
Identification of Unique and Conserved Neutralizing Epitopes of Vestigial Esterase Domain in HA Protein of the H9N2 Subtype of Avian Influenza Virus


Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus
2.2. Cells
2.3. HA Gene Cloning and Recombinant Protein Expression
2.4. Mice Immunization
2.5. Cell Fusion and Identification of Monoclonal Antibodies
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Design and Expression of the Truncated HA Gene
2.8. SDS-PAGE and Western Blotting
2.9. Indirect Immunofluorescence Assay (IFA)
2.10. Biological Information Analysis
2.11. Neutralisation Assay
2.12. Haemagglutinin Inhibition (HI) Assay
2.13. Quantitative Real-Time PCR
3. Results
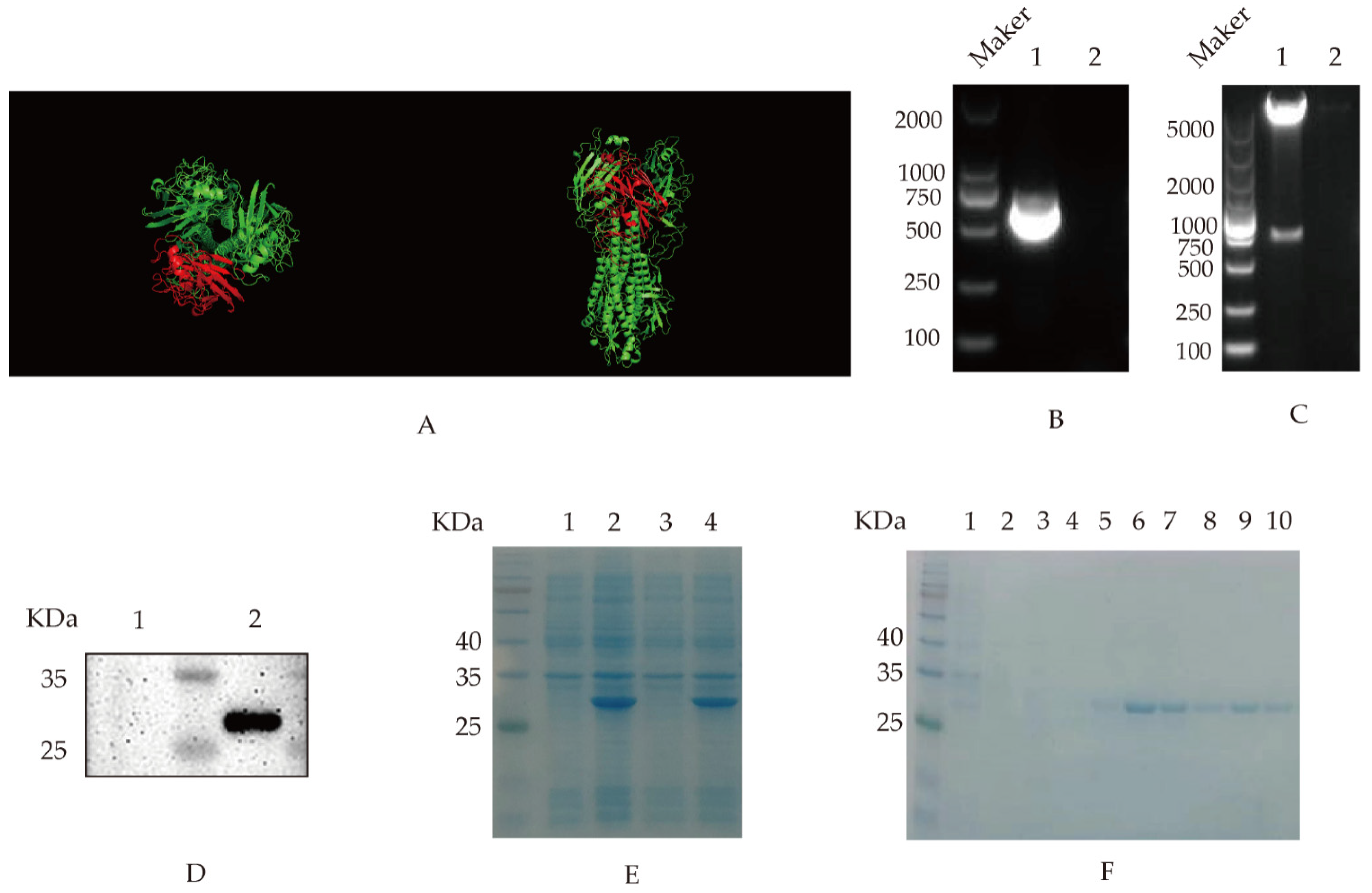
3.1. Cloning, Expression and Purification of Recombinant HA Protein
3.2. Screen of Monoclonal Antibody Specific to HA Proteins
3.3. Screening of B-Cell Determinants Specific to the Monoclonal Antibody
3.4. Haemagglutinin Inhibition and Neutralization Assay
3.5. The Identified Epitope Is Highly Conserved in AIV Strains
3.6. Spatial Location Prediction of Epitope Binding
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, Y.; Hu, S.; Bai, T.; Yang, L.; Zhao, X.; Zhu, W.; Huang, Y.; Deng, Z.; Zhang, H.; Bai, Z.; et al. Phylogenetic and antigenic characterization of reassortant H9N2 avian influenza viruses isolated from wild waterfowl in the East Dongting Lake wetland in 2011–2012. Virol. J. 2014, 11, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonas, M.; Sahesti, A.; Murwijati, T.; Lestariningsih, C.L.; Irine, I.; Ayesda, C.S.; Prihartini, W.; Mahardika, G.N. Identification of avian influenza virus subtype H9N2 in chicken farms in Indonesia. Prev. Vet. Med. 2018, 159, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.R.; Park, C.K.; Oem, J.K.; Bae, Y.C.; Choi, J.G.; Lee, O.S.; Lee, Y.J. Characterization of H5N2 influenza viruses isolated in South Korea and their influence on the emergence of a novel H9N2 influenza virus. J. Gen. Virol. 2010, 91 Pt 8, 1978–1983. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Yu, K.; Tian, G.; Yu, D.; Liu, L.; Jing, B.; Ping, J.; Chen, H. Evolution of H9N2 influenza viruses from domestic poultry in Mainland China. Virology 2005, 340, 70–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Wang, Y.; Zhao, C.; Gao, X.; Zhang, Y.; Li, J.; Wang, M.; Zhang, H.; Liu, W.; Wang, C.; et al. Molecular characterization, receptor binding property, and replication in chickens and mice of H9N2 avian influenza viruses isolated from chickens, peafowls, and wild birds in eastern China. Emerg. Microbes Infect. 2021, 10, 2098–2112. [Google Scholar] [CrossRef]
- Homme, P.J.; Easterday, B.C. Avian influenza virus infections. I. Characteristics of influenza A-turkey-Wisconsin-1966 virus. Avian Dis. 1970, 14, 66–74. [Google Scholar] [CrossRef]
- Chen, F.; Xia, C. Cloning and Analysis of Avian Influenza Nucleoprotein Gene from A/Chicken/Beijing/1/96(H9N2). Chin. J. Prev. Vet. Med. 1999, 21, 51+53–54. [Google Scholar]
- Gao, X.; Wang, N.; Chen, Y.; Gu, X.; Huang, Y.; Liu, Y.; Jiang, F.; Bai, J.; Qi, L.; Xin, S.; et al. Sequence characteristics and phylogenetic analysis of H9N2 subtype avian influenza A viruses detected from poultry and the environment in China, 2018. PeerJ 2021, 9, e12512. [Google Scholar] [CrossRef]
- Jin, X.; Zha, Y.; Hu, J.; Li, X.; Chen, J.; Xie, S.; Dai, Y.; Li, Z.; Wang, X.; Wang, F.; et al. New molecular evolutionary characteristics of H9N2 avian influenza virus in Guangdong Province, China. Infect. Genet. Evol. 2020, 77, 104064. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Liu, J. H9N2 influenza virus in China: A cause of concern. Protein Cell 2015, 6, 18–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Tang, Y.; Liu, X.; Peng, D.; Liu, W.; Liu, H.; Lu, S.; Liu, X. Characterization of H9N2 influenza viruses isolated from vaccinated flocks in an integrated broiler chicken operation in eastern China during a 5 year period (1998–2002). J. Gen. Virol. 2008, 89 Pt 12, 3102–3112. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Qin, K. Human-infecting influenza A (H9N2) virus: A forgotten potential pandemic strain? Zoonoses Public Health 2020, 67, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Peiris, M.; Yuen, K.Y.; Leung, C.W.; Chan, K.H.; Ip, P.L.; Lai, R.W.; Orr, W.K.; Shortridge, K.F. Human infection with influenza H9N2. Lancet 1999, 354, 916–917. [Google Scholar] [CrossRef] [PubMed]
- Butt, K.M.; Smith, G.J.; Chen, H.; Zhang, L.J.; Leung, Y.H.; Xu, K.M.; Lim, W.; Webster, R.G.; Yuen, K.Y.; Peiris, J.S.; et al. Human infection with an avian H9N2 influenza A virus in Hong Kong in 2003. J. Clin. Microbiol. 2005, 43, 5760–5767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.H.; Zhou, X.Z.; Li, M.X. Discoveries of avian influenza A(H9N2) virus in chickens and men infected by H9N2 virus in Guangzhou area. Zhonghua Shi Yan He Lin Chuang Bing Du Xue Za Zhi 2004, 18, 213–214. [Google Scholar] [PubMed]
- Gu, M.; Chen, H.; Li, Q.; Huang, J.; Zhao, M.; Gu, X.; Jiang, K.; Wang, X.; Peng, D.; Liu, X. Enzootic genotype S of H9N2 avian influenza viruses donates internal genes to emerging zoonotic influenza viruses in China. Vet. Microbiol. 2014, 174, 309–315. [Google Scholar] [CrossRef] [PubMed]
- RahimiRad, S.; Alizadeh, A.; Alizadeh, E.; Hosseini, S.M. The avian influenza H9N2 at avian-human interface: A possible risk for the future pandemics. J. Res. Med. Sci. 2016, 21, 51. [Google Scholar]
- Shen, Y.Y.; Ke, C.W.; Li, Q.; Yuan, R.Y.; Xiang, D.; Jia, W.X.; Yu, Y.D.; Liu, L.; Huang, C.; Qi, W.B.; et al. Novel Reassortant Avian Influenza A(H5N6) Viruses in Humans, Guangdong, China, 2015. Emerg. Infect. Dis. 2016, 22, 1507–1509. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, R.; Jiang, L.; Xiong, C.; Chen, Y.; Zhao, G.; Jiang, Q. The complexity of human infected AIV H5N6 isolated from China. BMC Infect. Dis. 2016, 16, 600. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Liu, B.; Ma, S.; Cui, P.; Liu, W.; Li, Y.; Guo, J.; Chen, H. High frequency of reassortment after co-infection of chickens with the H4N6 and H9N2 influenza A viruses and the biological characteristics of the reassortants. Vet. Microbiol. 2018, 222, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Coughlan, L.; Palese, P. Overcoming Barriers in the Path to a Universal Influenza Virus Vaccine. Cell Host Microbe 2018, 24, 18–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosik, I.; Yewdell, J.W. Influenza Hemagglutinin and Neuraminidase: Yin(-)Yang Proteins Coevolving to Thwart Immunity. Viruses 2019, 11, 346. [Google Scholar] [CrossRef]
- Kuraoka, M.; Adachi, Y.; Takahashi, Y. Hide and seek: Interplay between influenza viruses and B cells. Int. Immunol. 2020, 32, 605–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.K.; Rappuoli, R.; Xu, X.N. Correlates of protection against influenza infection in humans—On the path to a universal vaccine? Curr. Opin. Immunol. 2013, 25, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Gu, C.; Zeng, X.; Song, Y.; Li, Y.; Liu, L.; Kawaoka, Y.; Zhao, D.; Chen, H. Glycosylation and an amino acid insertion in the head of hemagglutinin independently affect the antigenic properties of H5N1 avian influenza viruses. Sci. China Life Sci. 2019, 62, 76–83. [Google Scholar] [CrossRef]
- Zhao, D.; Liang, L.; Wang, S.; Nakao, T.; Li, Y.; Liu, L.; Guan, Y.; Fukuyama, S.; Bu, Z.; Kawaoka, Y.; et al. Glycosylation of the Hemagglutinin Protein of H5N1 Influenza Virus Increases Its Virulence in Mice by Exacerbating the Host Immune Response. J. Virol. 2017, 91, e02215-16. [Google Scholar] [CrossRef] [Green Version]
- Shen, W.; Wang, Q.; Wang, Z.; Liu, M.; Du, Y.; Yuan, L.; Han, L.; Smietanka, K.; Chen, H.; Xu, S.; et al. Specific Monoclonal Antibodies Targeting Unique HA Epitopes Block H7N9 Influenza A Viral Replication. J. Virol. 2022, 96, e0123822. [Google Scholar] [CrossRef] [PubMed]
- Angeletti, D.; Gibbs, J.S.; Angel, M.; Kosik, I.; Hickman, H.D.; Frank, G.M.; Das, S.R.; Wheatley, A.K.; Prabhakaran, M.; Leggat, D.J.; et al. Defining B cell immunodominance to viruses. Nat. Immunol. 2017, 18, 456–463. [Google Scholar] [CrossRef]
- Caton, A.J.; Brownlee, G.G.; Yewdell, J.W.; Gerhard, W. The antigenic structure of the influenza virus A/PR/8/34 hemagglutinin (H1 subtype). Cell 1982, 31 Pt 1, 417–427. [Google Scholar] [CrossRef]
- Thyagarajan, B.; Bloom, J.D. The inherent mutational tolerance and antigenic evolvability of influenza hemagglutinin. Elife 2014, 3, e03300. [Google Scholar] [CrossRef] [PubMed]
- Wiley, D.C.; Wilson, I.A.; Skehel, J.J. Structural identification of the antibody-binding sites of Hong Kong influenza haemagglutinin and their involvement in antigenic variation. Nature 1981, 289, 373–378. [Google Scholar] [CrossRef]
- Padilla-Quirarte, H.O.; Lopez-Guerrero, D.V.; Gutierrez-Xicotencatl, L.; Esquivel-Guadarrama, F. Protective Antibodies Against Influenza Proteins. Front. Immunol. 2019, 10, 1677. [Google Scholar] [CrossRef]
- Yassine, H.M.; Boyington, J.C.; McTamney, P.M.; Wei, C.J.; Kanekiyo, M.; Kong, W.P.; Gallagher, J.R.; Wang, L.; Zhang, Y.; Joyce, M.G.; et al. Hemagglutinin-stem nanoparticles generate heterosubtypic influenza protection. Nat. Med. 2015, 21, 1065–1070. [Google Scholar] [CrossRef]
- Oh, H.L.; Akerstrom, S.; Shen, S.; Bereczky, S.; Karlberg, H.; Klingstrom, J.; Lal, S.K.; Mirazimi, A.; Tan, Y.J. An antibody against a novel and conserved epitope in the hemagglutinin 1 subunit neutralizes numerous H5N1 influenza viruses. J. Virol. 2010, 84, 8275–8286. [Google Scholar] [CrossRef] [Green Version]
- Ren, H.; Wang, G.; Wang, S.; Chen, H.; Chen, Z.; Hu, H.; Cheng, G.; Zhou, P. Cross-protection of newly emerging HPAI H5 viruses by neutralizing human monoclonal antibodies: A viable alternative to oseltamivir. MAbs 2016, 8, 1156–1166. [Google Scholar] [CrossRef] [Green Version]
- Tan, G.S.; Leon, P.E.; Albrecht, R.A.; Margine, I.; Hirsh, A.; Bahl, J.; Krammer, F. Broadly-Reactive Neutralizing and Non-neutralizing Antibodies Directed against the H7 Influenza Virus Hemagglutinin Reveal Divergent Mechanisms of Protection. PLoS Pathog. 2016, 12, e1005578. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Teo, S.H.C.; Arularasu, S.C.; Liu, Z.; Mohd-Ismail, N.K.; Mok, C.K.; Ong, C.B.; Chu, J.J.; Tan, Y.J. Contribution of Fc-dependent cell-mediated activity of a vestigial esterase-targeting antibody against H5N6 virus infection. Emerg. Microbes Infect. 2020, 9, 95–110. [Google Scholar] [CrossRef]
- Bangaru, S.; Zhang, H.; Gilchuk, I.M.; Voss, T.G.; Irving, R.P.; Gilchuk, P.; Matta, P.; Zhu, X.; Lang, S.; Nieusma, T.; et al. A multifunctional human monoclonal neutralizing antibody that targets a unique conserved epitope on influenza HA. Nat. Commun. 2018, 9, 2669. [Google Scholar] [CrossRef] [Green Version]
- Velkov, T.; Ong, C.; Baker, M.A.; Kim, H.; Li, J.; Nation, R.L.; Huang, J.X.; Cooper, M.A.; Rockman, S. The antigenic architecture of the hemagglutinin of influenza H5N1 viruses. Mol. Immunol. 2013, 56, 705–719. [Google Scholar] [CrossRef]
- Ren, H.; Zhou, P. Epitope-focused vaccine design against influenza A and B viruses. Curr. Opin. Immunol. 2016, 42, 83–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Z.; Paul, S.S.; Mo, X.; Yuan, Y.A.; Tan, Y.J. The Vestigial Esterase Domain of Haemagglutinin of H5N1 Avian Influenza A Virus: Antigenicity and Contribution to Viral Pathogenesis. Vaccines 2018, 6, 53. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Yang, D.; Ren, Q.; Yang, Y.; Liu, X.; Xu, X.; Liu, W.; Chen, S.; Peng, D.; Liu, X. Identification and characterization of a novel antigenic epitope in the hemagglutinin of the escape mutants of H9N2 avian influenza viruses. Vet. Microbiol. 2015, 178, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Wan, Z.; Ye, J.; Xu, L.; Shao, H.; Jin, W.; Qian, K.; Wan, H.; Qin, A. Antigenic mapping of the hemagglutinin of an H9N2 avian influenza virus reveals novel critical amino acid positions in antigenic sites. J. Virol. 2014, 88, 3898–3901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Li, Z.; Su, X.; Qiao, Y.; Fan, W.; Li, H.; Shi, B.; Qin, T.; Chen, S.; Peng, D.; et al. Enhanced cross-lineage protection induced by recombinant H9N2 avian influenza virus inactivated vaccine. Vaccine 2019, 37, 1736–1742. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Guo, Y.H.; Jiang, T.; Wang, Y.D.; Chan, K.H.; Li, X.F.; Yu, W.; McBride, R.; Paulson, J.C.; Yuen, K.Y.; et al. A unique and conserved neutralization epitope in H5N1 influenza viruses identified by an antibody against the A/Goose/Guangdong/1/96 hemagglutinin. J. Virol. 2013, 87, 12619–12635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.N.; Zheng, Y.; Hao, S.S.; Zhang, Z.; Cai, J.X.; Zong, M.M.; Feng, X.L.; Liu, Q.T. The molecular evolutionary characteristics of new isolated H9N2 AIV from East China and the function of vimentin on virus replication in MDCK cells. Virol. J. 2020, 17, 78. [Google Scholar] [CrossRef]
- Huang, X.; Huang, J.; Yin, G.; Cai, Y.; Chen, M.; Hu, J.; Feng, X. Identification of NP Protein-Specific B-Cell Epitopes for H9N2 Subtype of Avian Influenza Virus. Viruses 2022, 14, 1172. [Google Scholar] [CrossRef]
- Danilenko, D.M.; Eropkin, M.Y.; Leonenko, V.N.; Konovalova, N.I.; Petrova, P.A.; Zheltukhina, A.I.; Vassilieva, A.D. Assessment of rat polyclonal antisera’s suitability in hemagglutination inhibition assay for influenza surveillance and antigenic mapping. J. Virol. Methods 2021, 293, 114170. [Google Scholar] [CrossRef]
- Schulze, I.T. Effects of glycosylation on the properties and functions of influenza virus hemagglutinin. J. Infect. Dis. 1997, 176 (Suppl. 1), S24–S28. [Google Scholar] [CrossRef] [Green Version]
- Moremen, K.W.; Tiemeyer, M.; Nairn, A.V. Vertebrate protein glycosylation: Diversity, synthesis and function. Nat. Rev. Mol. Cell Biol. 2012, 13, 448–462. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Zhao, L.; Guo, Y.; Zhao, Y.; Li, Y.; Chen, N.; Lu, Y.; Yu, M.; Deng, L.; Ping, J. Antigenic Evolution Characteristics and Immunological Evaluation of H9N2 Avian Influenza Viruses from 1994–2019 in China. Viruses 2022, 14, 726. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.F.; Lai, H.Z.; Li, L.; Liu, Y.P.; Zhang, W.Y.; Gao, R.; Huang, W.K.; Luo, Q.F.; Gao, Y.; Luo, Q.; et al. Endemic Variation of H9N2 Avian Influenza Virus in China. Avian Dis. 2016, 60, 817–825. [Google Scholar] [CrossRef] [PubMed]
- DuBois, R.M.; Zaraket, H.; Reddivari, M.; Heath, R.J.; White, S.W.; Russell, C.J. Acid stability of the hemagglutinin protein regulates H5N1 influenza virus pathogenicity. PLoS Pathog. 2011, 7, e1002398. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Name | Primer Sequence (5′–3′) | Primer Size | Tm (°C) | Product Size (bp) |
|---|---|---|---|---|---|
| HA | HA-F | CGCGGATCCATTGAAGGACTGATCTATGGCAACC | 34 | 73.4 | 648 |
| HA-R | CCCAAGCTTACATTGCACTACACAGTTACCACTG | 34 | 73.4 | ||
| HA-75 | HA-74-F | CGCGGATCCATTGAAGGACTGATCTATGGCAACC | 34 | 74 | 414 |
| HA-74-R | ACGCGTCGACCACACTTGTTGTTGTGTCGGTCCTT | 35 | 73 | ||
| HA-159 | HA-158-F | CGCGGATCCATGAGATGGCTGACTCAAAAGAACA | 34 | 70.8 | 396 |
| HA-158-R | ACGCGTCGACACATTGCACTACACAGTTACCACTG | 35 | 69 | ||
| HA-100 | HA-99-F | CGCGGATCCAGACCATCGGCCGTTAATGGAATGT | 34 | 68.8 | 246 |
| HA-99-R | ACGCGTCGACTCCTCTATTATTTGTGTATTGGGCG | 35 | 62.1 | ||
| HA-131 | HA-130-F | CCGGAATTCCGGATGATAAAGCGAGGGATCAACG | 34 | 69.8 | 192 |
| HA-130-R | ACGCGTCGACTCGCACTTGATCCATCATTGCTCTT | 35 | 67 |
| Gene Name | Primer Name | Primer Sequence (5′–3′) | Primer Size | Product Size (bp) |
|---|---|---|---|---|
| NA | NA-F | TCGGCGACACACCAAGAAATGATG | 24 | 143 |
| NA-R | ATCCACTTTGATCGTTCGTCCCATC | 24 | ||
| GAPDH | GAPDH-F | AATTCCACGGCACAGTCAAGGC | 22 | 124 |
| GAPDH-R | ACAACATACTCAGCACCAGCATCAC | 25 |
| mAbs | Antibody Potency of Supernatant | Ascites Potency | Heavy Chain | Light Chain |
|---|---|---|---|---|
| 1E11 | 1:2000 | 1:20,000 | IgG2b | κ |
| 2B3 | 1:100,000 | 1:1,280,000 | IgG1 | κ |
| 3E5 | 1:2000 | 1:40,000 | IgG1 | κ |
| 3H3 | 1:20,000 | 1:640,000 | IgG2b | κ |
| 4F1 | 1:2000 | 1:20,000 | IgG1 | κ |
| 5E10 | 1:10,000 | 1:80,000 | IgG1 | κ |
| Vector Truncation | mAbs | |||||
|---|---|---|---|---|---|---|
| 1E11 | 2B3 | 3E5 | 3H3 | 4F1 | 5E10 | |
| HA-75 | + | + | + | + | + | + |
| HA-159 | - | - | + | - | + | - |
| HA-100 | + | + | + | + | + | + |
| HA-131 | - | - | + | - | + | - |
| Peptide HA-100 | - | - | - | - | - | - |
| Peptide HA-106 | - | - | - | - | - | - |
| Peptide HA-111 | + | - | - | + | - | - |
| Peptide HA-116 | - | + | - | - | - | + |
| Peptide HA-121 | - | - | - | - | - | - |
| Peptide HA-159 | - | - | - | - | - | - |
| Peptide HA-165 | - | - | + | - | + | - |
| Peptide HA-170 | - | - | + | - | + | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, X.; Yin, G.; Cai, Y.; Hu, J.; Huang, J.; Liu, Q.; Feng, X. Identification of Unique and Conserved Neutralizing Epitopes of Vestigial Esterase Domain in HA Protein of the H9N2 Subtype of Avian Influenza Virus. Viruses 2022, 14, 2739. https://doi.org/10.3390/v14122739
Huang X, Yin G, Cai Y, Hu J, Huang J, Liu Q, Feng X. Identification of Unique and Conserved Neutralizing Epitopes of Vestigial Esterase Domain in HA Protein of the H9N2 Subtype of Avian Influenza Virus. Viruses. 2022; 14(12):2739. https://doi.org/10.3390/v14122739
Chicago/Turabian StyleHuang, Xiangyu, Guihu Yin, Yiqin Cai, Jianing Hu, Jingwen Huang, Qingtao Liu, and Xiuli Feng. 2022. "Identification of Unique and Conserved Neutralizing Epitopes of Vestigial Esterase Domain in HA Protein of the H9N2 Subtype of Avian Influenza Virus" Viruses 14, no. 12: 2739. https://doi.org/10.3390/v14122739