
Seroprevalence of Four Polyomaviruses Linked to Dermatological Diseases: New Findings and a Comprehensive Analysis
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples and Patients
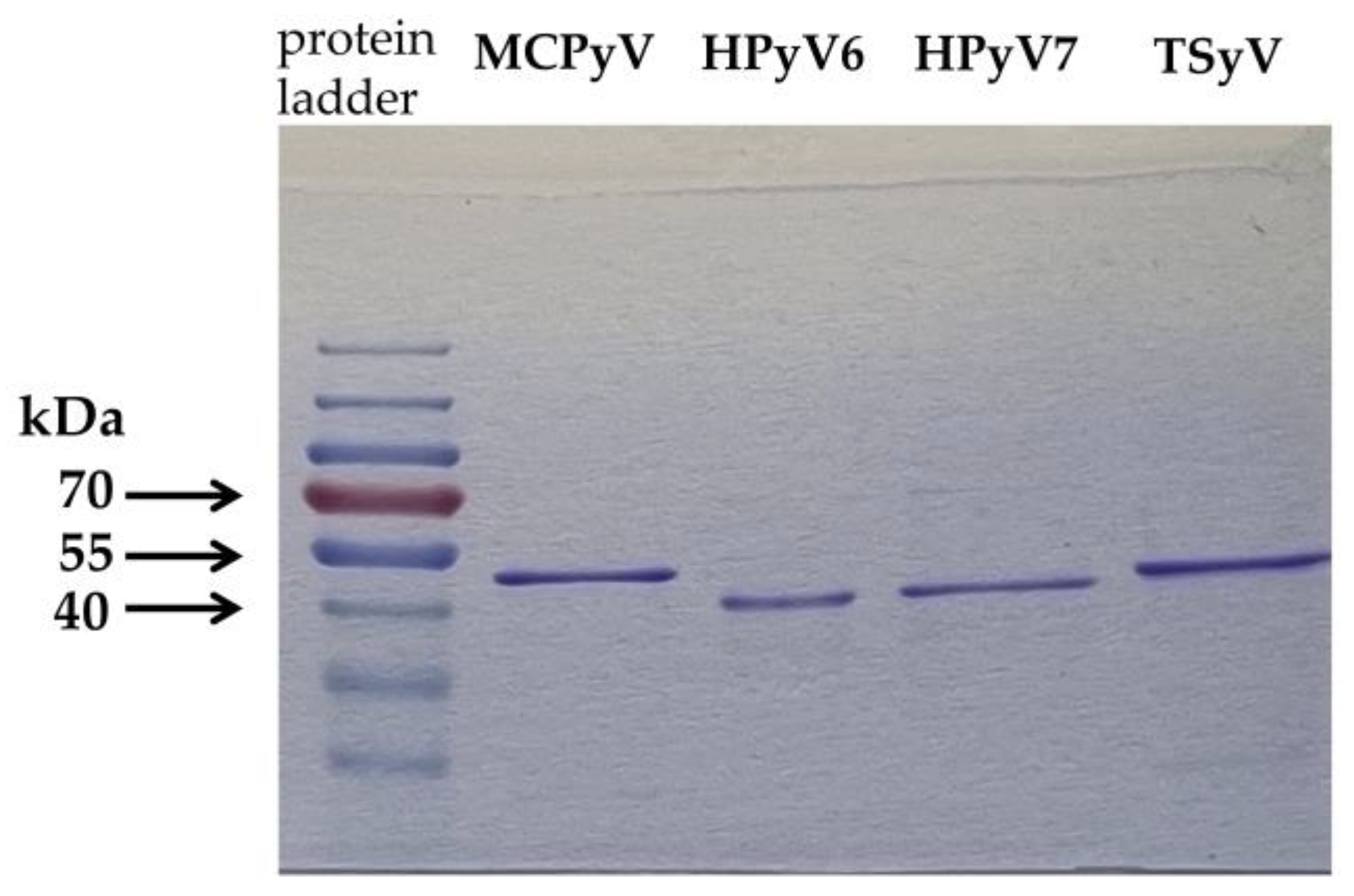
2.2. Antigen Production for ELISA
2.3. Detection of Anti-Polyomavirus Antibodies by ELISA
2.4. Statistical Analysis
3. Results and Discussion
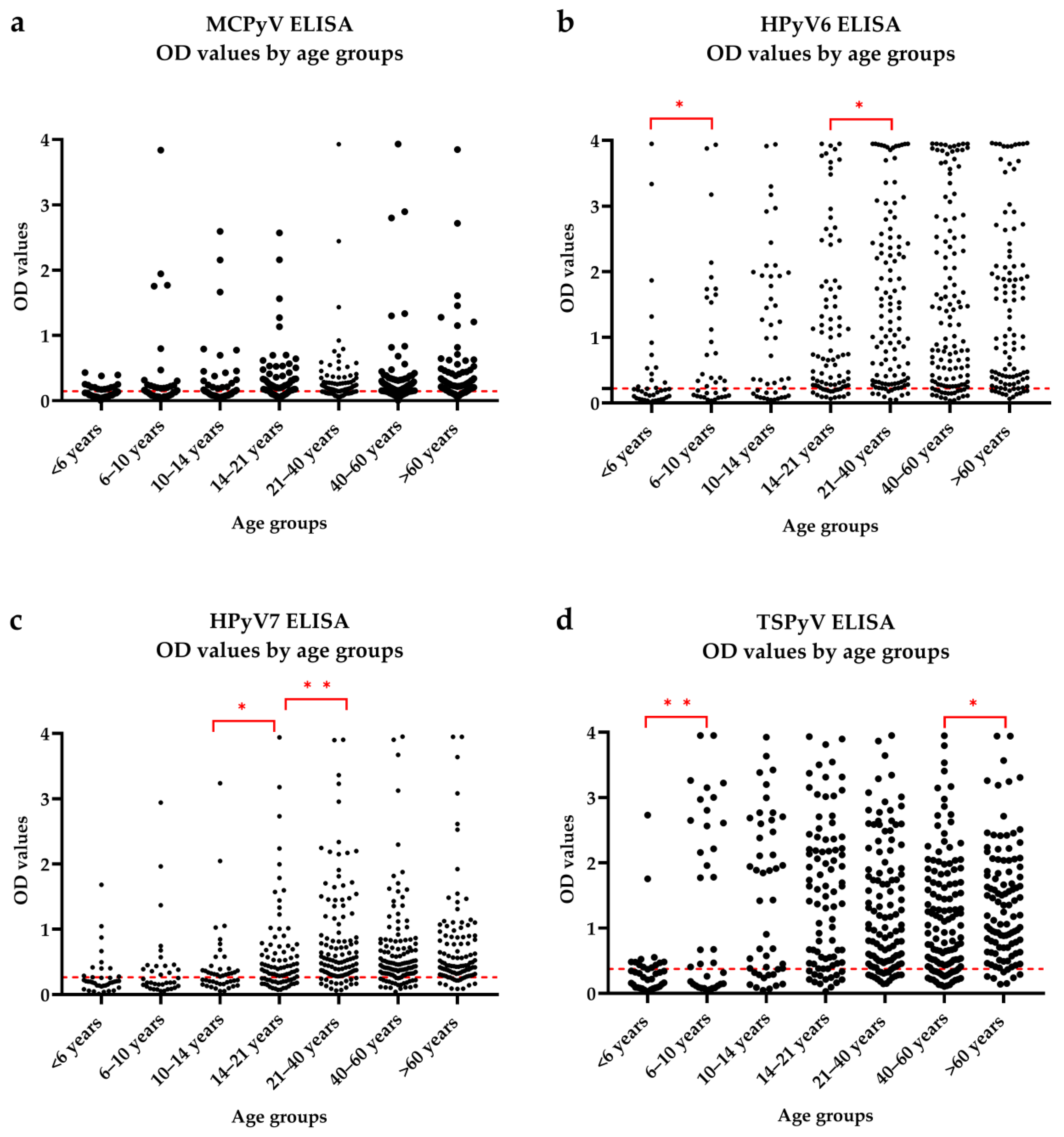
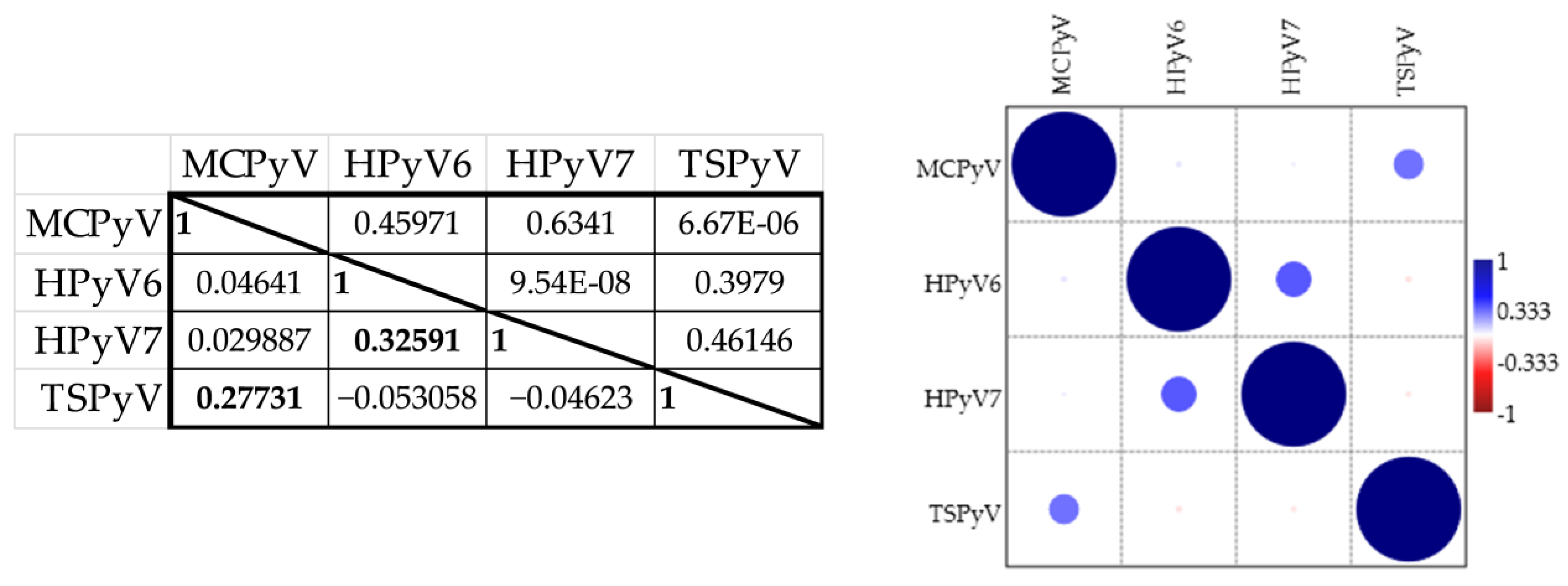
3.1. Seroresponses
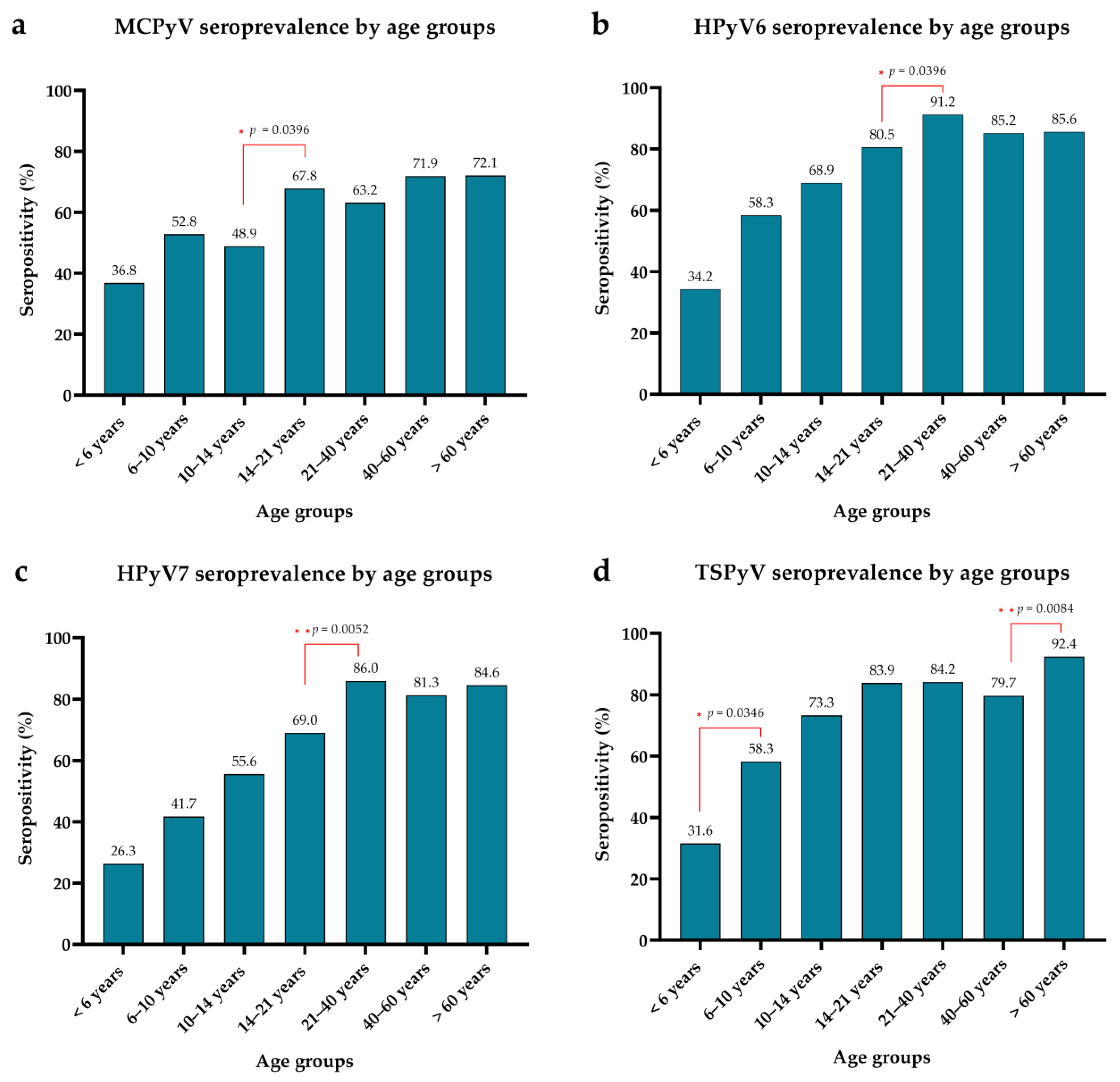
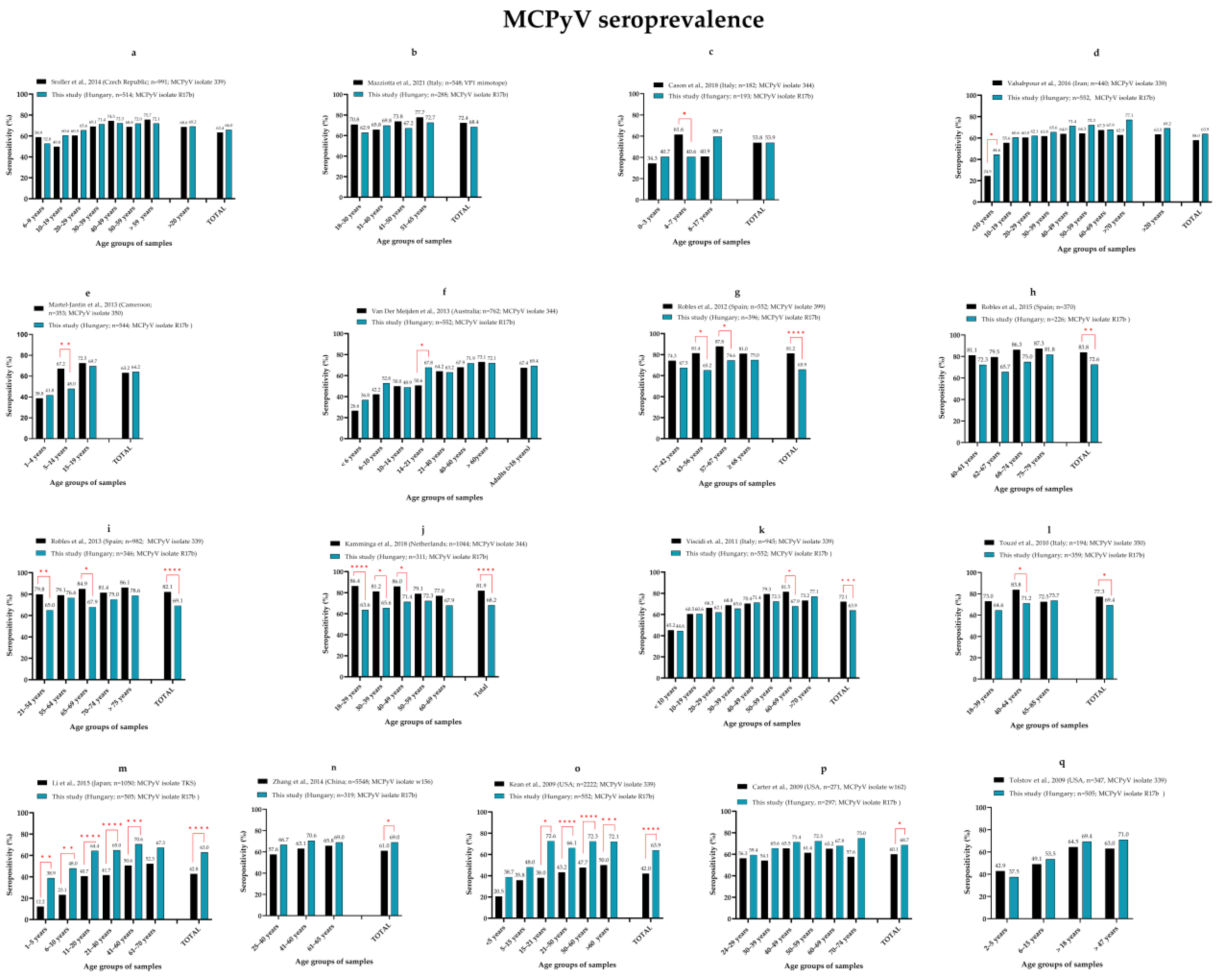
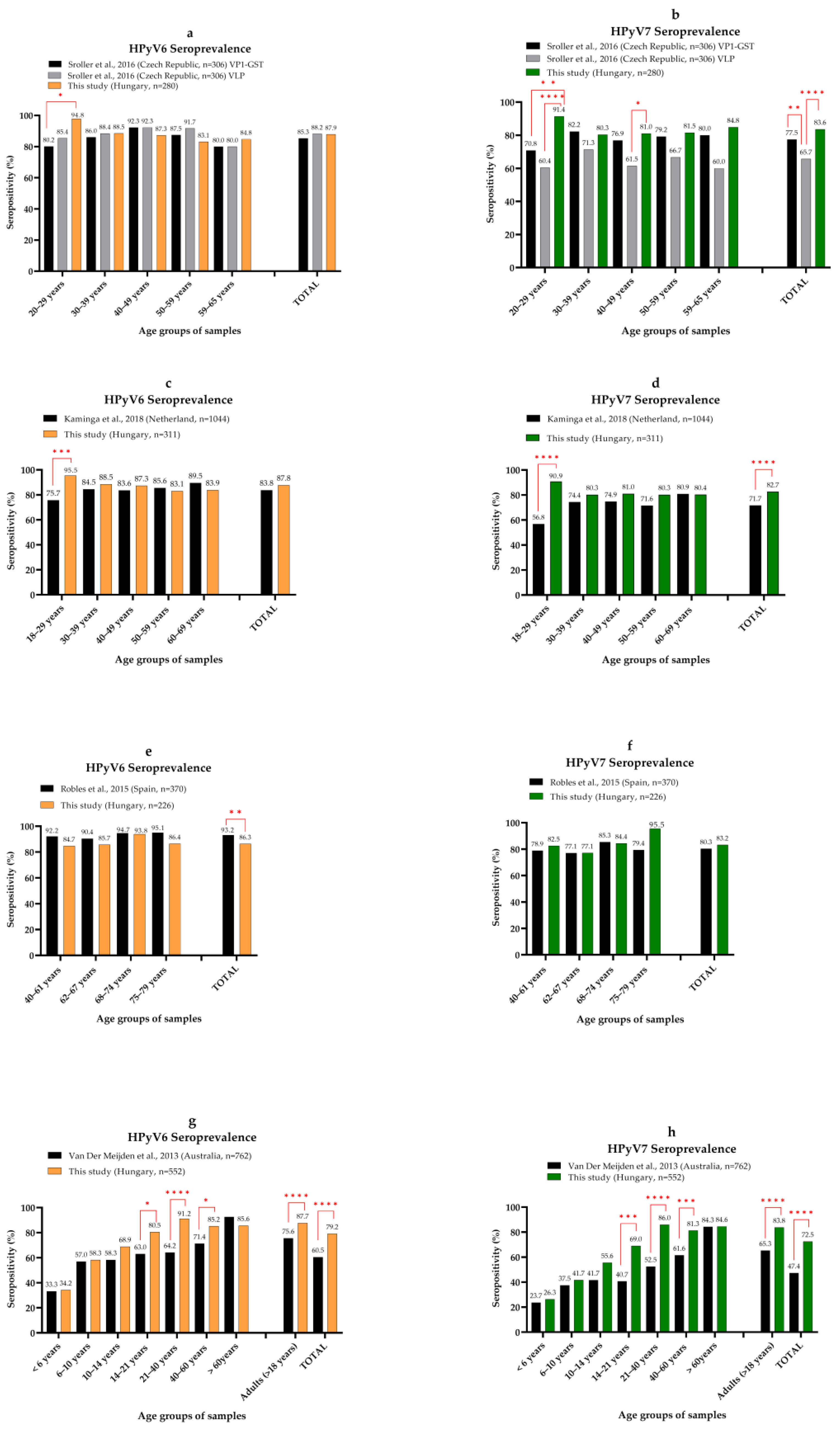
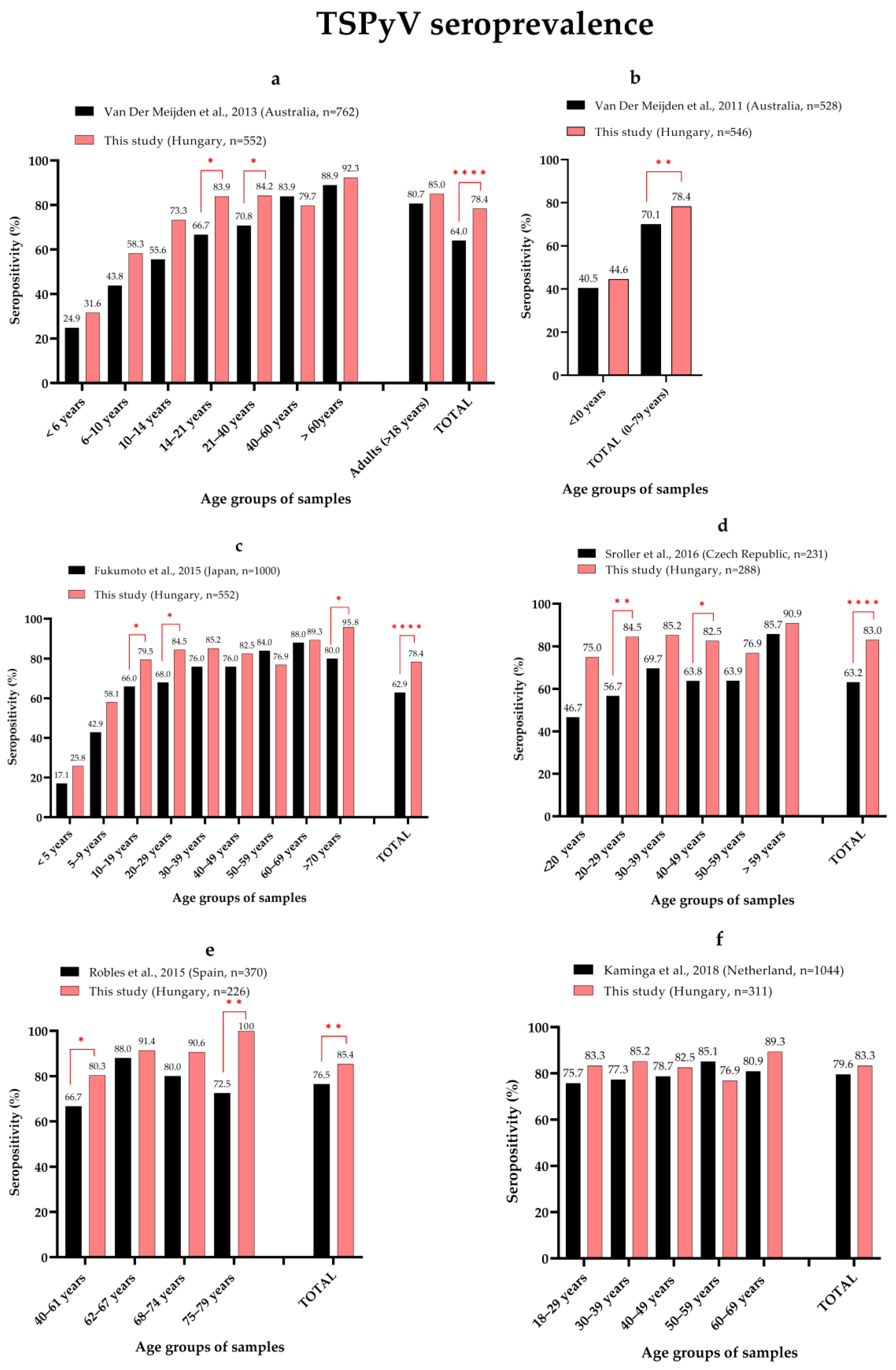
3.2. Seroprevalence of MCPyV, HPyV6, HPyV7 and TSPyV
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gardner, S.D.; Field, A.M.; Coleman, D.V.; Hulme, B. New human papovavirus (B.K.) isolated from urine after renal transplantation. Lancet 1971, 1, 1253–1257. [Google Scholar] [CrossRef]
- Padgett, B.L.; Walker, D.L.; ZuRhein, G.M.; Eckroade, R.J.; Dessel, B.H. Cultivation of papova-like virus from human brain with progressive multifocal leucoencephalopathy. Lancet 1971, 1, 1257–1260. [Google Scholar] [CrossRef]
- Moens, U.; Prezioso, C.; Pietropaolo, V. Genetic Diversity of the Noncoding Control Region of the Novel Human Polyomaviruses. Viruses 2020, 12, 1406. [Google Scholar] [CrossRef] [PubMed]
- Moens, U.; Calvignac-Spencer, S.; Lauber, C.; Ramqvist, T.; Feltkamp, M.C.W.; Daugherty, M.D.; Verschoor, E.J.; Ehlers, B.; Ictv Report, C. ICTV Virus Taxonomy Profile: Polyomaviridae. J. Gen. Virol. 2017, 98, 1159–1160. [Google Scholar] [CrossRef]
- Sheu, J.C.; Tran, J.; Rady, P.L.; Dao, H.; Tyring, S.K.; Nguyen, H.P. Polyomaviruses of the skin: Integrating molecular and clinical advances in an emerging class of viruses. Brit. J. Dermatol. 2019, 180, 1302–1311. [Google Scholar] [CrossRef]
- Nguyen, K.D.; Chamseddin, B.H.; Cockerell, C.J.; Wang, R.C. The Biology and Clinical Features of Cutaneous Polyomaviruses. J. Investig. Dermatol. 2019, 139, 285–292. [Google Scholar] [CrossRef] [Green Version]
- Feng, H.; Shuda, M.; Chang, Y.; Moore, P.S. Clonal integration of a polyomavirus in human Merkel cell carcinoma. Science 2008, 319, 1096–1100. [Google Scholar] [CrossRef] [Green Version]
- Paulson, K.G.; Park, S.Y.; Vandeven, N.A.; Lachance, K.; Thomas, H.; Chapuis, A.G.; Harms, K.L.; Thompson, J.A.; Bhatia, S.; Stang, A.; et al. Merkel cell carcinoma: Current US incidence and projected increases based on changing demographics. J. Am. Acad. Dermatol. 2018, 78, 457. [Google Scholar] [CrossRef]
- Becker, J.C.; Stang, A.; zur Hausen, A.; Fischer, N.; DeCaprio, J.A.; Tothill, R.W.; Lyngaa, R.; Hansen, U.K.; Ritter, C.; Nghiem, P.; et al. Epidemiology, biology and therapy of Merkel cell carcinoma: Conclusions from the EU project IMMOMEC. Cancer Immunol. Immun. 2018, 67, 341–351. [Google Scholar] [CrossRef]
- Fitzgerald, T.L.; Dennis, S.; Kachare, S.D.; Vohra, N.A.; Wong, J.H.; Zervos, E.E. Dramatic Increase in the Incidence and Mortality from Merkel Cell Carcinoma in the United States. Am. Surg. 2015, 81, 802–806. [Google Scholar] [CrossRef]
- Garbutcheon-Singh, K.B.; Curchin, D.J.; McCormack, C.J.; Smith, S.D. Trends in the incidence of Merkel cell carcinoma in Victoria, Australia, between 1986 and 2016. Australas. J. Dermatol. 2020, 61, E34–E38. [Google Scholar] [CrossRef]
- Tanda, E.T.; D’Amato, A.L.; Rossi, G.; Croce, E.; Boutros, A.; Cecchi, F.; Spagnolo, F.; Queirolo, P. Merkel Cell Carcinoma: An Immunotherapy Fairy-Tale? Front. Oncol. 2021, 11, 739006. [Google Scholar] [CrossRef]
- Wijaya, W.A.; Liu, Y.; Qing, Y.; Li, Z.Y. Prevalence of Merkel Cell Polyomavirus in Normal and Lesional Skin: A Systematic Review and Meta-Analysis. Front. Oncol. 2022, 12, 868781. [Google Scholar] [CrossRef]
- Spurgeon, M.E.; Lambert, P.F. Merkel cell polyomavirus: A newly discovered human virus with oncogenic potential. Virology 2013, 435, 118–130. [Google Scholar] [CrossRef] [Green Version]
- Mazzoni, E.; Rotondo, J.C.; Marracino, L.; Selvatici, R.; Bononi, I.; Torreggiani, E.; Touze, A.; Martini, F.; Tognon, M.G. Detection of Merkel Cell Polyomavirus DNA in Serum Samples of Healthy Blood Donors. Front. Oncol. 2017, 7, 294. [Google Scholar] [CrossRef] [Green Version]
- Mazziotta, C.; Lanzillotti, C.; Torreggiani, E.; Oton-Gonzalez, L.; Iaquinta, M.R.; Mazzoni, E.; Gaboriaud, P.; Touze, A.; Silvagni, E.; Govoni, M.; et al. Serum Antibodies Against the Oncogenic Merkel Cell Polyomavirus Detected by an Innovative Immunological Assay With Mimotopes in Healthy Subjects. Front. Immunol. 2021, 12, 676627. [Google Scholar] [CrossRef]
- Schowalter, R.M.; Pastrana, D.V.; Pumphrey, K.A.; Moyer, A.L.; Buck, C.B. Merkel cell polyomavirus and two previously unknown polyomaviruses are chronically shed from human skin. Cell Host Microbe 2010, 7, 509–515. [Google Scholar]
- Ho, J.H.; Jedrych, J.J.; Feng, H.C.; Natalie, A.A.; Grandinetti, L.; Mirvish, E.; Crespo, M.M.; Yadav, D.; Fasanella, K.E.; Proksell, S.; et al. Human Polyomavirus 7-Associated Pruritic Rash and Viremia in Transplant Recipients. J. Infect. Dis. 2015, 211, 1560–1565. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, K.D.; Lee, E.E.; Yue, Y.B.; Stork, J.; Pock, L.; North, J.P.; Vandergriff, T.; Cockerell, C.; Hosler, G.A.; Pastrana, D.V.; et al. Human polyomavirus 6 and 7 are associated with pruritic and dyskeratotic dermatoses. J. Am. Acad. Dermatol. 2017, 76, 932. [Google Scholar] [CrossRef] [Green Version]
- Hashida, Y.; Higuchi, T.; Matsuzaki, S.; Nakajima, K.; Sano, S.; Daibata, M. Prevalence and Genetic Variability of Human Polyomaviruses 6 and 7 in Healthy Skin Among Asymptomatic Individuals. J. Infect. Dis. 2018, 217, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Bopp, L.; Wieland, U.; Hellmich, M.; Kreuter, A.; Pfister, H.; Silling, S. Natural History of Cutaneous Human Polyomavirus Infection in Healthy Individuals. Front. Microbiol. 2021, 12, 740947. [Google Scholar] [CrossRef] [PubMed]
- Foulongne, V.; Sauvage, V.; Hebert, C.; Dereure, O.; Cheval, J.; Gouilh, M.A.; Pariente, K.; Segondy, M.; Burguiere, A.; Manuguerra, J.C.; et al. Human Skin Microbiota: High Diversity of DNA Viruses Identified on the Human Skin by High Throughput Sequencing. PLoS ONE 2012, 7, e38499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wieland, U.; Silling, S.; Hellmich, M.; Potthoff, A.; Pfister, H.; Kreuter, A. Human polyomaviruses 6, 7, 9, 10 and Trichodysplasia spinulosa-associated polyomavirus in HIV-infected men. J. Gen. Virol. 2014, 95, 928–932. [Google Scholar] [CrossRef] [PubMed]
- Wylie, K.M.; Mihindukulasuriya, K.A.; Zhou, Y.J.; Sodergren, E.; Storch, G.A.; Weinstock, G.M. Metagenomic analysis of double-stranded DNA viruses in healthy adults. BMC Biol. 2014, 12, 71. [Google Scholar] [CrossRef]
- Fava, P.; Merlino, C.; Novelli, M.; Ponti, R.; Galliano, I.; Montanari, P.; Tovo, P.A.; Fierro, M.T.; Bergallo, M. HPyV6, HPyV7 and TSPyV DNA sequences detection in skin disease patients and healthy subjects. J. Eur. Acad. Dermatol. 2016, 30, 624–627. [Google Scholar] [CrossRef]
- Klufah, F.; Mobaraki, G.; Liu, D.; Alharbi, R.A.; Kurz, A.K.; Speel, E.J.M.; Winnepenninckx, V.; zur Hausen, A. Emerging role of human polyomaviruses 6 and 7 in human cancers. Infect. Agents Cancer 2021, 16, 35. [Google Scholar] [CrossRef]
- Van der Meijden, E.; Janssens, R.W.; Lauber, C.; Bouwes Bavinck, J.N.; Gorbalenya, A.E.; Feltkamp, M.C. Discovery of a new human polyomavirus associated with trichodysplasia spinulosa in an immunocompromized patient. PLoS Pathog. 2010, 6, e1001024. [Google Scholar] [CrossRef] [Green Version]
- Curman, P.; Nasman, A.; Brauner, H. Trichodysplasia spinulosa: A comprehensive review of the disease and its treatment. J. Eur. Acad. Dermatol. 2021, 35, 1067–1076. [Google Scholar] [CrossRef]
- Kazem, S.; van der Meijden, E.; Kooijman, S.; Rosenberg, A.S.; Hughey, L.C.; Browning, J.C.; Sadler, G.; Busam, K.; Pope, E.; Benoit, T.; et al. Trichodysplasia spinulosa is characterized by active polyomavirus infection. J. Clin. Virol. 2012, 53, 225–230. [Google Scholar] [CrossRef]
- Van der Meijden, E.; Horvath, B.; Nijland, M.; de Vries, K.; Raez, E.; Diercks, G.F.; de Weerd, A.E.; Clahsen-van Groningen, M.C.; van der Blij-de Brouwer, C.S.; van der Zon, A.J.; et al. Primary Polyomavirus Infection, Not Reactivation, as the Cause of Trichodysplasia Spinulosa in Immunocompromised Patients. J. Infect. Dis. 2017, 215, 1080–1084. [Google Scholar] [CrossRef] [Green Version]
- Martel-Jantin, C.; Filippone, C.; Tortevoye, P.; Afonso, P.V.; Betsem, E.; Descorps-Declere, S.; Nicol, J.T.J.; Touze, A.; Coursaget, P.; Crouzat, M.; et al. Molecular Epidemiology of Merkel Cell Polyomavirus: Evidence for Geographically Related Variant Genotypes. J. Clin. Microbiol. 2014, 52, 1687–1690. [Google Scholar] [CrossRef]
- Hashida, Y.; Higuchi, T.; Matsui, K.; Shibata, Y.; Nakajima, K.; Sano, S.; Daibata, M. Genetic Variability of the Noncoding Control Region of Cutaneous Merkel Cell Polyomavirus: Identification of Geographically Related Genotypes. J. Infect. Dis. 2018, 217, 1601–1611. [Google Scholar] [CrossRef] [Green Version]
- Katona, M.; Jeles, K.; Kovacs, R.; Csoma, E. KI and WU Polyomaviruses: Seroprevalence Study and DNA Prevalence in SARS-CoV-2 RNA Positive and Negative Respiratory Samples. Microorganisms 2022, 10, 752. [Google Scholar] [CrossRef]
- Nicol, J.T.; Robinot, R.; Carpentier, A.; Carandina, G.; Mazzoni, E.; Tognon, M.; Touze, A.; Coursaget, P. Age-specific seroprevalence of Merkel cell polyomavirus, Human polyomaviruses 6, 7 and 9 and Trichodysplasia Spinulosa-associated polyomavirus. Clin. Vaccine Immunol. 2013, 20, 363–368. [Google Scholar] [CrossRef]
- Van der Meijden, E.; Bialasiewicz, S.; Rockett, R.J.; Tozer, S.J.; Sloots, T.P.; Feltkamp, M.C. Different serologic behavior of MCPyV, TSPyV, HPyV6, HPyV7 and HPyV9 polyomaviruses found on the skin. PLoS ONE 2013, 8, e81078. [Google Scholar] [CrossRef] [Green Version]
- Sroller, V.; Hamsikova, E.; Ludvikova, V.; Musil, J.; Nemeckova, S.; Salakova, M. Seroprevalence rates of HPyV6, HPyV7, TSPyV, HPyV9, MWPyV and KIPyV polyomaviruses among the healthy blood donors. J. Med. Virol. 2016, 88, 1254–1261. [Google Scholar] [CrossRef]
- Moens, U.; Van Ghelue, M.; Song, X.B.; Ehlers, B. Serological cross-reactivity between human polyomaviruses. Rev. Med. Virol. 2013, 23, 250–264. [Google Scholar] [CrossRef]
- Kamminga, S.; van der Meijden, E.; Feltkamp, M.C.W.; Zaaijer, H.L. Seroprevalence of fourteen human polyomaviruses determined in blood donors. PLoS ONE 2018, 13, e0206273. [Google Scholar] [CrossRef] [Green Version]
- Tolstov, Y.L.; Pastrana, D.V.; Feng, H.C.; Becker, J.C.; Jenkins, F.J.; Moschos, S.; Chang, Y.; Buck, C.B.; Moore, P.S. Human Merkel cell polyomavirus infection II. MCV is a common human infection that can be detected by conformational capsid epitope immunoassays. Int. J. Cancer 2009, 125, 1250–1256. [Google Scholar] [CrossRef] [Green Version]
- Robles, C.; Casabonne, D.; Benavente, Y.; Costas, L.; Gonzalez-Barca, E.; Aymerich, M.; Campo, E.; Tardon, A.; Jimenez-Moleon, J.J.; Castano-Vinyals, G.; et al. Seroreactivity against Merkel cell polyomavirus and other polyomaviruses in chronic lymphocytic leukaemia, the MCC-Spain study. J. Gen. Virol. 2015, 96, 2286–2292. [Google Scholar] [CrossRef] [Green Version]
- Sroller, V.; Hamsikova, E.; Ludvikova, V.; Vochozkova, P.; Kojzarova, M.; Fraiberk, M.; Salakova, M.; Moravkova, A.; Forstova, J.; Nemeckova, S. Seroprevalence Rates of BKV, JCV, and MCPyV Polyomaviruses in the General Czech Republic Population. J. Med. Virol. 2014, 86, 1560–1568. [Google Scholar] [CrossRef]
- Cason, C.; Monasta, L.; Zanotta, N.; Campisciano, G.; Maestri, I.; Tommasino, M.; Pawlita, M.; Villani, S.; Comar, M.; Delbue, S. Antibody response to polyomavirus primary infection: High seroprevalence of Merkel cell polyomavirus and lymphoid tissue involvement. J. Neurovirol. 2018, 24, 314–322. [Google Scholar] [CrossRef]
- Vahabpour, R.; Aghasadeghi, M.R.; Salehi-Vaziri, M.; Mohajel, N.; Keyvani, H.; Nasimi, M.; Esghaei, M.; Monavari, S.H. Prevalence of Merkel Cell Polyomavirus in Tehran: An Age-Specific Serological Study. Iran. Red Crescent Med. J. 2016, 18, e26097. [Google Scholar] [CrossRef] [Green Version]
- Martel-Jantin, C.; Pedergnana, V.; Nicol, J.T.J.; Leblond, V.; Tregouet, D.A.; Tortevoye, P.; Plancoulaine, S.; Coursaget, P.; Touze, A.; Abel, L.; et al. Merkel cell polyomavirus infection occurs during early childhood and is transmitted between siblings. J. Clin. Virol. 2013, 58, 288–291. [Google Scholar] [CrossRef] [Green Version]
- Robles, C.; Poloczek, A.; Casabonne, D.; Gonzalez-Barca, E.; Bosch, R.; Benavente, Y.; Viscidi, R.P.; de Sanjos, S. Antibody Response to Merkel Cell Polyomavirus Associated with Incident Lymphoma in the Epilymph Case-Control Study in Spain. Cancer Epidem. Biomar. 2012, 21, 1592–1598. [Google Scholar] [CrossRef] [Green Version]
- Robles, C.; Viscidi, R.; Malats, N.; Silverman, D.T.; Tardon, A.; Garcia-Closas, R.; Serra, C.; Carrato, A.; Herranz, J.; Lloreta, J.; et al. Bladder cancer and seroreactivity to BK, JC and Merkel cell polyomaviruses: The Spanish bladder cancer study. Int. J. Cancer 2013, 133, 597–603. [Google Scholar] [CrossRef] [Green Version]
- Viscidi, R.P.; Rollison, D.E.; Sondak, V.K.; Silver, B.; Messina, J.L.; Giuliano, A.R.; Fulp, W.; Ajidahun, A.; Rivanera, D. Age-Specific Seroprevalence of Merkel Cell Polyomavirus, BK Virus, and JC Virus. Clin. Vaccine Immunol. 2011, 18, 1737–1743. [Google Scholar] [CrossRef] [Green Version]
- Touze, A.; Gaitan, J.; Arnold, F.; Cazal, R.; Fleury, M.J.; Combelas, N.; Sizaret, P.Y.; Guyetant, S.; Maruani, A.; Baay, M.; et al. Generation of Merkel Cell Polyomavirus (MCV)-Like Particles and Their Application to Detection of MCV Antibodies. J. Clin. Microbiol. 2010, 48, 1767–1770. [Google Scholar] [CrossRef] [Green Version]
- Li, T.C.; Iwasaki, K.; Katano, H.; Kataoka, M.; Nagata, N.; Kobayashi, K.; Mizutani, T.; Takeda, N.; Wakita, T.; Suzuki, T. Characterization of Self-Assembled Virus-Like Particles of Merkel Cell Polyomavirus. PLoS ONE 2015, 10, e0115646. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.Y.; Liu, F.F.; He, Z.H.; Deng, Q.J.; Pan, Y.Q.; Liu, Y.; Zhang, C.T.; Ning, T.; Guo, C.H.; Liang, Y.M.; et al. Seroprevalence of Merkel Cell Polyomavirus in the General Rural Population of Anyang, China. PLoS ONE 2014, 9, e106430. [Google Scholar] [CrossRef]
- Kean, J.M.; Rao, S.; Wang, M.; Garcea, R.L. Seroepidemiology of Human Polyomaviruses. PLoS Pathog. 2009, 5, e1000363. [Google Scholar] [CrossRef] [PubMed]
- Carter, J.J.; Paulson, K.G.; Wipf, G.C.; Miranda, D.; Madeleine, M.M.; Johnson, L.G.; Lemos, B.D.; Lee, S.; Warcola, A.H.; Iyer, J.G.; et al. Association of Merkel Cell Polyomavirus-Specific Antibodies With Merkel Cell Carcinoma. JNCI J. Natl. Cancer I. 2009, 101, 1510–1522. [Google Scholar] [CrossRef] [PubMed]
- Van der Meijden, E.; Kazem, S.; Burgers, M.M.; Janssens, R.; Bavinck, J.N.B.; de Melker, H.; Feltkamp, M.C.W. Seroprevalence of Trichodysplasia Spinulosa-associated Polyomavirus. Emerg. Infect. Dis. 2011, 17, 1355–1363. [Google Scholar] [CrossRef]
- Fukumoto, H.; Li, T.C.; Kataoka, M.; Hasegawa, H.; Wakita, T.; Saeki, H.; Suzuki, T.; Katano, H. Seroprevalence of trichodysplasia spinulosa-associated polyomavirus in Japan. J. Clin. Virol. 2015, 65, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.F.; Nakashima, K.; Ito, M.; Zhang, X.L.; Sakai, S.; Feng, C.H.; Sun, H.B.; Chen, H.Y.; Li, T.C.; Suzuki, T. Prevalence and viral loads of polyomaviruses BKPyV, JCPyV, MCPyV, TSPyV and NJPyV and hepatitis viruses HBV, HCV and HEV in HIV-infected patients in China. Sci. Rep. 2020, 10, 17066. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, M.; Aaltonen, L.M.; Hedman, L.; Chen, T.T.; Soderlund-Venermo, M.; Hedman, K. Detection of TS polyomavirus DNA in tonsillar tissues of children and adults: Evidence for site of viral latency. J. Clin. Virol. 2014, 59, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Gossai, A.; Waterboer, T.; Nelson, H.H.; Michel, A.; Willhauck-Fleckenstein, M.; Farzan, S.F.; Hoen, A.G.; Christensen, B.C.; Kelsey, K.T.; Marsit, C.J.; et al. Seroepidemiology of Human Polyomaviruses in a US Population. Am. J. Epidemiol. 2016, 183, 61–69. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age Groups | Number of Samples | Age in Years, Min–Max (Median) | Female/Male |
|---|---|---|---|
| <6 years | 38 | 0.8–5.9 (2.8) | 15/23 |
| 6–10 years | 36 | 6.1–9.9 (7.6) | 19/17 |
| 10–14 years | 45 | 10.1–13.9 (11.7) | 22/23 |
| 14–21 years | 87 | 14–20 (16) | 47/40 |
| 21–40 years | 114 | 21–39.5 (30.8) | 57/57 |
| 40–60 years | 128 | 40–59.5 (50) | 66/62 |
| >60 years | 104 | 60–85 (69) | 57/47 |
| Total | 552 | 0.8–85 (33) | 283/269 |
| Adults | 359 | 18–85 (47.3) | 187/172 |
| Children | 193 | 0.8–17.9 (11.7) | 96/97 |
| Number of Seropositive Samples (Female/Male) | ||||
|---|---|---|---|---|
| Age Groups | MCPyV | HPyV6 | HPyV7 | TSPyV |
| <6 years | 14 (6/8) | 13 (6/7) | 10 (5/5) | 12 (4/8) |
| 6–10 years | 19 (12/7) | 21 (15/6) * | 15 (10/5) | 21 (8/13) * |
| 10–14 years | 22 (14/8) | 31 (17/14) | 25 (15/10) | 33 (15/18) |
| 14–21 years | 59 (31/28) | 70 (37/33) | 60 (36/24) | 73 (40/33) |
| 21–40 years | 72 (36/36) | 104 (51/53) | 98 (49/49) | 96 (49/47) |
| 40–60 years | 92 (52/40) | 109 (58/51) | 104 (54/50) | 102 (52/50) |
| >60 years | 75 (44/31) | 89 (50/39) | 88 (49/39) | 96 (55/41) |
| Total | 353 (195/158) * | 437 (234/203) * | 400 (218/182) * | 433 (223/210) |
| Adults | 249 (137/112) | 315 (166/149) | 301 (159/142) | 305 (162/143) |
| Children | 104 (58/46) | 122 (68/54) * | 99 (59/40) ** | 128 (61/67) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeles, K.; Katona, M.; Csoma, E. Seroprevalence of Four Polyomaviruses Linked to Dermatological Diseases: New Findings and a Comprehensive Analysis. Viruses 2022, 14, 2282. https://doi.org/10.3390/v14102282
Jeles K, Katona M, Csoma E. Seroprevalence of Four Polyomaviruses Linked to Dermatological Diseases: New Findings and a Comprehensive Analysis. Viruses. 2022; 14(10):2282. https://doi.org/10.3390/v14102282
Chicago/Turabian StyleJeles, Krisztina, Melinda Katona, and Eszter Csoma. 2022. "Seroprevalence of Four Polyomaviruses Linked to Dermatological Diseases: New Findings and a Comprehensive Analysis" Viruses 14, no. 10: 2282. https://doi.org/10.3390/v14102282