

Emergence of High Pathogenicity Avian Influenza Virus H5N1 Clade 2.3.4.4b in Wild Birds and Poultry in Botswana

 , , , , and add
Show full author list
, , , , and add
Show full author list
Abstract
:1. Introduction
2. Materials and Methods
2.1. Surveillance Activities within Botswana
2.2. Pathological Assessment of Submitted Carcasses
2.3. Virological Investigation
3. Results and Discussion
4. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Adlhoch, C.; Fusaro, A.; Gonzales, J.L.; Kuiken, T.; Marangon, S.; Niqueux; Staubach, C.; Terregino, C.; Aznar, I.; Guajardo, I.M.; et al. Avian influenza overview December 2021–March 2022. EFSA J. 2022, 20, e07289. [Google Scholar] [CrossRef] [PubMed]
- Oliver, I.; Roberts, J.; Brown, C.S.; Byrne, A.M.; Mellon, D.; Hansen, R.; Banyard, A.C.; James, J.; Donati, M.; Porter, R.; et al. A case of avian influenza A(H5N1) in England, January 2022. Eur. Commun. Dis. Bull. 2022, 27, 2200061. [Google Scholar] [CrossRef] [PubMed]
- Pyankova, O.G.; Susloparov, I.M.; Moiseeva, A.A.; Kolosova, N.P.; Onkhonova, G.S.; Danilenko, A.V.; Vakalova, E.V.; Shendo, G.L.; Nekeshina, N.N.; Noskova, L.N.; et al. Isolation of clade 2.3.4.4b A(H5N8), a highly pathogenic avian influenza virus, from a worker during an outbreak on a poultry farm, Russia, December 2020. Eur. Commun. Dis. Bull. 2021, 26, 2100439. [Google Scholar] [CrossRef]
- Pohlmann, A.; King, J.; Fusaro, A.; Zecchin, B.; Banyard, A.C.; Brown, I.H.; Byrne, A.M.P.; Beerens, N.; Liang, Y.; Heutink, R.; et al. Has Epizootic Become Enzootic? Evidence for a Fundamental Change in the Infection Dynamics of Highly Pathogenic Avian Influenza in Europe, 2021. mBio 2022, 13, e0060922. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority; European Centre for Disease Prevention and Control and European Union Reference Laboratory for Avian Influenza; Adlhoch, C.; Fusaro, A.; Gonzales, J.L.; Kuiken, T.; Marangon, S.; Niqueux, É.; Staubach, C.; Terregino, C.; et al. Avian influenza overview December 2020–February 2021. EFSA J. 2021, 19, e06497. [Google Scholar] [CrossRef]
- FAO. Sub-Saharan Africa HPAI Situation Update. Available online: https://www.fao.org/animal-health/situation-updates/sub-saharan-africa-hpai/ (accessed on 10 February 2022).
- The Animal and Plant Health Agency. High Pathogenicity Avian Influenza H5N8 and H5N1 Outbreaks in Great Britain: November 2020 to April 2021; The Animal and Plant Health Agency: Singapore, 2021. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/1011519/aiv-hpai-h5n8-h5n1-epi-rept-1120-0421.pdf (accessed on 1 June 2022).
- Khomenko, S.; Abolnik, C.; Roberts, L.; Waller, L.; Shaw, K.; Monne, I.; Taylor, J.; Dhingra, M.; Claudia, P.; Mugyeom, M.; et al. 2016–2018 Spread of H5N8 Highly Pathogenic Avian Influenza (HPAI) in Sub-Saharan Africa: Epidemiological and Ecological Observations; FOCUS ON, No. 12, Aug 2018, Rome; Food and Agriculture Organization: Québec City, QC, Canada, 2018. [Google Scholar]
- Fusaro, A.; Zecchin, B.; Vrancken, B.; Abolnik, C.; Ademun, R.; Alassane, A.; Arafa, A.; Awuni, J.A.; Couacy-Hymann, E.; Coulibaly, M.B.; et al. Disentangling the role of Africa in the global spread of H5 highly pathogenic avian influenza. Nat. Commun. 2019, 10, 5310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makalo, M.R.J.; Dundon, W.G.; Settypalli, T.B.K.; Datta, S.; Lamien, C.E.; Cattoli, G.; Phalatsi, M.S.; Lepheana, R.J.; Matlali, M.; Mahloane, R.G.; et al. Highly pathogenic avian influenza (A/H5N1) virus outbreaks in Lesotho, May 2021. Emerg. Microbes Infect. 2022, 11, 757–760. [Google Scholar] [CrossRef]
- Department of Agriculture. Avian Influenza: H5 and H7 Outbreak Update Report; Department of Agriculture: Quezon City, Philippines, 2022.
- Lewis, N.S.; Banyard, A.C.; Whittard, E.; Karibayev, T.; Al Kafagi, T.; Chvala, I.; Byrne, A.; Meruyert Akberovna, S.; King, J.; Harder, T.; et al. Emergence and spread of novel H5N8, H5N5 and H5N1 clade 2.3.4.4 highly pathogenic avian influenza in 2020. Emerg. Microbes Infect. 2021, 10, 148–151. [Google Scholar] [CrossRef]
- WOAH. Avian Influenza. Available online: https://www.woah.org/en/disease/avian-influenza/ (accessed on 15 November 2021).
- Banyard, A.C.; Lean, F.Z.X.; Robinson, C.; Howie, F.; Tyler, G.; Nisbet, C.; Seekings, J.; Meyer, S.; Whittard, E.; Ashpitel, H.F.; et al. Detection of Highly Pathogenic Avian Influenza Virus H5N1 Clade 2.3.4.4b in Great Skuas: A Species of Conservation Concern in Great Britain. Viruses 2022, 14, 212. [Google Scholar] [CrossRef]
- Botswana Ministry of Agricultural Development and Food Security (Publisher). Botswana Ministry of Agricultural Development and Food Security 2020/2021 Annual Report; 2021. Available online: https://www.gov.bw/ministries/ministry-agricultural-development-and-food-security (accessed on 1 June 2022).
- Nagy, A.; Vostinakova, V.; Pirchanova, Z.; Cernikova, L.; Dirbakova, Z.; Mojzis, M.; Jirincova, H.; Havlickova, M.; Dan, A.; Ursu, K.; et al. Development and evaluation of a one-step real-time RT-PCR assay for universal detection of influenza A viruses from avian and mammal species. Arch. Virol. 2010, 155, 665–673. [Google Scholar] [CrossRef]
- Slomka, M.J.; Pavlidis, T.; Coward, V.J.; Voermans, J.; Koch, G.; Hanna, A.; Banks, J.; Brown, I.H. Validated RealTime reverse transcriptase PCR methods for the diagnosis and pathotyping of Eurasian H7 avian influenza viruses. Influenza Other Respir. Viruses 2009, 3, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Slomka, M.J.; Pavlidis, T.; Banks, J.; Shell, W.; McNally, A.; Essen, S.; Brown, I.H. Validated H5 Eurasian real-time reverse transcriptase-polymerase chain reaction and its application in H5N1 outbreaks in 2005–2006. Avian Dis. 2007, 51, 373–377. [Google Scholar] [CrossRef] [PubMed]
- James, J.; Slomka, M.J.; Reid, S.M.; Thomas, S.S.; Mahmood, S.; Byrne, A.M.P.; Cooper, J.; Russell, C.; Mollett, B.C.; Agyeman-Dua, E.; et al. Development and Application of Real-Time PCR Assays for Specific Detection of Contemporary Avian Influenza Virus Subtypes N5, N6, N7, N8, and N9. Avian Dis. 2019, 63, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Sutton, D.A.; Allen, D.P.; Fuller, C.M.; Mayers, J.; Mollett, B.C.; Londt, B.Z.; Reid, S.M.; Mansfield, K.L.; Brown, I.H. Development of an avian avulavirus 1 (AAvV-1) L-gene real-time RT-PCR assay using minor groove binding probes for application as a routine diagnostic tool. J. Virol. Methods 2019, 265, 9–14. [Google Scholar] [CrossRef] [PubMed]
- WOAH. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. version May 2022. Available online: https://www.woah.org/en/what-we-do/standards/codes-and-manuals/terrestrial-manual-online-access/ (accessed on 15 July 2022).
- Payungporn, S.; Chutinimitkul, S.; Chaisingh, A.; Damrongwantanapokin, S.; Buranathai, C.; Amonsin, A.; Theamboonlers, A.; Poovorawan, Y. Single step multiplex real-time RT-PCR for H5N1 influenza A virus detection. J. Virol. Methods 2006, 131, 143–147. [Google Scholar] [CrossRef]
- Nagy, A.; Černíková, L.; Kunteová, K.; Dirbáková, Z.; Thomas, S.S.; Slomka, M.J.; Dán, N.; Varga, T.; Máté, M.; Jiřincová, H.; et al. A universal RT-qPCR assay for “One Health” detection of influenza A viruses. PLoS ONE 2021, 16, e0244669. [Google Scholar] [CrossRef]
- James, J.; Seekings, A.H.; Skinner, P.; Purchase, K.; Mahmood, S.; Brown, I.H.; Hansen, R.D.E.; Banyard, A.C.; Reid, S.M. Rapid and sensitive detection of high pathogenicity Eurasian clade 2.3.4.4b avian influenza viruses in wild birds and poultry. J. Virol. Methods 2022, 301, 114454. [Google Scholar] [CrossRef]
- Commission, E. European Commision Decision 2006/437/EC of 4 August 2006 approving Diagnostic Manual for avian influenza as provided for in Council Directive 2005/94/EC of 4 August 2006 approving Diagnostic Manual for avian influenza as provided for in Council Directive 2005/94/EC. Off. J. Eur. Union 2006, 237, 1–27. [Google Scholar]
- World Organisation for Animal Health (WOAH). Terrestrial Manual: Avian Influenza (Infection with Avian Influenza Viruses). Available online: https://www.woah.org/fileadmin/Home/eng/Health_standards/tahc/2018/en_chapitre_avian_influenza_viruses.htm (accessed on 26 July 2022).
- Seekings, A.; Warren, C.J.; Thomas, S.S.; Mahmood, S.; James, J.; Byrne, A.M.P.; Watson, S.; Bianco, C.; Nunez, A.; Brown, I.H.; et al. Highly pathogenic avian influenza virus H5N6 (clade 2.3.4.4b) has a preferable host tropism for waterfowl reflected in its inefficient transmission to terrestrial poultry. Virology 2021, 559, 74–85. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2017, 35, 518–522. [Google Scholar] [CrossRef]
- Animalia. African Fish Eagle. Available online: https://animalia.bio/african-fish-eagle (accessed on 15 November 2021).
- Chittenden, H. Roberts Bird Guide: A Comprehensive Field Guide Over 950 Bird Species in Southern Africa; John Voelcker Bird Book Fund: Cape Town, South Africa, 2007. [Google Scholar]
- Animalia. Mourning Collared Dove. Animalia, Ukraine. Available online: https://animalia.bio/mourning-collared-dove (accessed on 15 November 2022).
- Flack, A.; Fiedler, W.; Blas, J.; Pokrovsky, I.; Kaatz, M.; Mitropolsky, M.; Aghababyan, K.; Fakriadis, I.; Makrigianni, E.; Jerzak, L.; et al. Costs of migratory decisions: A comparison across eight white stork populations. Sci. Adv. 2016, 2, e1500931. [Google Scholar] [CrossRef]
- Avibase. Avibase The World Bird Database: Okavango. Available online: https://avibase.bsc-eoc.org/checklist.jsp?region=NAok&list=howardmoore (accessed on 15 November 2022).




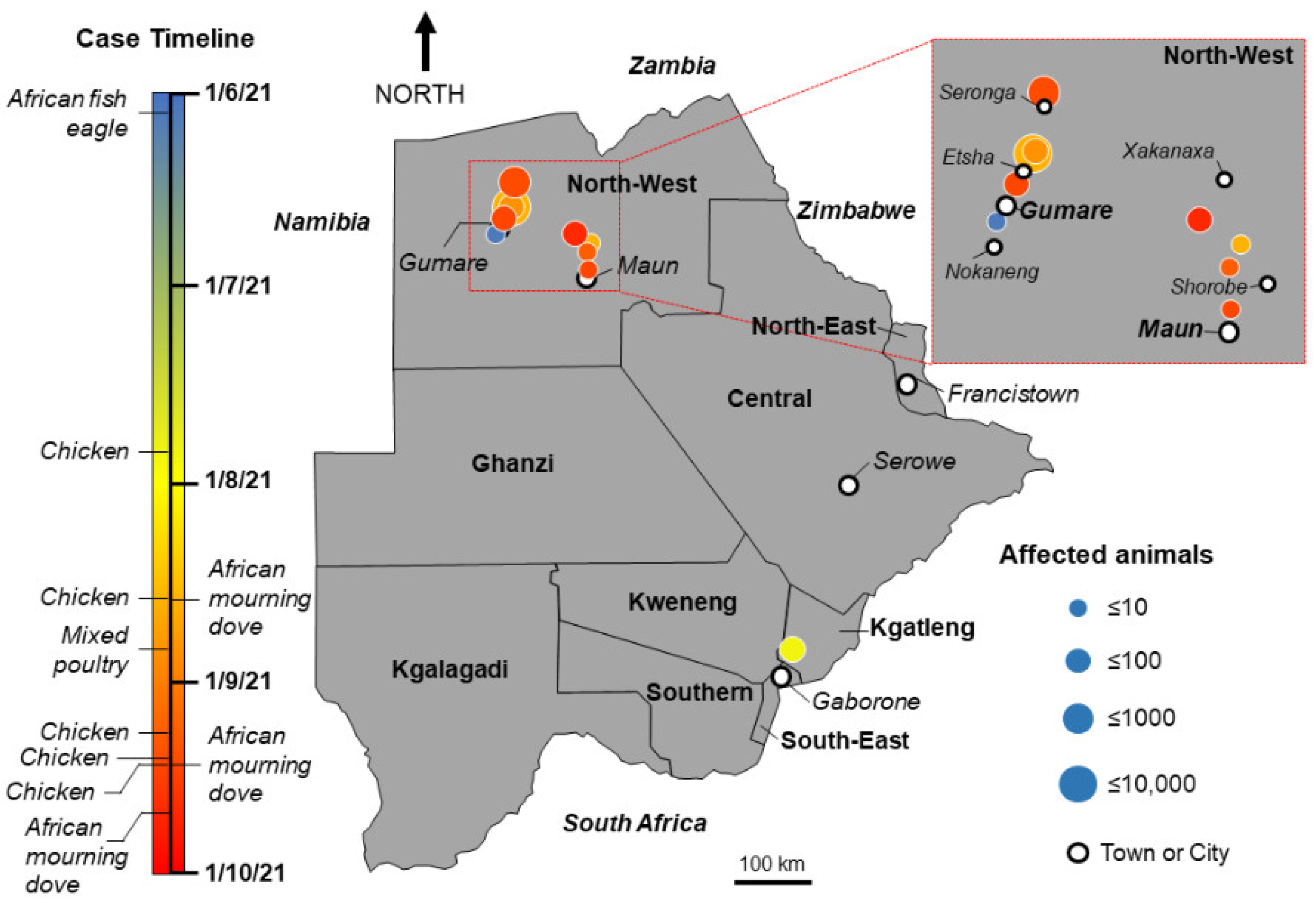{kind=link}
{kind=link}
{kind=link}
| Sample Location (and Collection Date) | Species | Sample ID | Sample Type | Accession Number |
|---|---|---|---|---|
| Habu, Okavango Delta 3 June 2021 | African fish eagle (Haliaeetus vocifer) | BNVL 2021/1338 | Pooled tissues (lung, trachea, liver, Spleen, intestine, proventriculus) | EPI_ISL_12045920 |
| Sedie, Maun East 18 August 2021 | African mourning dove (Streptopelia decipiens) | BNVL 2021/2097 | Pooled tissues (lung, trachea, liver, spleen, intestine, proventriculus) | EPI_ISL_12045918 |
| Etsha 13, Gumare Sub-District | Broiler backyard chickens 18 August 2021 | BNVL 2021/2163 | Trachea | EPI_ISL_12045319 |
| African mourning dove (Streptopelia decipiens) 13 September 2021 | BNVL 2021/2334 | Pooled tissues (Lung, trachea, liver, spleen, intestine, proventriculus) | EPI_ISL_12045919 | |
| Shorobe, 8 September 2021 | Tswana backyard chicken | BNVL 2021/2248 | Proventriculus | EPI_ISL_12045320 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Letsholo, S.L.; James, J.; Meyer, S.M.; Byrne, A.M.P.; Reid, S.M.; Settypalli, T.B.K.; Datta, S.; Oarabile, L.; Kemolatlhe, O.; Pebe, K.T.; et al. Emergence of High Pathogenicity Avian Influenza Virus H5N1 Clade 2.3.4.4b in Wild Birds and Poultry in Botswana. Viruses 2022, 14, 2601. https://doi.org/10.3390/v14122601
Letsholo SL, James J, Meyer SM, Byrne AMP, Reid SM, Settypalli TBK, Datta S, Oarabile L, Kemolatlhe O, Pebe KT, et al. Emergence of High Pathogenicity Avian Influenza Virus H5N1 Clade 2.3.4.4b in Wild Birds and Poultry in Botswana. Viruses. 2022; 14(12):2601. https://doi.org/10.3390/v14122601
Chicago/Turabian StyleLetsholo, Samantha L., Joe James, Stephanie M. Meyer, Alexander M. P. Byrne, Scott M. Reid, Tirumala B. K. Settypalli, Sneha Datta, Letlhogile Oarabile, Obakeng Kemolatlhe, Kgakgamatso T. Pebe, and et al. 2022. "Emergence of High Pathogenicity Avian Influenza Virus H5N1 Clade 2.3.4.4b in Wild Birds and Poultry in Botswana" Viruses 14, no. 12: 2601. https://doi.org/10.3390/v14122601