
Molecular Characterization of Novel Mycoviruses in Seven Umbelopsis Strains
 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Cultivation
2.2. Purification of Nucleic Acids
2.3. cDNA Synthesis and Sequencing of the dsRNA Molecules
2.4. Generation of RNA Sequencing Library, NGS Sequencing and Bioinformatic Analysis
2.5. Sequence and Phylogenetic Analysis
2.6. Hybridization Analysis
2.7. Examination of Virus Particles
3. Results
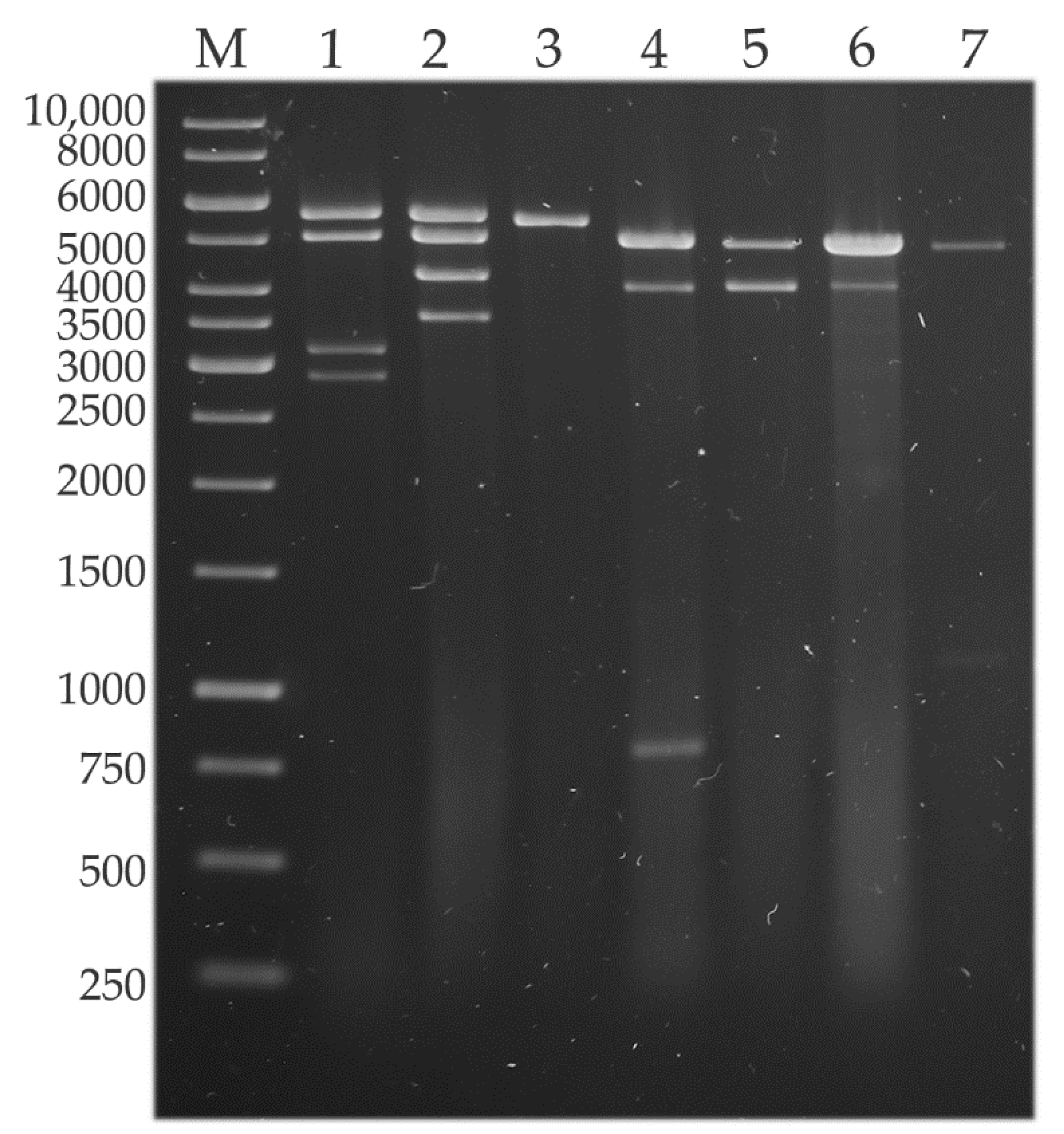
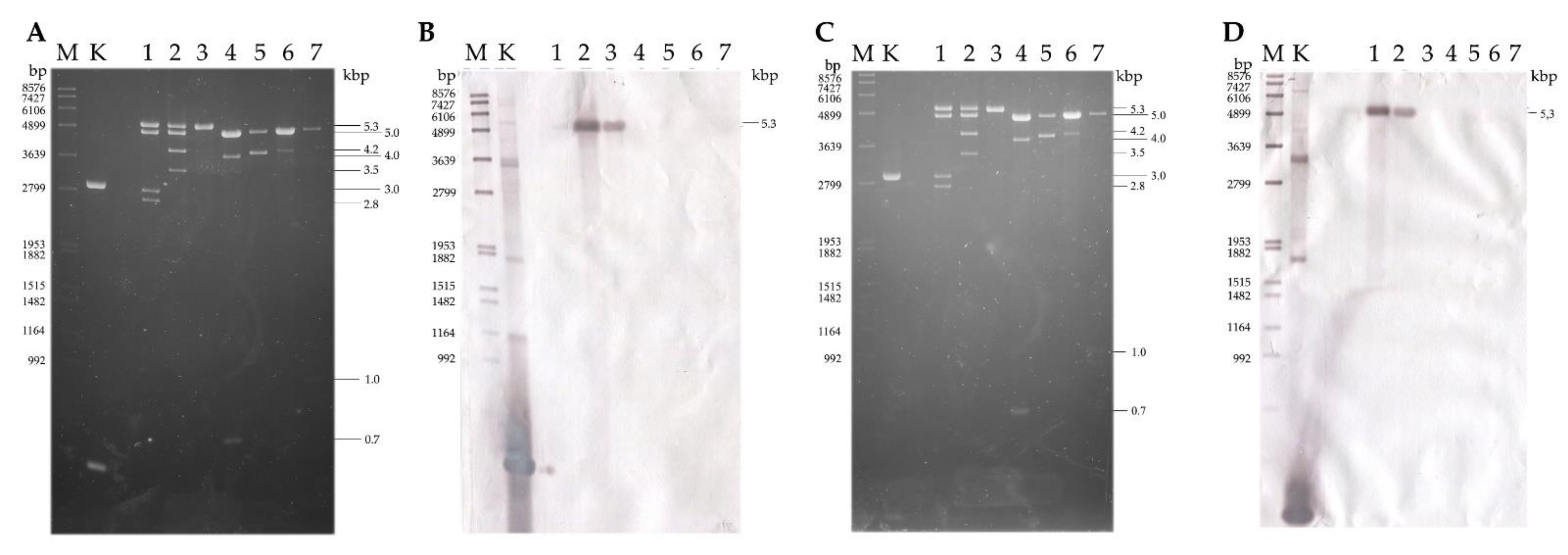
3.1. dsRNA Harboring in the Genus Umbelopsis
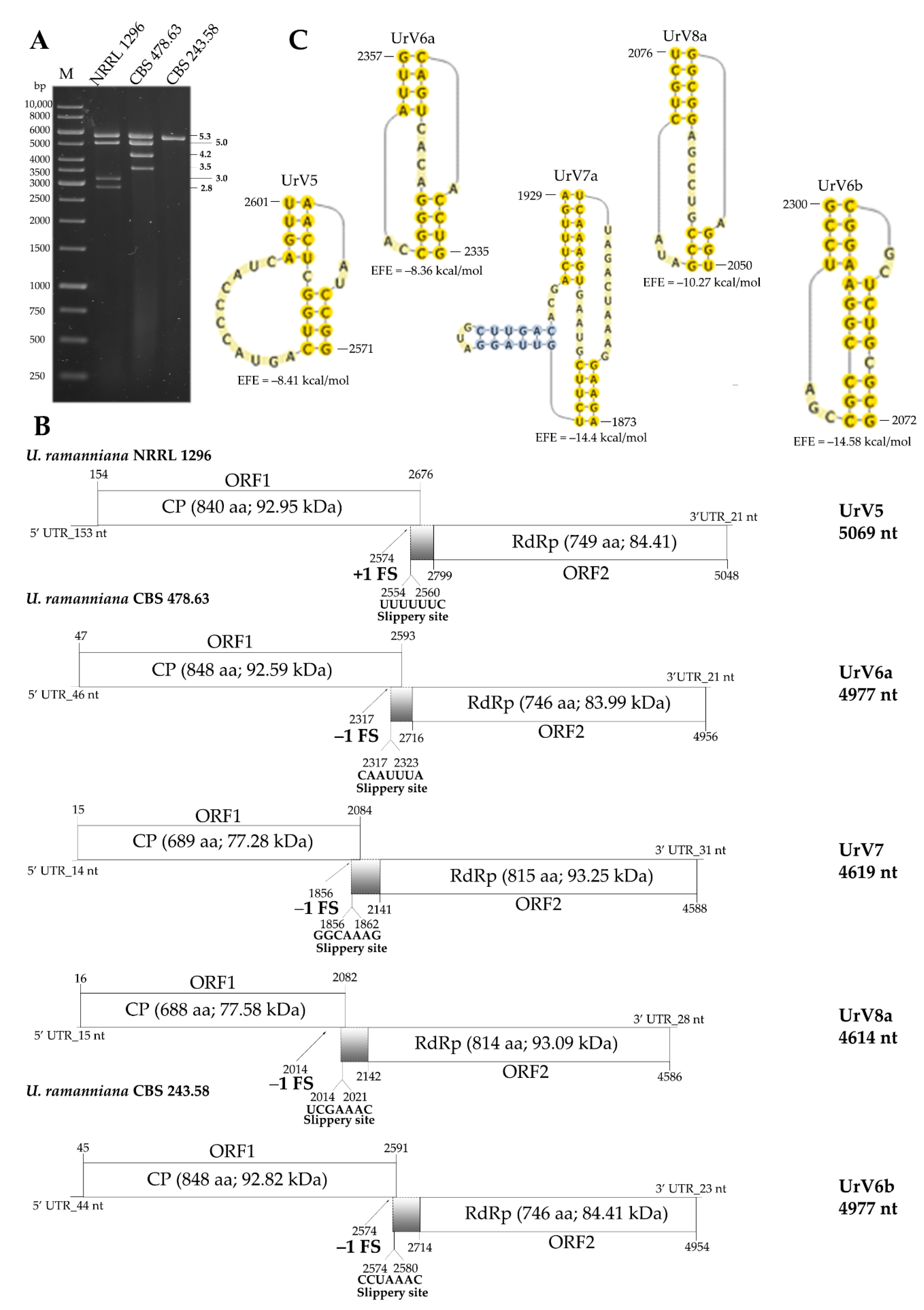
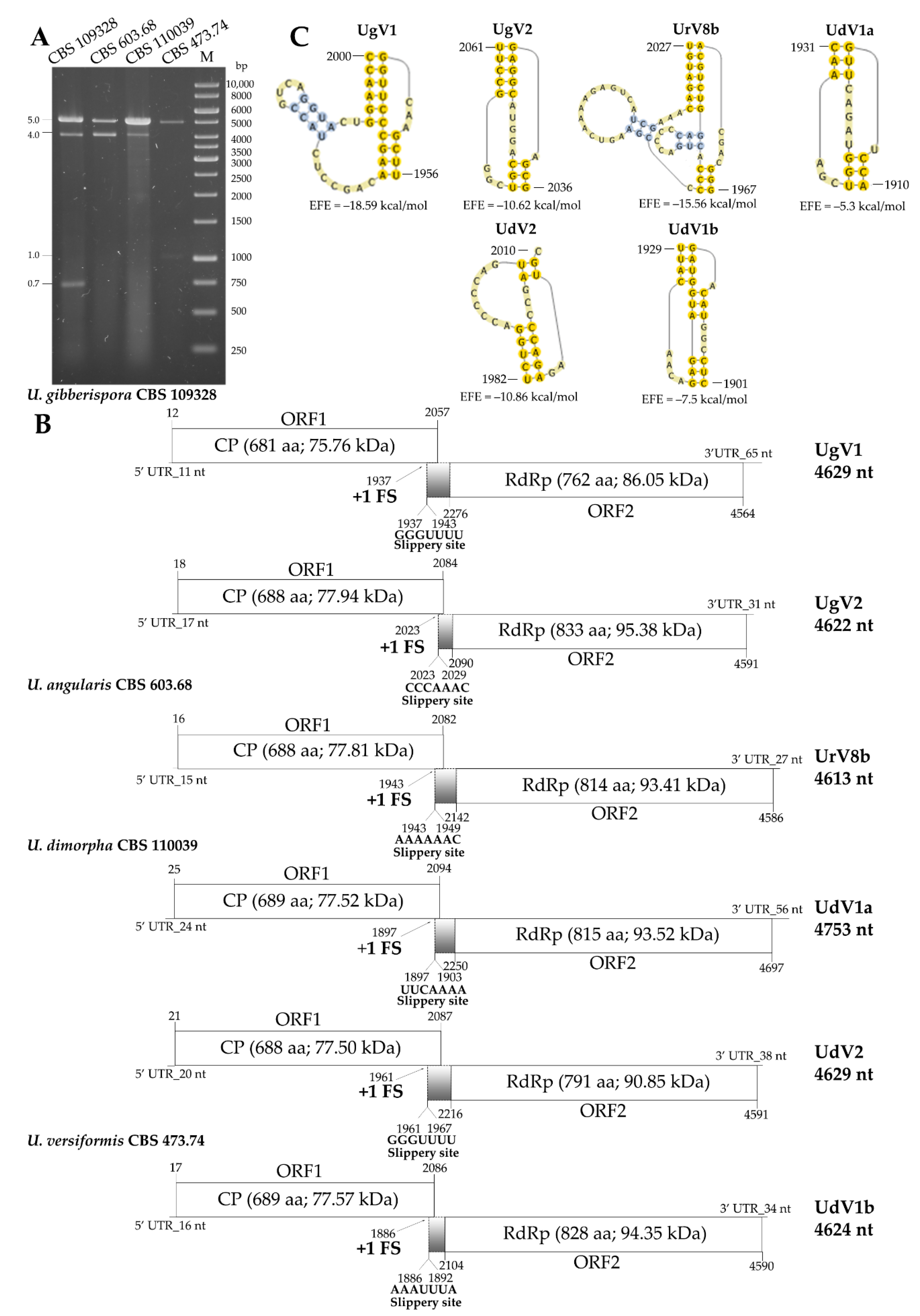
3.2. Whole Virus Genomes Identified in the Umbelopsis Isolates
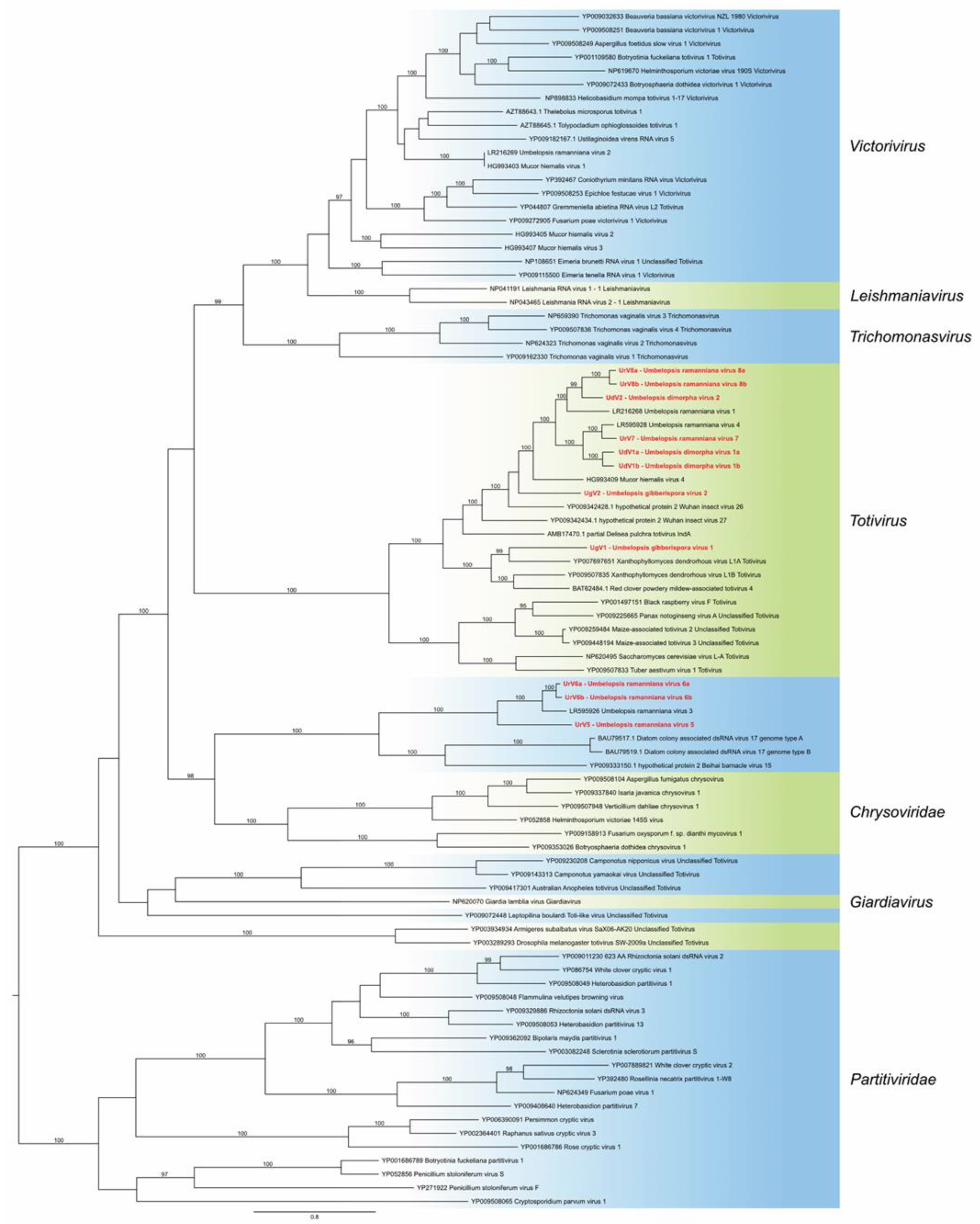
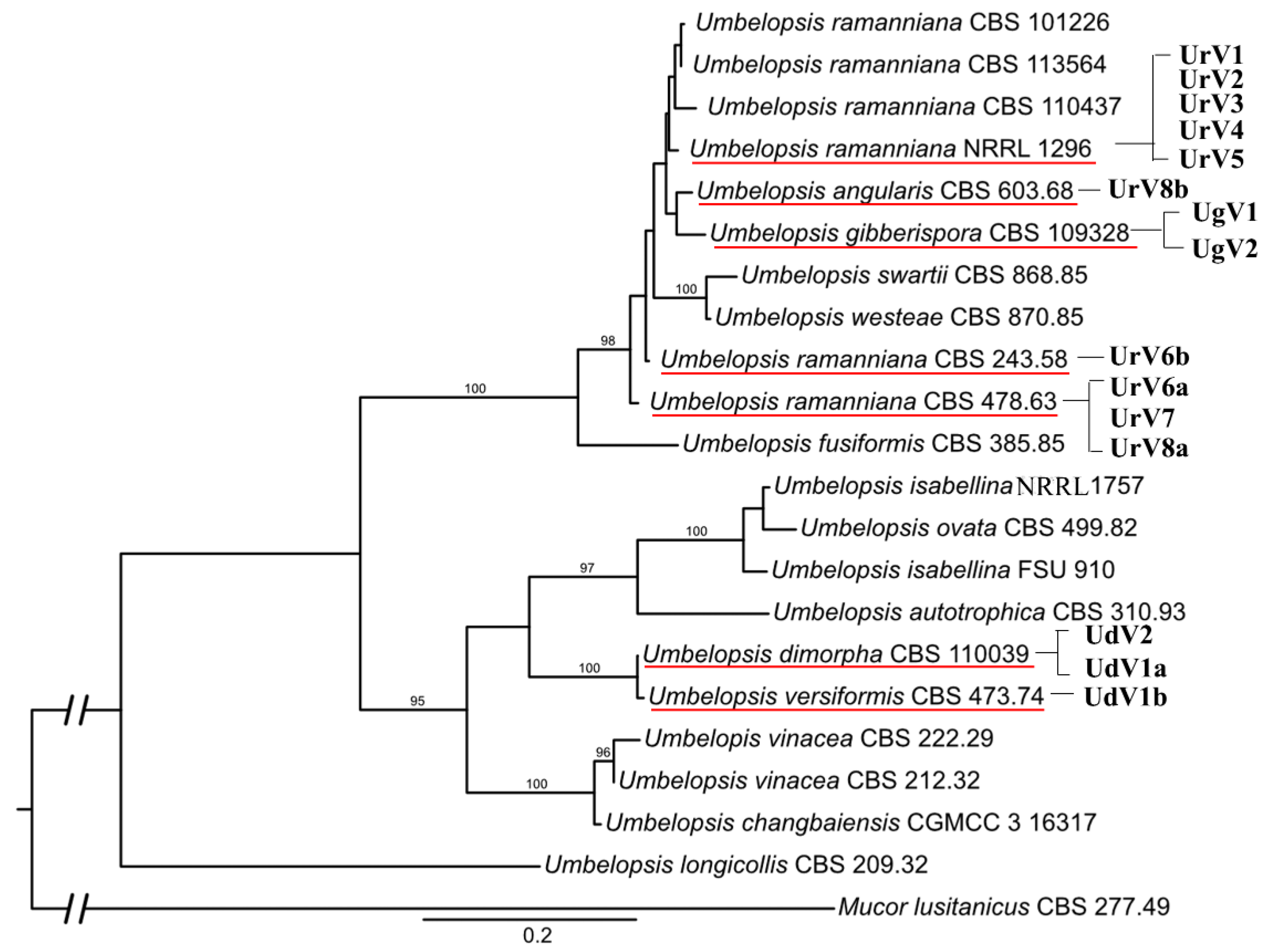
3.3. Phylogenetic Analysis
3.4. Hybridization Analysis of the dsRNA Patterns
3.5. Diversity of Mycoviruses in the Genus Umbelopsis
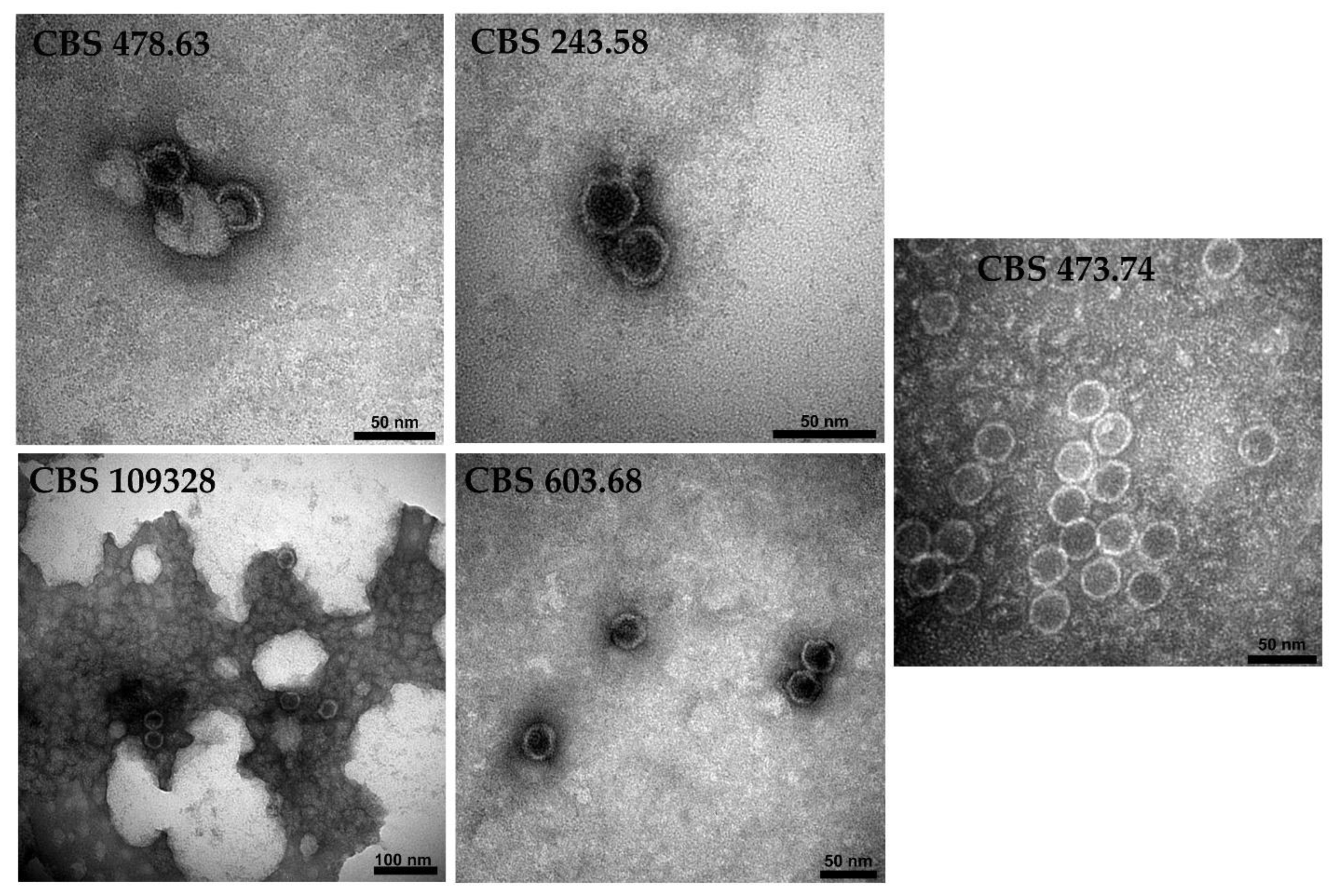
3.6. Detection of Virus Particles in the Virus-Harboring Umbelopsis Strains
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ghabrial, S.A.; Castón, J.R.; Jiang, D.; Nibert, M.L.; Suzuki, N. 50-plus years of fungal viruses. Virology 2015, 479–480, 356–368. [Google Scholar] [CrossRef] [Green Version]
- Son, M.; Yu, J.; Kim, K.H. Five questions about mycoviruses. PLoS Pathog. 2015, 11, e1005172. [Google Scholar] [CrossRef] [Green Version]
- Myers, J.M.; Bonds, A.E.; Clemons, R.A.; Thapa, N.A.; Simmons, D.R.; Carter-House, D.; Ortanez, J.; Liu, P.; Miralles-Durán, A.; Desirò, A.; et al. Survey of early-diverging lineages of fungi reveals abundant and diverse mycoviruses. mBio 2020, 11, e02027-20. [Google Scholar] [CrossRef]
- Kartali, T.; Nyilasi, I.; Szabó, B.; Kocsubé, S.; Patai, R.; Polgár, T.F.; Nagy, G.; Vágvölgyi, C.; Papp, T. Detection and molecular characterization of novel dsRNA viruses related to the Totiviridae family in Umbelopsis ramanniana. Front. Cell Infect. Microbiol. 2019, 9, 249. [Google Scholar] [CrossRef] [Green Version]
- Kartali, T.; Nyilasi, I.; Kocsubé, S.; Patai, R.; Polgár, T.F.; Zsindely, N.; Nagy, G.; Bodai, L.; Lipinszki, Z.; Vágvölgyi, C.; et al. Characterization of Four Novel dsRNA Viruses Isolated from Mucor hiemalis Strains. Viruses 2021, 13, 2319. [Google Scholar] [CrossRef]
- Hillman, B.I.; Cohen, A.B. Totiviruses (Totiviridae). In Encyclopedia of Virology, 4th ed.; Bamford, D.H., Zuckerman, M., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 648–657. [Google Scholar] [CrossRef]
- Spatafora, J.W.; Chang, Y.; Benny, G.L.; Lazarus, K.; Smith, M.E.; Berbee, M.L.; Bonito, G.; Corradi, N.; Grigoriev, I.; Gryganskyi, A.; et al. A phylum-level phylogenetic classification of zygomycete fungi based on genome-scale data. Mycologia 2016, 108, 1028–1046. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.-S.; Lu, X.; Dai, Y.-C.; Hyde, K.D.; Kan, Y.-H.; Kušan, I.; He, S.-H.; Liu, N.-G.; Sarma, V.V.; Zhao, C.-L.; et al. Fungal diversity notes 1277–1386: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2020, 104, 1–266. [Google Scholar] [CrossRef]
- Hoff, J.A.; Klopfenstein, N.B.; McDonald, G.I. Fungal endophytes in woody roots of Douglas-fir (Pseudotsuga menziesii) and ponderosa pine (Pinus ponderosa). For. Pathol. 2004, 34, 255–271. [Google Scholar] [CrossRef] [Green Version]
- Takeda, I.; Tamano, K.; Yamane, N.; Ishii, T.; Miura, A.; Umemura, M.; Terai, G.; Baker, S.E.; Koike, H.; Machida, M. Genome sequence of the Mucoromycotina fungus Umbelopsis isabellina, an effective producer of lipids. Genome Announc. 2014, 2, e00071-14. [Google Scholar] [CrossRef]
- Wang, Y.N.; Liu, X.Y.; Zheng, R.Y. Umbelopsis longicollis comb. nov. and the synonymy of U. roseonana and U. versiformis with U. nana. Mycologia 2015, 107, 1023–1032. [Google Scholar] [CrossRef]
- Mysyakina, I.S.; Sergeeva, Y.E.; Bokareva, D.A. Lipid composition of the spores of zygomycetous and ascomycetous fungi during cessation of the exogenous dormancy state. Microbiology 2018, 87, 51–59. [Google Scholar] [CrossRef]
- Vágvölgyi, C.; Varga, J.; Ferenczy, L. Detection of double-stranded RNA in Mucor ramannianus. Fungal Genet. Newsl. 1993, 40, 79. [Google Scholar] [CrossRef] [Green Version]
- Vágvölgyi, C.; Magyar, K.; Papp, T.; Vastag, M.; Ferenczy, L.; Hornok, L.; Fekete, C. Detection of double-stranded RNA molecules and virus-like particles in different Mucor species. Ant. Van. Leeuwenhoek 1998, 73, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Leach, J.; Finkelstein, D.B.; Rambosek, J.A. Rapid miniprep of DNA from filamentous fungi. Fungal Genet. Rep. 1986, 33, 9. [Google Scholar] [CrossRef] [Green Version]
- Maan, S.; Rao, S.; Maan, N.S.; Anthony, S.J.; Attoui, H.; Samuel, A.R. Rapid cDNA synthesis and sequencing techniques for the genetic study of bluetongue and other dsRNA viruses. J. Virol. Methods 2007, 143, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Darissa, O.; Willingmann, P.; Adam, G. Optimized approaches for the sequence determination of double-stranded RNA templates. J. Virol. Methods 2010, 169, 397–403. [Google Scholar] [CrossRef]
- Potgieter, A.C.; Page, N.A.; Liebenberg, J.; Wright, I.M.; Landt, O.; van Dijk, A.A. Improved strategies for sequence-independent amplification and sequencing of viral double-stranded RNA genomes. J. Gen. Virol. 2009, 90, 1423–1432. [Google Scholar] [CrossRef]
- Sperschneider, J.; Datta, A. DotKnot: Pseudoknot prediction using the probability dot plot under a refined energy model. Nucleic Acids Res. 2010, 38, e103. [Google Scholar] [CrossRef] [PubMed]
- Byun, Y.; Han, K. PseudoViewer3: Generating planar drawings of large-scale RNA structures with pseudoknots. Bioinformatics 2009, 25, 1435–1437. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Schwarz, G. Estimating the Dimension of a Model. Ann. Stat. 1978, 6, 461–464. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Hong, Y.; Cole, T.E.; Brasier, C.M.; Buck, K.W. Novel structures of two virus-like RNA elements from a diseased isolate of the Dutch elm disease fungus. Ophiostoma Novo-Ulmi. Virol. 1998, 242, 80–89. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor: New York, NY, USA, 1989. [Google Scholar]
- Lot, H.; Marrou, J.; Quiot, J.B.; Esvan, C. Contribution a l0 etude du virus de la mosaique du concombre (CMV). II. Methode de purification rapide du virus. Ann. Phytopathol. 1972, 4, 25–38. [Google Scholar]
- Papp, T.; Nyilasi, I.; Fekete, C.; Ferenczy, L.; Vágvölgyi, C. Presence of double-stranded RNA and virus-like particles in Rhizopus isolates. Can. J. Microbiol. 2001, 47, 443–447. [Google Scholar] [CrossRef]
- Shi, M.; Lin, X.-D.; Tian, J.-H.; Chen, L.-J.; Chen, X.; Li, C.-X.; Qin, X.-C.; Li, J.; Cao, J.-P.; Eden, J.-S.; et al. Redefining the invertebrate RNA virosphere. Nature 2016, 540, 539–543. [Google Scholar] [CrossRef]
- Urayama, S.; Takaki, Y.; Nunoura, T. FLDS: A comprehensive dsRNA sequencing method for intracellular RNA virus surveillance. Microbes Environ. 2016, 31, 33–40. [Google Scholar] [CrossRef] [Green Version]
- King, A.M.Q.; Adams, M.J.; Carstens, E.B.; Lefkowitz, E.J. Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Chen, S.; Cao, L.; Huang, Q.; Qian, Y.; Zhou, X. The complete genome sequence of a novel maize-associated totivirus. Arch. Virol. 2016, 161, 487–490. [Google Scholar] [CrossRef]
- Khalifa, M.E.; MacDiarmid, R.M. A Novel Totivirus Naturally Occurring in Two Different Fungal Genera. Front. Microbiol. 2019, 10, 2318. [Google Scholar] [CrossRef]
- Chiapello, M.; Rodríguez-Romero, J.; Ayllón, M.A.; Turina, M. Analysis of the virome associated to grapevine downy mildew lesions reveals new mycovirus lineages. Virus Evol. 2020, 6, veaa058. [Google Scholar] [CrossRef] [PubMed]
- Baeza, M.; Bravo, N.; Sanhueza, M.; Flores, O.; Villarreal, P.; Cifuentes, V. Molecular characterization of totiviruses in Xanthophyllomyces dendrorhous. Virol. J 2012, 9, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoll, J.; Verweij, P.E.; Melchers, W.J.G. Discovery and characterization of novel Aspergillus fumigatus mycoviruses. PLoS ONE 2018, 13, e0200511. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Cai, X.; Teng, L.; Li, X.; Zhong, N.; Liu, H. Molecular characterization of three novel mycoviruses in the plant pathogenic fungus Exobasidium. Virus Res. 2022, 307, 198608. [Google Scholar] [CrossRef]
- Dinman, J.D.; Icho, T.; Wickner, R.B. A –1 ribosomal frameshift in a double-stranded RNA virus of yeast forms a gag-pol fusion protein. Proc. Natl. Acad. Sci. USA 1991, 88, 174–178. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.E.; Suh, J.M.; Scheffter, S.; Patterson, J.L.; Chung, I.K. Identification of a ribosomal frameshift in Leishmania RNA virus 1–4. J. Biochem. 1996, 120, 22–25. [Google Scholar] [CrossRef]
- Goodman, R.P.; Ghabrial, S.A.; Fichorova, R.N.; Nibert, M.L. Trichomonasvirus: A new genus of protozoan viruses in the family Totiviridae. Arch. Virol. 2011, 156, 171–179. [Google Scholar] [CrossRef]
- Wickner, R.B.; Ghabrial, S.A.; Nibert, M.L.; Patterson, J.L.; Wang, C.C. Family Totiviridae. In Ninth Report of the International Committee for the Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Virus Taxonomy; Elsevier: Amsterdam, The Netherlands; Academic Press: San Diego, CA, USA, 2011; pp. 639–650. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Collection Number | Substrate, Origin | Name of the Detected Virus; Abbreviation | NCBI Accession No. of the Genomes |
|---|---|---|---|---|
| U. ramanniana | NRRL 1296 | Wisconsin, USA | Umbelopsis ramanniana virus 5; UrV5 | OM931140 |
| U. ramanniana | CBS 478.63 | Amanita sp., Netherlands | Umbelopsis ramanniana virus 6a; UrV6a Umbelopsis ramanniana virus 7; UrV7 Umbelopsis ramanniana virus 8a; Ur8a | OM931130 OM931131 OM931132 |
| U. ramanniana | CBS 243.58 | Skin between toes, Netherlands | Umbelopsis ramanniana virus 6b; UrV6b | OM931133 |
| U. gibberispora | CBS 109328 | Fagus crenata, Japan | Umbelopsis gibberispora virus 1; UgV1 Umbelopsis gibberispora virus 2; UgV2 | OM931134 OM931135 |
| U. angularis | CBS 603.68 | Soil, Netherlands | Umbelopsis ramanniana virus 8b; UrV8b | OM931136 |
| U. dimorpha | CBS 110039 | Soil, New Zealand | Umbelopsis dimorpha virus 1a; UdV1a Umbelopsis dimorpha virus 2; UdV2 | OM931137 OM931138 |
| U. versiformis | CBS 473.74 | Soil, Victoria, Australia | Umbelopsis dimorpha virus 1b; UdV1b | OM931139 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kartali, T.; Zsindely, N.; Nyilasi, I.; Németh, O.; Sávai, G.N.; Kocsubé, S.; Lipinszki, Z.; Patai, R.; Spisák, K.; Nagy, G.; et al. Molecular Characterization of Novel Mycoviruses in Seven Umbelopsis Strains. Viruses 2022, 14, 2343. https://doi.org/10.3390/v14112343
Kartali T, Zsindely N, Nyilasi I, Németh O, Sávai GN, Kocsubé S, Lipinszki Z, Patai R, Spisák K, Nagy G, et al. Molecular Characterization of Novel Mycoviruses in Seven Umbelopsis Strains. Viruses. 2022; 14(11):2343. https://doi.org/10.3390/v14112343
Chicago/Turabian StyleKartali, Tünde, Nóra Zsindely, Ildikó Nyilasi, Orsolya Németh, Gergő Norbert Sávai, Sándor Kocsubé, Zoltán Lipinszki, Roland Patai, Krisztina Spisák, Gábor Nagy, and et al. 2022. "Molecular Characterization of Novel Mycoviruses in Seven Umbelopsis Strains" Viruses 14, no. 11: 2343. https://doi.org/10.3390/v14112343