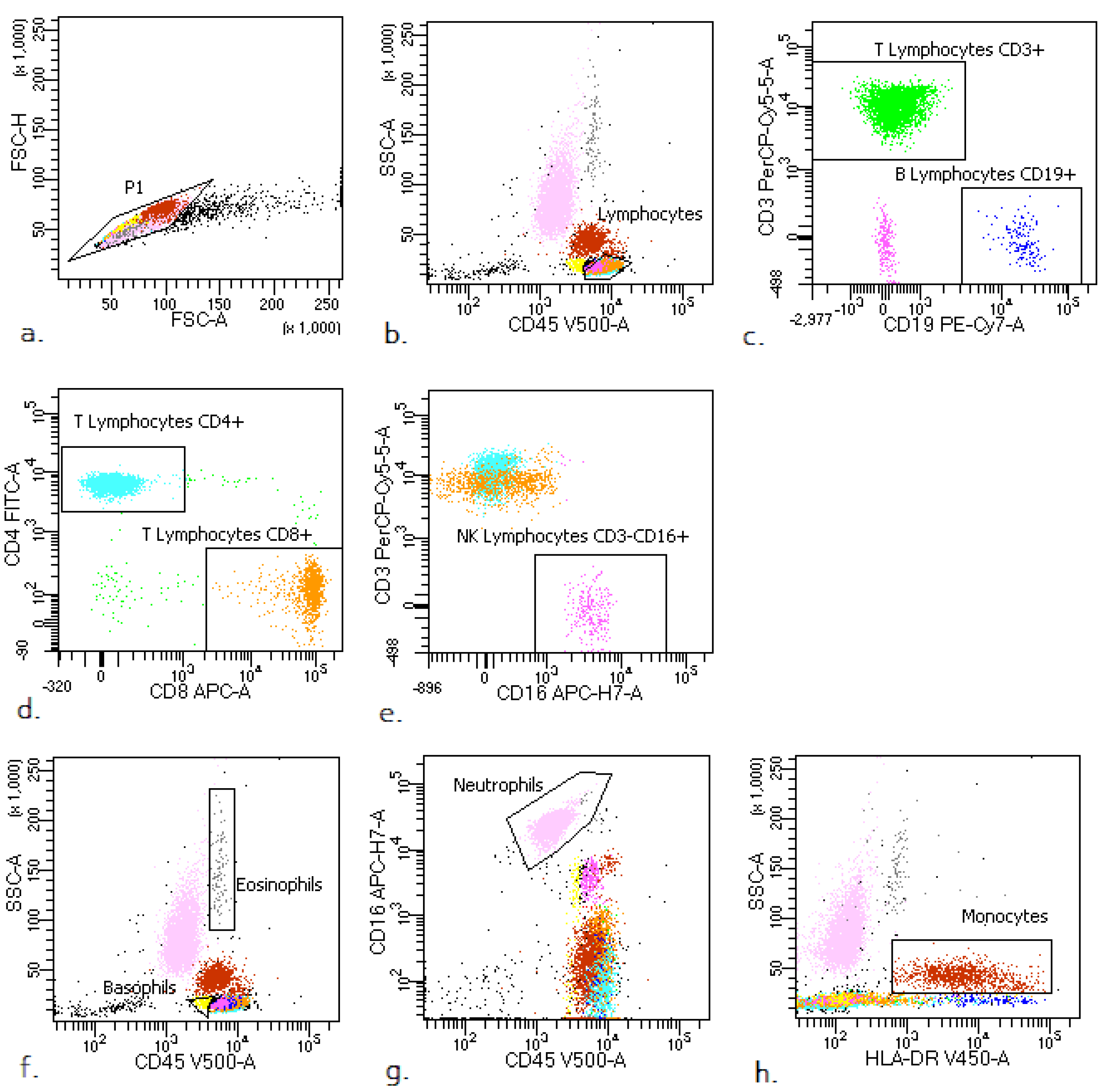
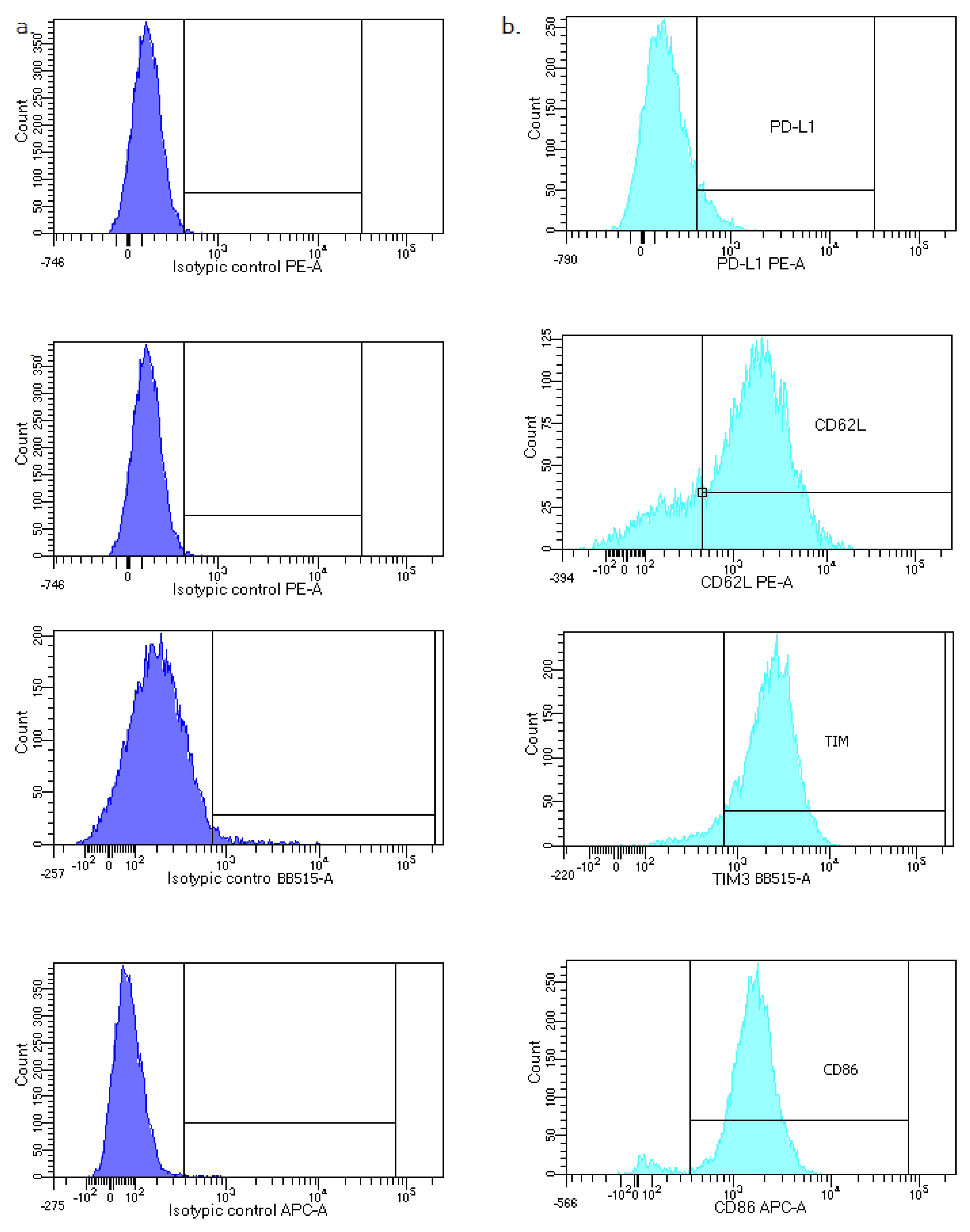
2.3. Flow Cytometry Analysis
- lymphocytes: CD45+ bright SSClow;
- lymphocytes T: CD45+ bright SSClow CD3+;
- lymphocytes B: CD45+ bright SSClow CD19+;
- NK cells: CD45+ bright SSC low CD3− CD16+;
- neutrophils: CD45+ SSCbright CD16+;
- eosinophils: CD45+ bright SSCbright;
- basophils: CD45+ dim SSClow;
- monocytes: CD45+ bright SSC+ HLA-DR+.
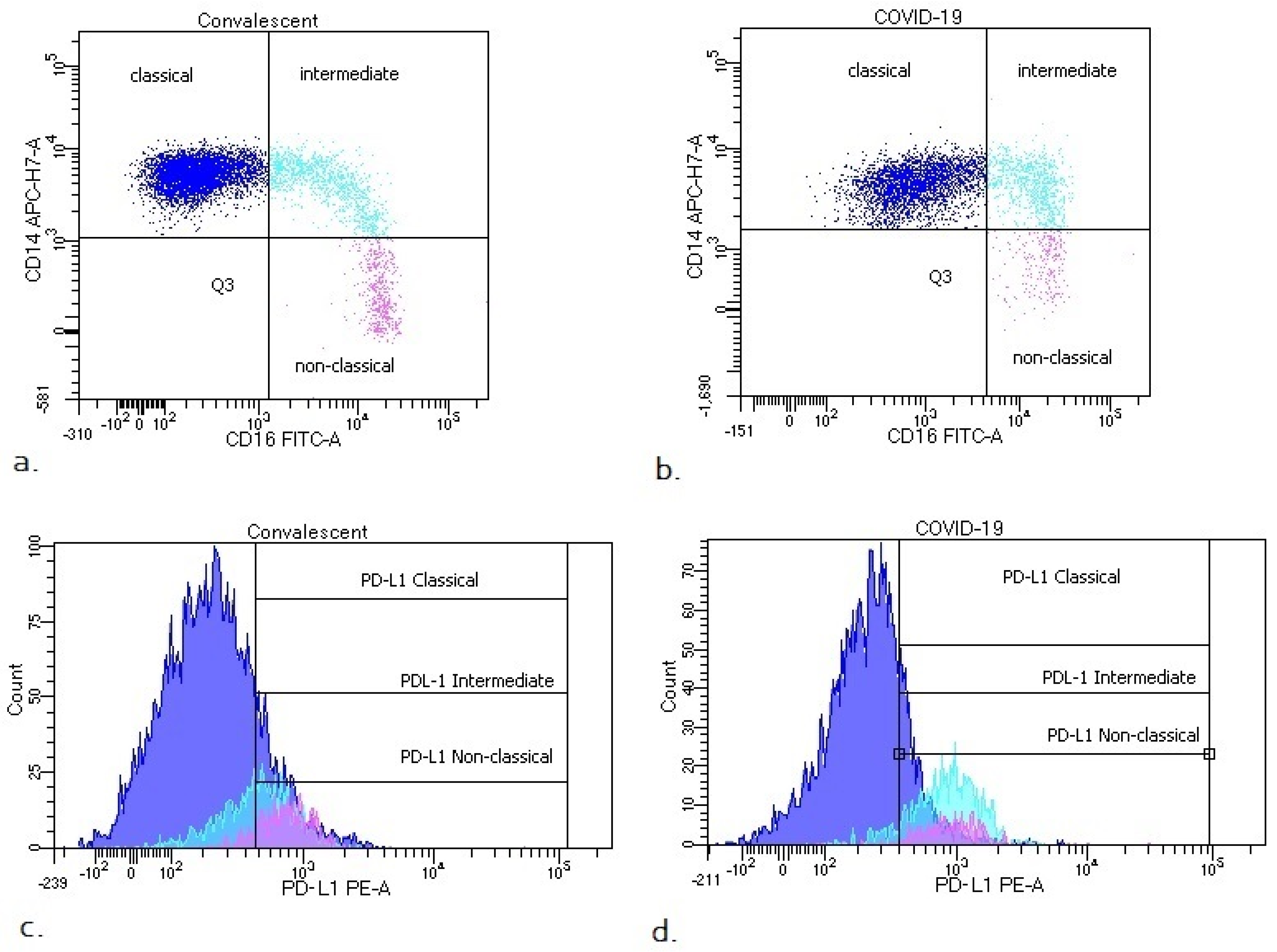
- Classical monocytes: CD14++ CD16−;
- Intermediate monocytes: CD14+ CD16+;
- Non classical monocytes: CD14−/+ CD16++.
2.4. Statistical Analysis
3. Results
3.1. Patients’ Characteristics, White Blood Cell (WBC) Count, Leukocytes and Main Lymphocyte Subpopulation Counts in Study Groups
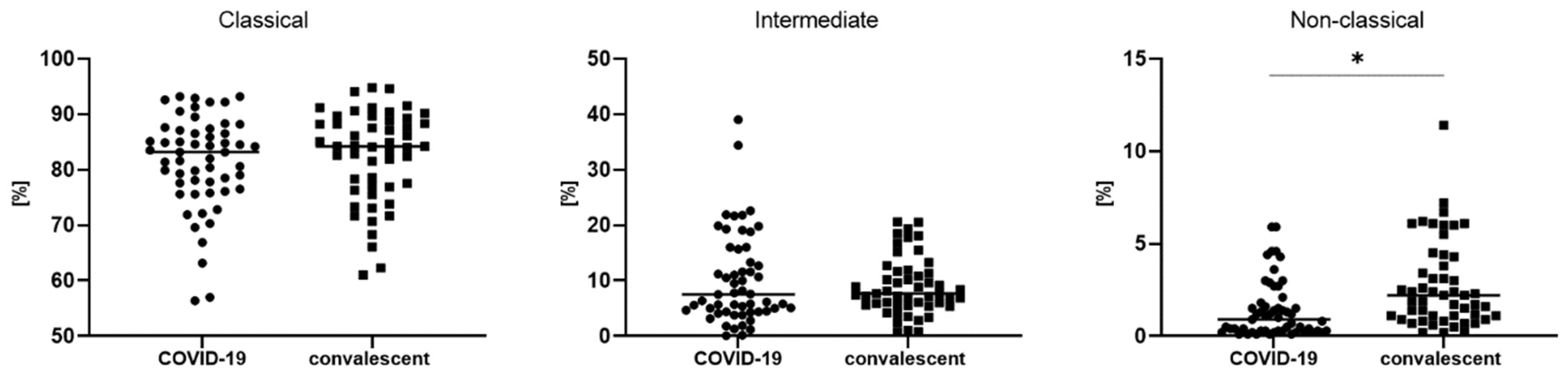
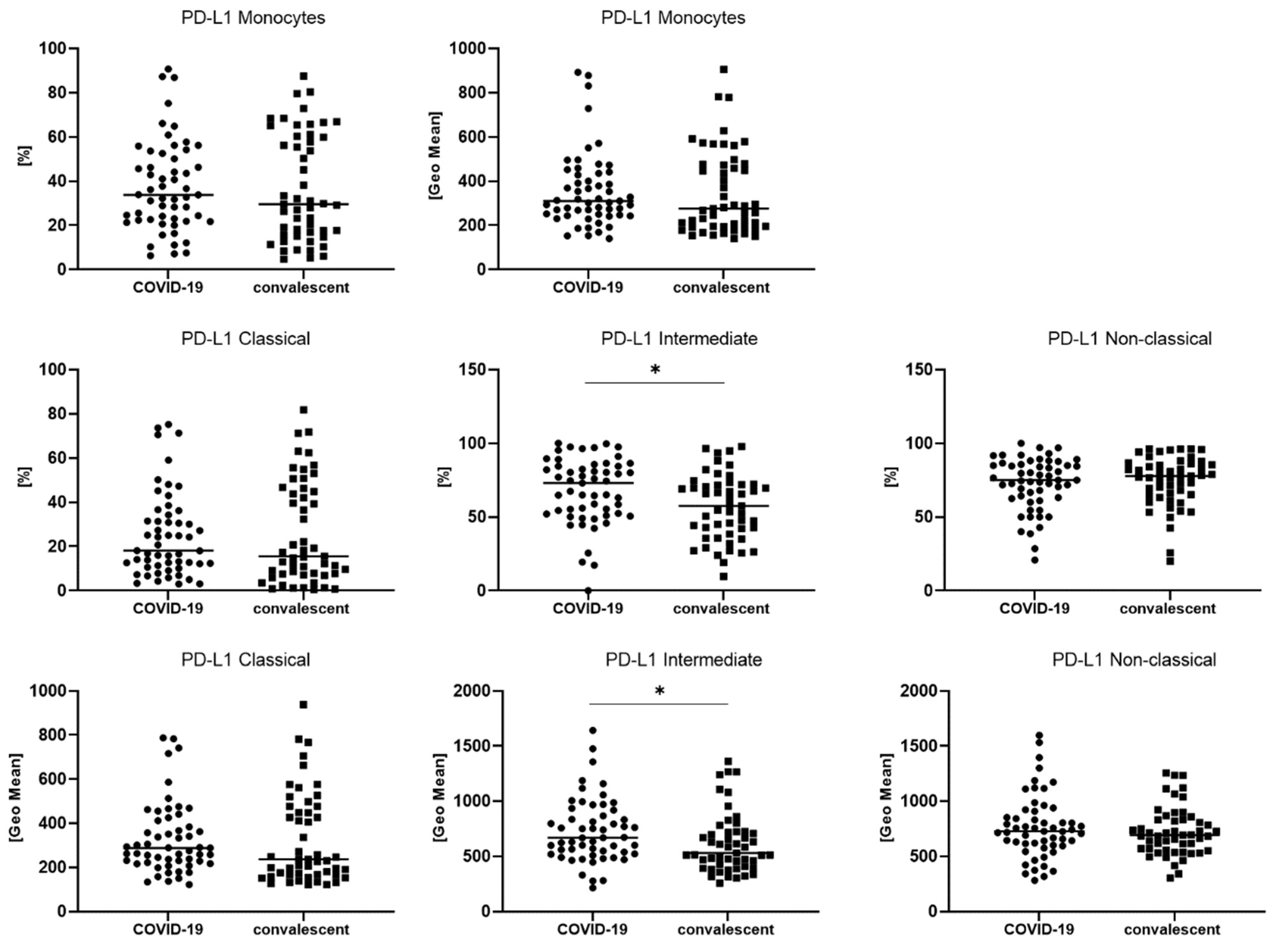
3.2. Differences in Monocyte Subsets: Classical/Intermediate/Non-Classical and Monocyte Subpopulations with PD-L1 Expression
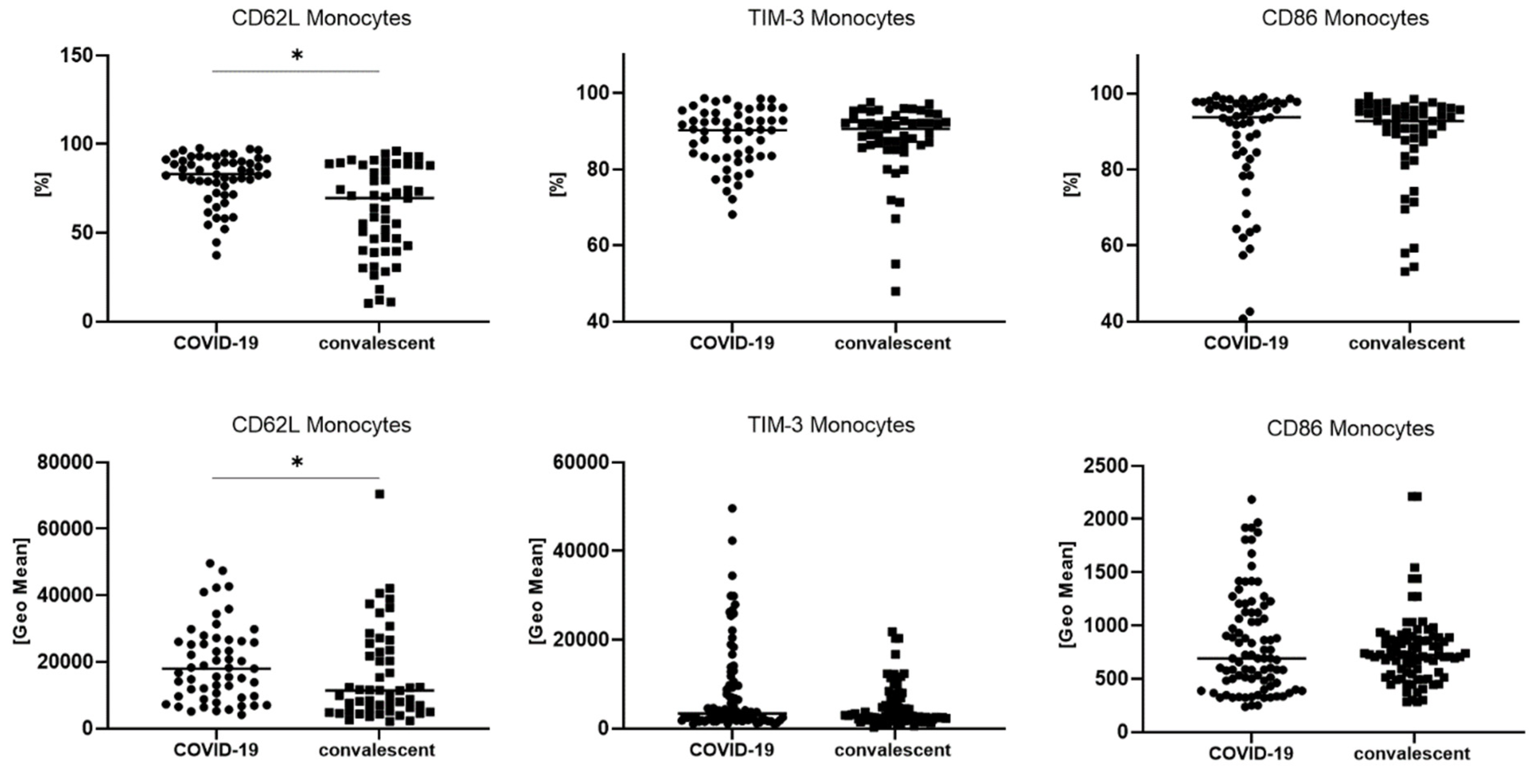
3.3. The Difference in the Number of TIM-3, CD62L and CD86 Postive Monocytes between COVID-19 and Convalescent Patients
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patients with COVID-19 a Median (Q1–Q3) | Convalescent b Median (Q1–Q3) | Control Group c Median (Q1–Q3) | p < 0.05 * Group A-B-C ANOVA, Kruskal–Wallis | p < 0.05 * between Groups Post-Hoc | |
|---|---|---|---|---|---|
| All monocytes [%] | 5.5 (3.8–7.6) | 7.0 (4.6–9.1) | 6.4 (4.4–7.5) | p = 0.1038 | - |
| All monocytes [k/ul] | 376 (260–453) | 535 (399–815) | 356 (287–437) | * p < 0.0001 | * a–b, b–c |
| Classical monocytes CD14++ CD16− | 83.2 (76.5–87.1) | 84.2 (76.9–88.8) | 67.3 (60.7–70.9) | * p < 0.0001 | * a–c, b–c |
| Intermediate monocytes CD14+ CD16+ | 7.5 (4.4–15.7) | 7.7 (5.6–11.7) | 5.2 (3.9–7.2) | * p = 0.0472 | - |
| Non-classical monocytes CD14−/+ CD16++ | 0.9 (0.3–1.8) | 2.2 (1.1–4.3) | 14.9 (12.5–16.8) | * p < 0.0001 | * a–b, a–c, b–c |


References
- Hu, B.; Guo, H.; Zhou, P.; Shi, Z.L. Characteristics of SARS-CoV-2 and COVID-19. Nat. Rev. Microbiol. 2021, 19, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Gibson, P.G.; Qin, L.; Puah, S.H. COVID-19 acute respiratory distress syndrome (ARDS): Clinical features and differences from typical pre-COVID-19 ARDS. Med. J. Aust. 2020, 213, 54–56.e1. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.Y.; Danielson, M.L.; Town, M.; Derado, G.; Greenlund, K.J.; Kirley, P.D.; Alden, N.B.; Yousey-Hindes, K.; Anderson, E.J.; Ryan, P.A.; et al. Risk Factors for Coronavirus Disease 2019 (COVID-19)-Associated Hospitalization: COVID-19-Associated Hospitalization Surveillance Network and Behavioral Risk Factor Surveillance System. Clin. Infect. Dis. 2021, 72, e695–e703. [Google Scholar] [CrossRef] [PubMed]
- Petrakis, D.; Margina, D.; Tsarouhas, K.; Tekos, F.; Stan, M.; Nikitovic, D.; Kouretas, D.; Spandidos, D.A.; Tsatsakis, A. Obesity a risk factor for increased COVID19 prevalence, severity and lethality (Review). Mol. Med. Rep. 2020, 22, 9–19. [Google Scholar] [CrossRef]
- Song, P.; Li, W.; Xie, J.; Hou, Y.; You, C. Cytokine storm induced by SARS-CoV-2. Clin. Chim. Acta 2020, 509, 280–287. [Google Scholar] [CrossRef]
- Hosseini, A.; Hashemi, V.; Shomali, N.; Asghari, F.; Gharibi, T.; Akbari, M.; Gholizadeh, S.; Jafari, A. Innate and adaptive immune responses against coronavirus. Biomed. Pharmacother. 2020, 132, 110859. [Google Scholar] [CrossRef]
- Lee, J.; Park, S.S.; Kim, T.Y.; Lee, D.G.; Kim, D.W. Lymphopenia as a Biological Predictor of Outcomes in COVID-19 Patients: A Nationwide Cohort Study. Cancers 2021, 13, 471. [Google Scholar] [CrossRef]
- Cortes-Vieyra, R.; Gutierrez-Castellanos, S.; Alvarez-Aguilar, C.; Baizabal-Aguirre, V.M.; Nunez-Anita, R.E.; Rocha-Lopez, A.G.; Gomez-Garcia, A. Behavior of Eosinophil Counts in Recovered and Deceased COVID-19 Patients over the Course of the Disease. Viruses 2021, 13, 1675. [Google Scholar] [CrossRef]
- Borges, R.C.; Hohmann, M.S.; Borghi, S.M. Dendritic cells in COVID-19 immunopathogenesis: Insights for a possible role in determining disease outcome. Int. Rev. Immunol. 2021, 40, 108–125. [Google Scholar] [CrossRef]
- Rutkowska, E.; Kwiecien, I.; Kulik, K.; Chelstowska, B.; Klos, K.; Rzepecki, P.; Chcialowski, A. Usefulness of the New Hematological Parameter: Reactive Lymphocytes RE-LYMP with Flow Cytometry Markers of Inflammation in COVID-19. Cells 2021, 10, 82. [Google Scholar] [CrossRef]
- Kwiecien, I.; Rutkowska, E.; Klos, K.; Wiesik-Szewczyk, E.; Jahnz-Rozyk, K.; Rzepecki, P.; Chcialowski, A. Maturation of T and B Lymphocytes in the Assessment of the Immune Status in COVID-19 Patients. Cells 2020, 9, 2615. [Google Scholar] [CrossRef] [PubMed]
- Kwiecien, I.; Rutkowska, E.; Kulik, K.; Klos, K.; Plewka, K.; Raniszewska, A.; Rzepecki, P.; Chcialowski, A. Neutrophil Maturation, Reactivity and Granularity Research Parameters to Characterize and Differentiate Convalescent Patients from Active SARS-CoV-2 Infection. Cells 2021, 10, 2332. [Google Scholar] [CrossRef] [PubMed]
- Reusch, N.; De Domenico, E.; Bonaguro, L.; Schulte-Schrepping, J.; Bassler, K.; Schultze, J.L.; Aschenbrenner, A.C. Neutrophils in COVID-19. Front. Immunol. 2021, 12, 652470. [Google Scholar] [CrossRef]
- Rutkowska, E.; Kwiecien, I.; Zabicka, M.; Maliborski, A.; Raniszewska, A.; Klos, K.; Urbanska, W.; Klajnowicz, I.; Rzepecki, P.; Chcialowski, A. Cytokines and Leukocytes Subpopulations Profile in SARS-CoV-2 Patients Depending on the CT Score Severity. Viruses 2021, 13, 880. [Google Scholar] [CrossRef]
- Parihar, A.; Eubank, T.D.; Doseff, A.I. Monocytes and macrophages regulate immunity through dynamic networks of survival and cell death. J. Innate Immun. 2010, 2, 204–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaplin, D.D. Overview of the immune response. J. Allergy Clin. Immunol. 2010, 125, S3–S23. [Google Scholar] [CrossRef]
- Marimuthu, R.; Francis, H.; Dervish, S.; Li, S.C.H.; Medbury, H.; Williams, H. Characterization of Human Monocyte Subsets by Whole Blood Flow Cytometry Analysis. J. Vis. Exp. 2018, 140, e57941. [Google Scholar] [CrossRef] [Green Version]
- Kwiecien, I.; Rutkowska, E.; Polubiec-Kownacka, M.; Raniszewska, A.; Rzepecki, P.; Domagala-Kulawik, J. Blood Monocyte Subsets with Activation Markers in Relation with Macrophages in Non-Small Cell Lung Cancer. Cancers 2020, 12, 2513. [Google Scholar] [CrossRef]
- Rossol, M.; Kraus, S.; Pierer, M.; Baerwald, C.; Wagner, U. The CD14(bright) CD16+ monocyte subset is expanded in rheumatoid arthritis and promotes expansion of the Th17 cell population. Arthritis Rheum. 2012, 64, 671–677. [Google Scholar] [CrossRef]
- Wildgruber, M.; Aschenbrenner, T.; Wendorff, H.; Czubba, M.; Glinzer, A.; Haller, B.; Schiemann, M.; Zimmermann, A.; Berger, H.; Eckstein, H.H.; et al. The “Intermediate” CD14(++)CD16(+) monocyte subset increases in severe peripheral artery disease in humans. Sci. Rep. 2016, 6, 39483. [Google Scholar] [CrossRef] [Green Version]
- Stansfield, B.K.; Ingram, D.A. Clinical significance of monocyte heterogeneity. Clin. Transl. Med. 2015, 4, 5. [Google Scholar] [CrossRef]
- Adams, D.H.; Shaw, S. Leucocyte-endothelial interactions and regulation of leucocyte migration. Lancet 1994, 343, 831–836. [Google Scholar] [CrossRef]
- Belge, K.U.; Dayyani, F.; Horelt, A.; Siedlar, M.; Frankenberger, M.; Frankenberger, B.; Espevik, T.; Ziegler-Heitbrock, L. The proinflammatory CD14+CD16+DR++ monocytes are a major source of TNF. J. Immunol. 2002, 168, 3536–3542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, M.; Zhu, C.; Kuchroo, V.K. Tim-3 and its role in regulating anti-tumor immunity. Immunol. Rev. 2017, 276, 97–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, J.; Zhang, L.; Wu, H.; Gao, P.; Xu, Z. Increased Tim-3(+) monocytes/macrophages are associated with disease severity in patients with IgA nephropathy. Int. Immunopharmacol. 2021, 97, 107666. [Google Scholar] [CrossRef] [PubMed]
- Sakuishi, K.; Jayaraman, P.; Behar, S.M.; Anderson, A.C.; Kuchroo, V.K. Emerging Tim-3 functions in antimicrobial and tumor immunity. Trends Immunol. 2011, 32, 345–349. [Google Scholar] [CrossRef] [Green Version]
- Yasuoka, H.; Asai, A.; Ohama, H.; Tsuchimoto, Y.; Fukunishi, S.; Higuchi, K. Increased both PD-L1 and PD-L2 expressions on monocytes of patients with hepatocellular carcinoma was associated with a poor prognosis. Sci. Rep. 2020, 10, 10377. [Google Scholar] [CrossRef]
- Butte, M.J.; Pena-Cruz, V.; Kim, M.J.; Freeman, G.J.; Sharpe, A.H. Interaction of human PD-L1 and B7-1. Mol. Immunol. 2008, 45, 3567–3572. [Google Scholar] [CrossRef] [Green Version]
- Flisiak, R.; Horban, A.; Jaroszewicz, J.; Kozielewicz, D.; Mastalerz-Migas, A.; Owczuk, R.; Parczewski, M.; Pawlowska, M.; Piekarska, A.; Simon, K.; et al. Management of SARS-CoV-2 infection: Recommendations of the Polish Association of Epidemiologists and Infectiologists as of 26 April 2021. Pol. Arch. Intern. Med. 2021, 131, 487–496. [Google Scholar] [CrossRef]
- Liu, J.; Li, H.; Luo, M.; Liu, J.; Wu, L.; Lin, X.; Li, R.; Wang, Z.; Zhong, H.; Zheng, W.; et al. Lymphopenia predicted illness severity and recovery in patients with COVID-19: A single-center, retrospective study. PLoS ONE 2020, 15, e0241659. [Google Scholar] [CrossRef]
- Mateos Gonzalez, M.; Sierra Gonzalo, E.; Casado Lopez, I.; Arnalich Fernandez, F.; Beato Perez, J.L.; Monge Monge, D.; Vargas Nunez, J.A.; Garcia Fenoll, R.; Suarez Fernandez, C.; Freire Castro, S.J.; et al. The Prognostic Value of Eosinophil Recovery in COVID-19: A Multicentre, Retrospective Cohort Study on Patients Hospitalised in Spanish Hospitals. J. Clin. Med. 2021, 10, 305. [Google Scholar] [CrossRef] [PubMed]
- Schulte-Schrepping, J.; Reusch, N.; Paclik, D.; Bassler, K.; Schlickeiser, S.; Zhang, B.; Kramer, B.; Krammer, T.; Brumhard, S.; Bonaguro, L.; et al. Severe COVID-19 Is Marked by a Dysregulated Myeloid Cell Compartment. Cell 2020, 182, 1419–1440.e23. [Google Scholar] [CrossRef] [PubMed]
- Liao, M.; Liu, Y.; Yuan, J.; Wen, Y.; Xu, G.; Zhao, J.; Cheng, L.; Li, J.; Wang, X.; Wang, F.; et al. Single-cell landscape of bronchoalveolar immune cells in patients with COVID-19. Nat. Med. 2020, 26, 842–844. [Google Scholar] [CrossRef]
- Qin, S.; Jiang, Y.; Wei, X.; Liu, X.; Guan, J.; Chen, Y.; Lu, H.; Qian, J.; Wang, Z.; Lin, X. Dynamic changes in monocytes subsets in COVID-19 patients. Hum. Immunol. 2021, 82, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Spinetti, T.; Hirzel, C.; Fux, M.; Walti, L.N.; Schober, P.; Stueber, F.; Luedi, M.M.; Schefold, J.C. Reduced Monocytic Human Leukocyte Antigen-DR Expression Indicates Immunosuppression in Critically Ill COVID-19 Patients. Anesth. Analg. 2020, 131, 993–999. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Shi, L.; Wang, Y.; Zhang, J.; Huang, L.; Zhang, C.; Liu, S.; Zhao, P.; Liu, H.; Zhu, L.; et al. Pathological findings of COVID-19 associated with acute respiratory distress syndrome. Lancet Respir. Med. 2020, 8, 420–422. [Google Scholar] [CrossRef]
- Narasimhan, P.B.; Marcovecchio, P.; Hamers, A.A.J.; Hedrick, C.C. Nonclassical Monocytes in Health and Disease. Annu. Rev. Immunol. 2019, 37, 439–456. [Google Scholar] [CrossRef]
- Tacke, F.; Alvarez, D.; Kaplan, T.J.; Jakubzick, C.; Spanbroek, R.; Llodra, J.; Garin, A.; Liu, J.; Mack, M.; van Rooijen, N.; et al. Monocyte subsets differentially employ CCR2, CCR5, and CX3CR1 to accumulate within atherosclerotic plaques. J. Clin. Investig. 2007, 117, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Naranjo-Gomez, J.S.; Castillo, J.A.; Rojas, M.; Restrepo, B.N.; Diaz, F.J.; Velilla, P.A.; Castano, D. Different phenotypes of non-classical monocytes associated with systemic inflammation, endothelial alteration and hepatic compromise in patients with dengue. Immunology 2019, 156, 147–163. [Google Scholar] [CrossRef]
- Ferreira, A.C.; Soares, V.C.; de Azevedo-Quintanilha, I.G.; Dias, S.; Fintelman-Rodrigues, N.; Sacramento, C.Q.; Mattos, M.; de Freitas, C.S.; Temerozo, J.R.; Teixeira, L.; et al. SARS-CoV-2 engages inflammasome and pyroptosis in human primary monocytes. Cell Death Discov. 2021, 7, 43. [Google Scholar] [CrossRef]
- Cros, J.; Cagnard, N.; Woollard, K.; Patey, N.; Zhang, S.Y.; Senechal, B.; Puel, A.; Biswas, S.K.; Moshous, D.; Picard, C.; et al. Human CD14dim monocytes patrol and sense nucleic acids and viruses via TLR7 and TLR8 receptors. Immunity 2010, 33, 375–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilk, A.J.; Rustagi, A.; Zhao, N.Q.; Roque, J.; Martinez-Colon, G.J.; McKechnie, J.L.; Ivison, G.T.; Ranganath, T.; Vergara, R.; Hollis, T.; et al. A single-cell atlas of the peripheral immune response in patients with severe COVID-19. Nat. Med. 2020, 26, 1070–1076. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Cerrillo, I.; Landete, P.; Aldave, B.; Sanchez-Alonso, S.; Sanchez-Azofra, A.; Marcos-Jimenez, A.; Avalos, E.; Alcaraz-Serna, A.; de Los Santos, I.; Mateu-Albero, T.; et al. COVID-19 severity associates with pulmonary redistribution of CD1c+ DCs and inflammatory transitional and nonclassical monocytes. J. Clin. Investig. 2020, 130, 6290–6300. [Google Scholar] [CrossRef] [PubMed]
- Pan, T.; Zhou, T.; Li, L.; Liu, Z.; Chen, Y.; Mao, E.; Li, M.; Qu, H.; Liu, J. Monocyte programmed death ligand-1 expression is an early marker for predicting infectious complications in acute pancreatitis. Crit. Care 2017, 21, 186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.H.; Wu, M.F.; Hsu, C.Y.; Lin, S.Y.; Chang, Y.N.; Lee, H.S.; Wei, Y.F.; Shu, C.C. The Dynamic Change of Immune Checkpoints and CD14+ Monocytes in Latent Tuberculosis Infection. Biomedicines 2021, 9, 1479. [Google Scholar] [CrossRef] [PubMed]
- Keir, M.E.; Butte, M.J.; Freeman, G.J.; Sharpe, A.H. PD-1 and its ligands in tolerance and immunity. Annu. Rev. Immunol. 2008, 26, 677–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabbatino, F.; Conti, V.; Franci, G.; Sellitto, C.; Manzo, V.; Pagliano, P.; De Bellis, E.; Masullo, A.; Salzano, F.A.; Caputo, A.; et al. PD-L1 Dysregulation in COVID-19 Patients. Front. Immunol. 2021, 12, 695242. [Google Scholar] [CrossRef]
- Aghbash, P.S.; Eslami, N.; Shamekh, A.; Entezari-Maleki, T.; Baghi, H.B. SARS-CoV-2 infection: The role of PD-1/PD-L1 and CTLA-4 axis. Life Sci. 2021, 270, 119124. [Google Scholar] [CrossRef]
- Parackova, Z.; Zentsova, I.; Bloomfield, M.; Vrabcova, P.; Smetanova, J.; Klocperk, A.; Meseznikov, G.; Casas Mendez, L.F.; Vymazal, T.; Sediva, A. Disharmonic Inflammatory Signatures in COVID-19: Augmented Neutrophils’ but Impaired Monocytes’ and Dendritic Cells’ Responsiveness. Cells 2020, 9, 2206. [Google Scholar] [CrossRef]
- Christensen, E.E.; Jorgensen, M.J.; Nore, K.G.; Dahl, T.B.; Yang, K.; Ranheim, T.; Huse, C.; Lind, A.; Nur, S.; Stiksrud, B.; et al. Critical COVID-19 is associated with distinct leukocyte phenotypes and transcriptome patterns. J. Intern. Med. 2021, 290, 677–692. [Google Scholar] [CrossRef]
- El Sehmawy, A.A.; Abdul-Mohymen, A.M.; Seliem, N.; Elamir, R.Y.; Ibrahim, H.F.; Mahmoud, N.A.; Abdou, A.E. Study of Monocyte Subsets and Their Surface Expression of CD86 and Serum IL-17 Compared to Serum Procalcitonin as Markers of Early Neonatal Sepsis. Infect. Drug Resist. 2021, 14, 5375–5382. [Google Scholar] [CrossRef] [PubMed]
- Varchetta, S.; Mele, D.; Oliviero, B.; Mantovani, S.; Ludovisi, S.; Cerino, A.; Bruno, R.; Castelli, A.; Mosconi, M.; Vecchia, M.; et al. Unique immunological profile in patients with COVID-19. Cell Mol. Immunol. 2021, 18, 604–612. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wu, K.; Tao, K.; Chen, L.; Zheng, Q.; Lu, X.; Liu, J.; Shi, L.; Liu, C.; Wang, G.; et al. Tim-3/galectin-9 signaling pathway mediates T-cell dysfunction and predicts poor prognosis in patients with hepatitis B virus-associated hepatocellular carcinoma. Hepatology 2012, 56, 1342–1351. [Google Scholar] [CrossRef] [PubMed]
- Carter, M.J.; Fish, M.; Jennings, A.; Doores, K.J.; Wellman, P.; Seow, J.; Acors, S.; Graham, C.; Timms, E.; Kenny, J.; et al. Peripheral immunophenotypes in children with multisystem inflammatory syndrome associated with SARS-CoV-2 infection. Nat. Med. 2020, 26, 1701–1707. [Google Scholar] [CrossRef] [PubMed]
- Arunachalam, P.S.; Wimmers, F.; Mok, C.K.P.; Perera, R.; Scott, M.; Hagan, T.; Sigal, N.; Feng, Y.; Bristow, L.; Tak-Yin Tsang, O.; et al. Systems biological assessment of immunity to mild versus severe COVID-19 infection in humans. Science 2020, 369, 1210–1220. [Google Scholar] [CrossRef]





| COVID-19 Patients n = 55 | |
|---|---|
| Sex: f/m (n) | 21/34 |
| Age (mean ± SD years) | 58 ± 15 |
| Women (mean ± SD years) | 61 ± 14 |
| Men (mean ± SD years) | 59 ± 13 |
| Clinical symptoms (%) (no/yes) | |
| - fever | 12.7/87.3 |
| - cough | 23.6/76.4 |
| - dyspnea | 25.4/74.6 |
| - respiratory failure | 78.1/21.9 |
| Diseases comorbidities (%) (no/yes) | |
| - diabetes | 78.1/21.9 |
| - hypertension | 50.9/49.1 |
| - obesity | 81.8/18.2 |
| - coronary heart disease | 81.8/18.2 |
| - neoplastic diseases | 90.9/9.1 |
| Saturation (mean ± SD years) | 90.0 ± 6.5 |
| Chest X-ray changes (%) (no/yes) | 14.5/85.5 |
| Oxygen supplementation (%) (no/yes) | 7.2/92.8 |
| Invasive ventilation (%) (no/yes) | 94.5/5.5 |
| WBC and Study Subpopulation [k/µL] | Patients with COVID-19 Median (Q1–Q3) | Convalescent Median (Q1–Q3) | * p < 0.05 Mann–Whitney U Test |
|---|---|---|---|
| WBC | 7000 (4640–9020) | 7930 (6600–10,470) | * 0.024360 |
| Lymphocytes | 1025 (710–1570) | 1662 (1170–2199) | * 0.000014 |
| T Lymphocytes | 680 (422–1103) | 1271 (826–1607) | * 0.000005 |
| CD4 cells | 461 (271–675) | 775 (559–1117) | * 0.000047 |
| CD8 cells | 204 (130–403) | 439 (238–567) | * 0.000133 |
| Ratio CD4/CD8 | 2.0 (1.3–2.9) | 2.1 (1.2–2.8) | 0.825576 |
| B Lymphocytes | 120 (64–216) | 172 (120–281) | * 0.016123 |
| NK cells | 146 (79–253) | 178 (74–300) | 0.434570 |
| Neutrophils | 5143 (3192–7941) | 5319 (4135–7581) | 0.457164 |
| Eosinophils | 8 (0–38) | 72 (18–190) | * 0.000006 |
| Basophils | 9 (4–21) | 24 (8–54) | * 0.005186 |
| Monocytes | 377 (260–454) | 536 (399–815) | * 0.000020 |
| % of all leukocytes | |||
| Lymphocytes | 14,7 (9.1–28.2) | 22.4 (12.8–30.5) | * 0.044030 |
| T Lymphocytes | 10.5 (5.5–20.7) | 16.9 (8.9–22.2) | * 0.013242 |
| CD4 cells | 5.6 (3.5–13.3) | 10.5 (5.4–14.7) | * 0.016998 |
| CD8 cells | 3.5 (1.8–6.1) | 4.9 (3.4–7.1) | * 0.046088 |
| B Lymphocytes | 1.7 (1.0–2.6) | 2.1 (1.4–3.2) | 0.225750 |
| NK cells | 2.5 (1.1–4.1) | 2.0 (1.0–3.9) | 0.492249 |
| Neutrophils | 79.4 (63.4–86.9) | 68.8 (60.1–79.1) | * 0.015562 |
| Eosinophils | 0.1 (0.0–0.8) | 1.1 (0.2–2.4) | * 0.000093 |
| Basophils | 0.1 (0.1–0.3) | 0.3 (0.1–0.6) | 0.060189 |
| Monocytes | 5.5 (3.8–7.6) | 7.0 (4.6–9.1) | * 0.037130 |
| Patients with COVID-19 Median (Q1–Q3) | Convalescent Median (Q1–Q3) | * p < 0.05 Mann–Whitney U Test | |
|---|---|---|---|
| All monocytes [% of leukocytes] | 5.5 (3.8–7.6) | 7.0 (4.6–9.1) | * 0.037130 |
| [% of monocytes] | |||
| Classical monocytes CD14++ CD16- | 83.2 (76.5–87.1) | 84.2 (76.9–88.8) | 0.460992 |
| Intermediate monocytes CD14+ CD16+ | 7.5 (4.4–15.7) | 7.7 (5.6–11.7) | 0.904777 |
| Non classical monocytes CD14−/+ CD16++ | 0.9 (0.3–1.8) | 2.2 (1.1–4.3) | * 0.000098 |
| PD-L1+ monocytes [%] | 33.8 (22.4–52.5) | 29.6 (15.7–60.3) | 0.659322 |
| PD-L1+ classical monocytes [%] | 18.1 (11.4–34.3) | 15.5 (7.5–46.3) | 0.659322 |
| PD-L1+ intermediate monocytes [%] | 73.1 (52.1–84.4) | 57.6 (38.4–71.9) | * 0.007261 |
| PD-L1+ non-classical monocytes [%] | 75.0 (62.5–85.7) | 77.8 (64.8–86.2) | 0.673065 |
| PD-L1+ monocytes [GeoMean] | 310 (247–435) | 276 (194–476) | 0.391353 |
| PD-L1+ classical monocytes [GeoMean] | 288 (222–412) | 237 (166–477) | 0.228186 |
| PD-L1+ intermediate monocytes [GeoMean] | 669 (523–919) | 530 (407–723) | * 0.015287 |
| PD-L1+ non-classical monocytes [GeoMean] | 728 (600–909) | 692 (558–832) | 0.427183 |
| Patients with COVID-19 Median (Q1–Q3) | Convalescent Median (Q1–Q3) | * p < 0.05 Mann–Whitney U Test | |
|---|---|---|---|
| CD62L+ monocytes [%] | 83.0 (72.4–91.3) | 69.6 (40.1–88.2) | * 0.000107 |
| TIM-3+ monocytes [%] | 90.2 (83.3–94.8) | 90.6 (86.3–93.2) | 0.855115 |
| CD86+ monocytes [%] | 93.7 (82.7–97.3) | 92.7 (87.3–95.7) | 0.387873 |
| CD62L+ monocytes [GeoMean] | 17,970 (9645–26,204) | 11,362 (5154–23,498) | * 0.020923 |
| TIM-3+ monocytes [GeoMean] | 2333 (1715–3197) | 2141 (1706–2638) | 0.172704 |
| CD86+ monocytes [GeoMean] | 724 (483–1188) | 725 (539–886) | 0.618789 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rutkowska, E.; Kwiecień, I.; Kłos, K.; Rzepecki, P.; Chciałowski, A. Intermediate Monocytes with PD-L1 and CD62L Expression as a Possible Player in Active SARS-CoV-2 Infection. Viruses 2022, 14, 819. https://doi.org/10.3390/v14040819
Rutkowska E, Kwiecień I, Kłos K, Rzepecki P, Chciałowski A. Intermediate Monocytes with PD-L1 and CD62L Expression as a Possible Player in Active SARS-CoV-2 Infection. Viruses. 2022; 14(4):819. https://doi.org/10.3390/v14040819
Chicago/Turabian StyleRutkowska, Elżbieta, Iwona Kwiecień, Krzysztof Kłos, Piotr Rzepecki, and Andrzej Chciałowski. 2022. "Intermediate Monocytes with PD-L1 and CD62L Expression as a Possible Player in Active SARS-CoV-2 Infection" Viruses 14, no. 4: 819. https://doi.org/10.3390/v14040819