
Effect of Forest Fires on the Alpha and Beta Diversity of Soil Bacteria in Taiga Forests: Proliferation of Rare Species as Successional Pioneers

Abstract
:1. Introduction
2. Materials and Methods
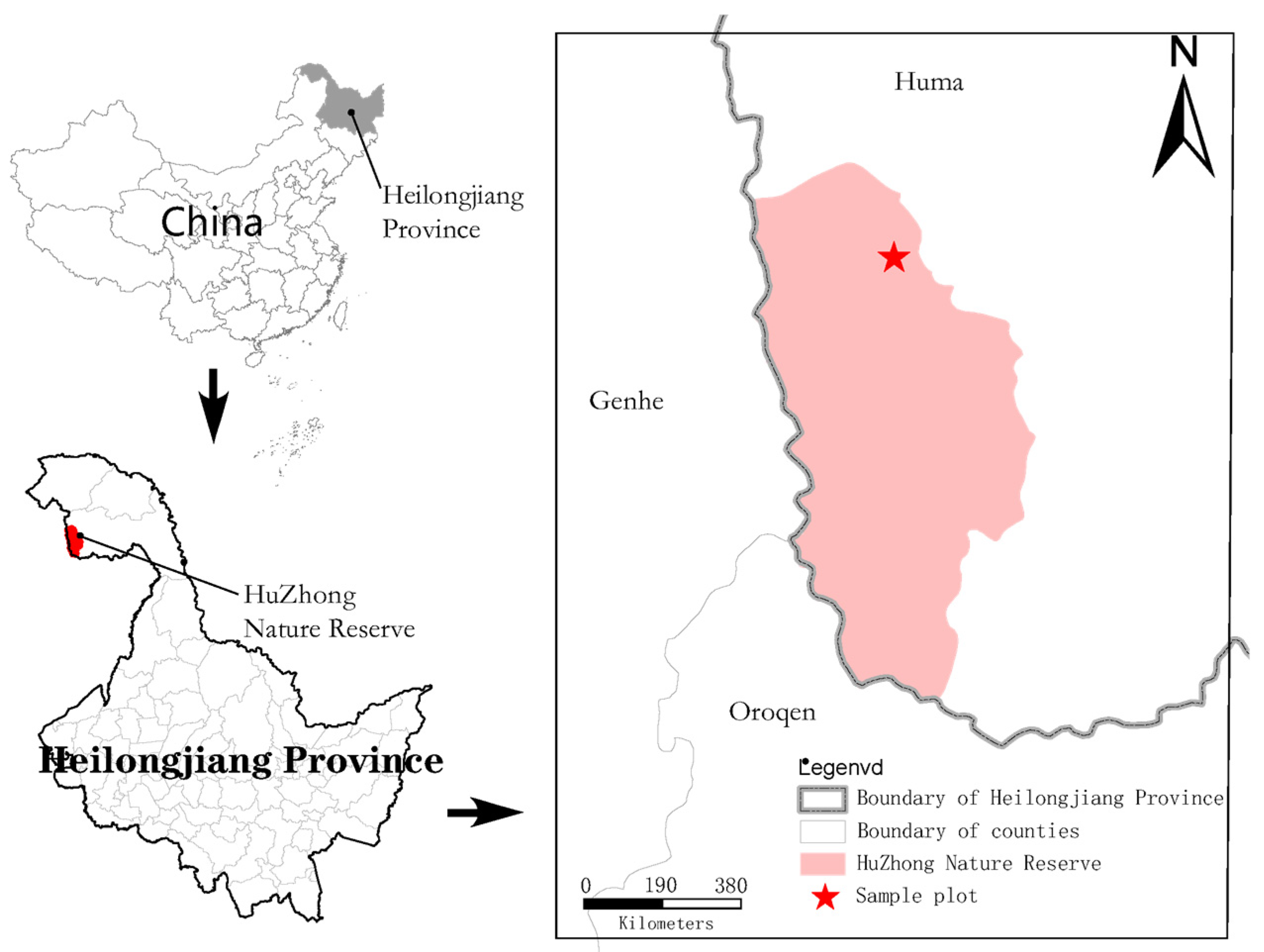
2.1. Study Area
2.2. Sample Plot
2.3. Determination of Physicochemical Properties of Soil
2.4. Soil Bacterial DNA Extraction and Sequencing
2.5. Sequencing Data Processing and Statistics
3. Results
3.1. Differences in Physicochemical Properties of Fire-Burnt Site Soils
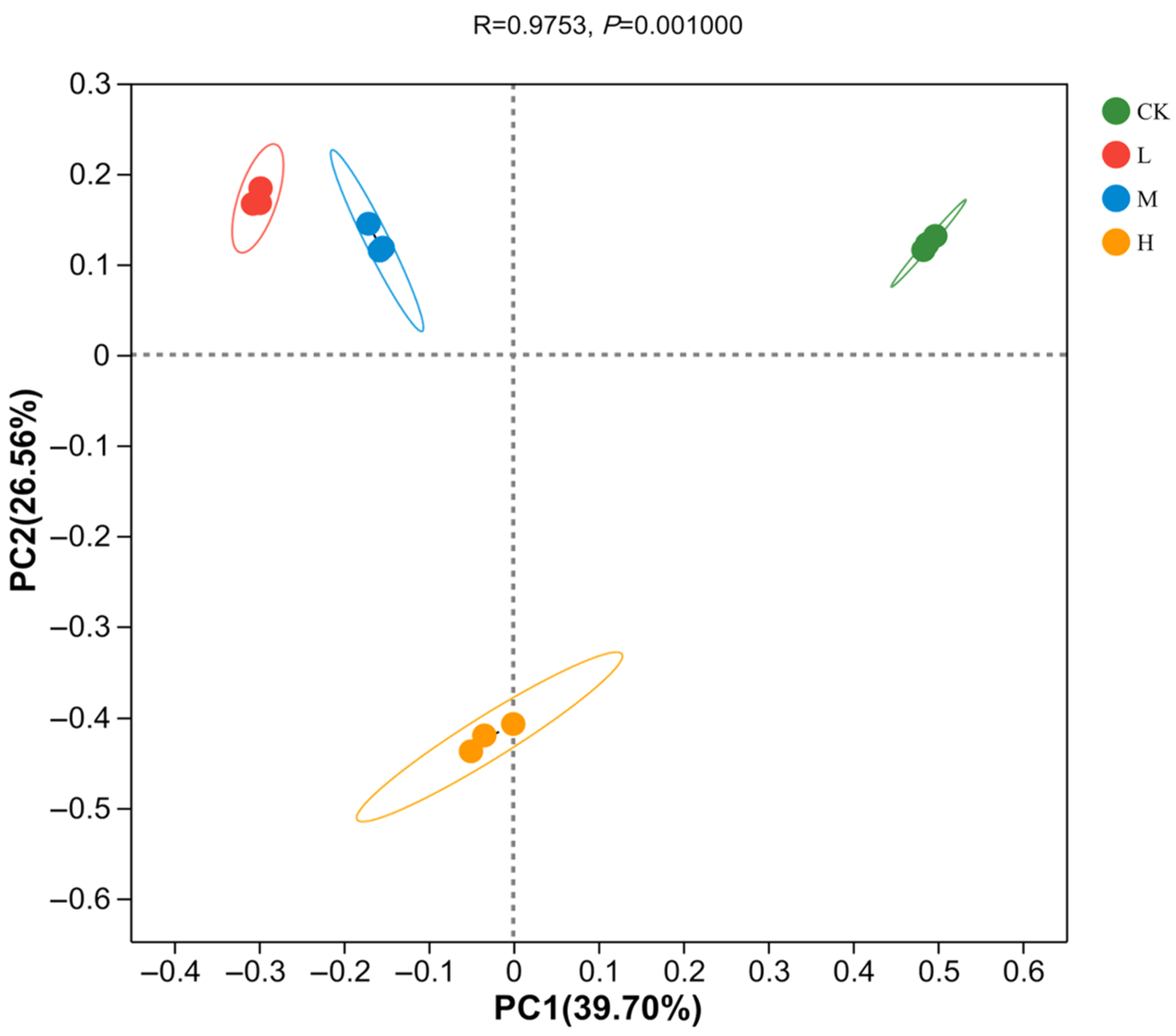
3.2. Analysis of Differences in Soil Bacterial Diversity in Burnt Sites
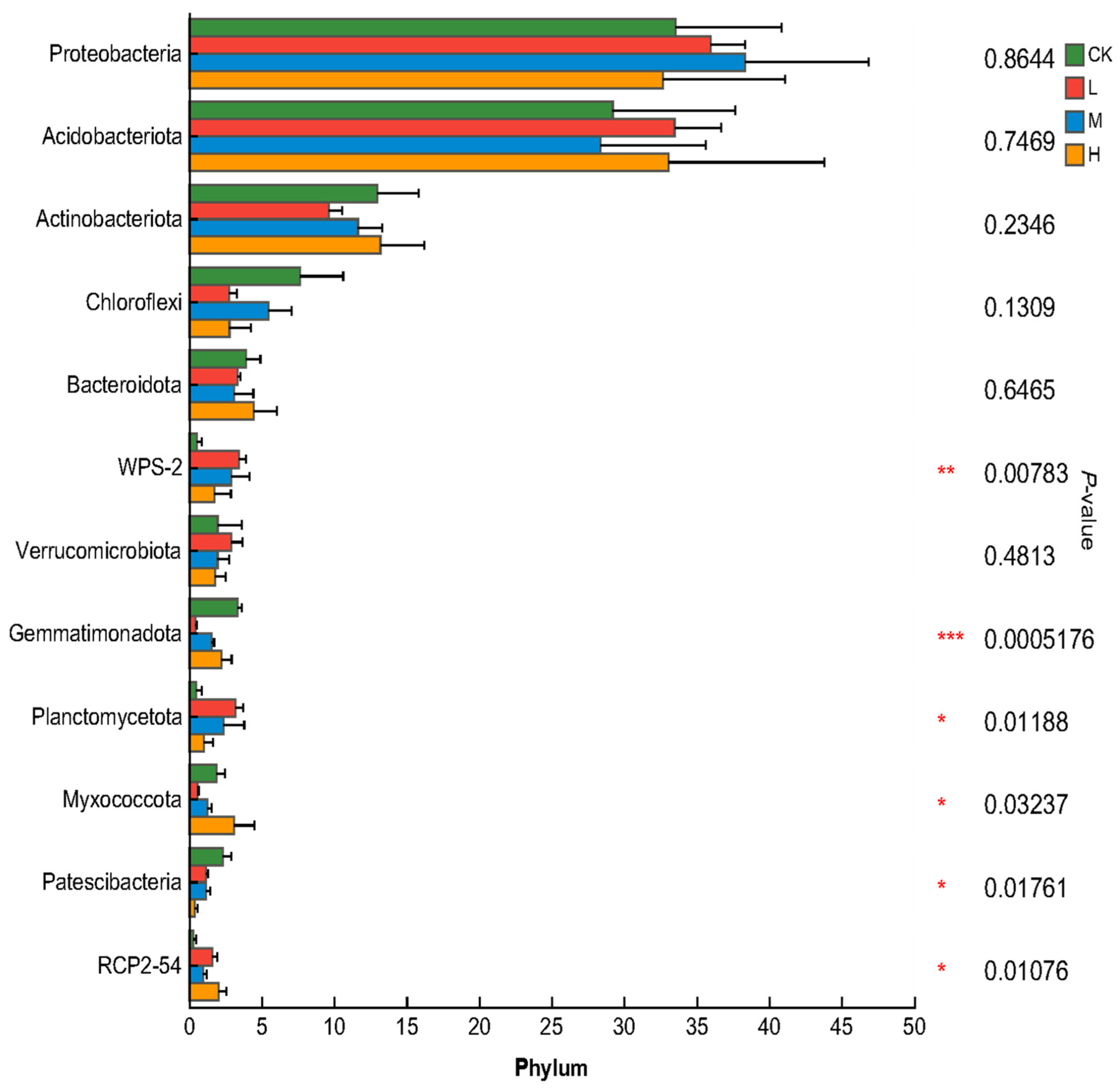
3.3. Analysis of the Differences in Soil Bacterial Community Composition in the Fire-Burnt Site
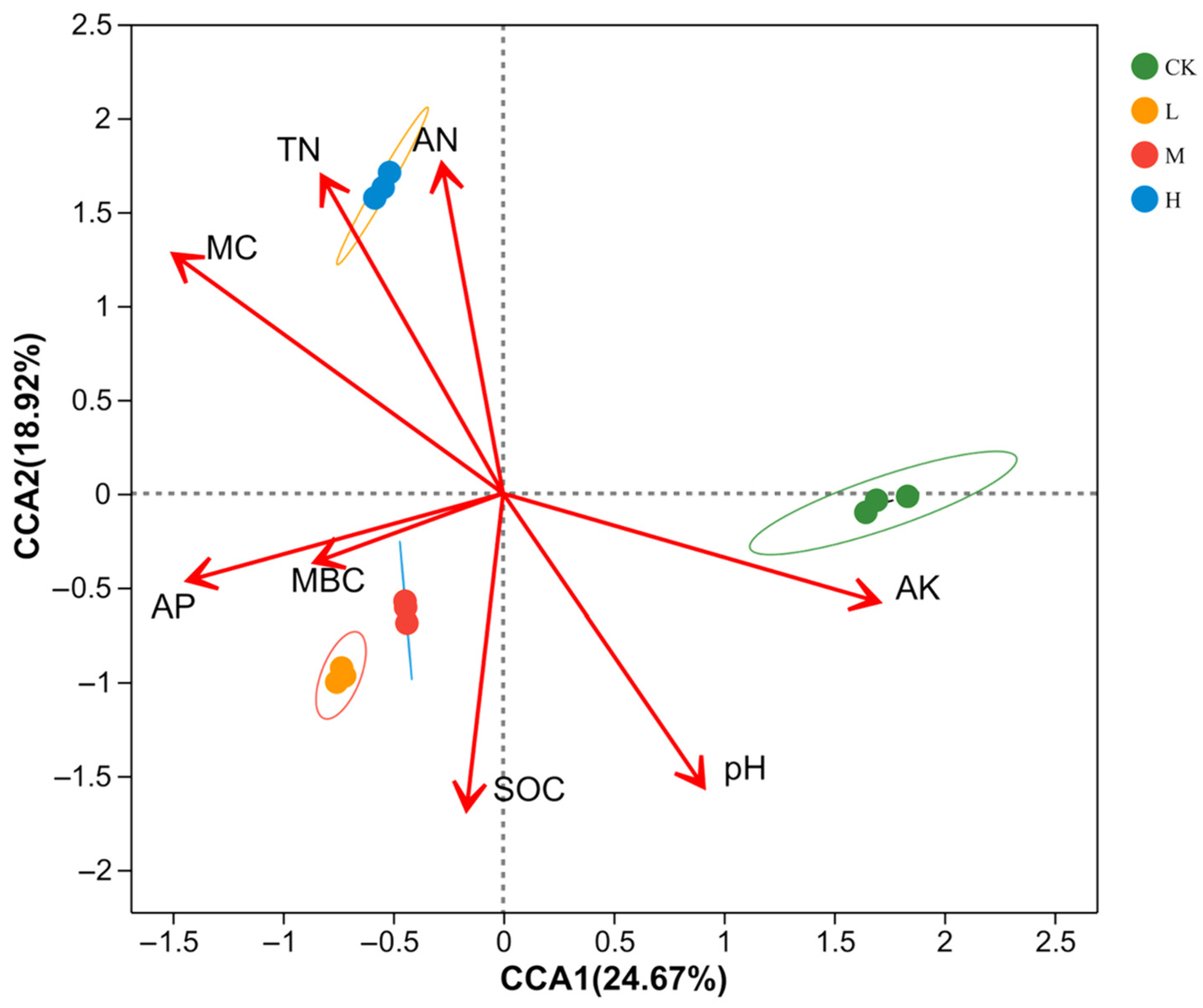
3.4. Correlation Analyses of Factors Affecting Soil Bacterial Community Structure and Diversity in the Fire-Burnt Sites
3.5. Prediction of Bacterial Functionality of Soil
4. Discussion
4.1. Effects of Fire on the Alpha Diversity of Soil Bacteria
4.2. Effects of Fire on the Structural Composition of Soil Bacterial Communities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fraser, R.H.; Li, Z. Estimating fire-related parameters in boreal forest using SPOT VEGETATION. Remote Sens. Environ. Interdiscip. J. 2002, 82, 95–110. [Google Scholar] [CrossRef]
- Allen, H.D. Fire: Plant functional types and patch mosaic burning in fire-prone ecosystems. Earth Environ. 2008, 32, 421–437. [Google Scholar] [CrossRef]
- Certini, G. Effects of fire on properties of forest soils: A review. Oecologia 2005, 143, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Doerr, S.H.; Shakesby, R.A.; Walsh, R.P.D. Soil water repellency: Its causes, characteristics and hydro-geomorphological significance. Earth-Sci. Rev. 2000, 51, 33–65. [Google Scholar] [CrossRef]
- Arocena, J.M.; Opio, C. Prescribed fire-induced changes in properties of sub-boreal forest soils. Geoderma 2003, 113, 1–16. [Google Scholar] [CrossRef]
- MacKenzie, M.D.; DeLuca, T.H.; Sala, A. Forest structure and organic horizon analysis along a fire chronosequence in the low elevation forests of western Montana. For. Ecol. Manag. 2004, 203, 331–343. [Google Scholar] [CrossRef]
- Wanthongchai, K.; Bauhus, J.; Goldammer, J.G. Nutrient losses through prescribed burning of aboveground litter and understorey in dry dipterocarp forests of different fire history. Catena 2008, 74, 321–332. [Google Scholar] [CrossRef]
- Krawchuk, M.A.; Moritz, M.A. Burning issues: Statistical analyses of global fire data to inform assessments of environmental change. Environmetrics 2014, 25, 472–481. [Google Scholar] [CrossRef]
- Köster, K.; Berninger, F.; Heinonsalo, J.; Lindén, A.; Köster, E.; Ilvesniemi, H.; Pumpanen, J. The long-term impact of low-intensity surface fires on litter decomposition and enzyme activities in boreal coniferous forests. Int. J. Wildland Fire 2016, 25, 213–223. [Google Scholar] [CrossRef]
- Jentsch, P.C.; Bauch, C.T.; Anand, M. Fire mitigates bark beetle outbreaks in serotinous forests. Theor. Ecol. 2021, 14, 611–621. [Google Scholar] [CrossRef]
- Wang, X.J.; Zhang, Z.C.; Yu, Z.Q.; Shen, G.F.; Cheng, H.F.; Tao, S. Composition and diversity of soil microbial communities in the alpine wetland and alpine forest ecosystems on the Tibetan Plateau. Sci. Total Environ. 2020, 747, 141358. [Google Scholar] [CrossRef]
- Yin, Y.L.; Wang, Y.Q.; Li, S.X.; Liu, Y.; Zhao, W.; Ma, Y.S.; Bao, G.S. Soil microbial character response to plant community variation after grazing prohibition for 10 years in a Qinghai-Tibetan alpine meadow. Plant Soil 2021, 458, 175–189. [Google Scholar] [CrossRef]
- Yan, Y.; Li, B.; Huang, Z.; Zhang, H.; Wu, X.; Farooq, T.H.; Wu, P.; Li, M.; Ma, X. Characteristics and Driving Factors of Rhizosphere Bacterial Communities of Chinese Fir Provenances. Forests 2021, 12, 1362. [Google Scholar] [CrossRef]
- Cappuyns, V.; Swennen, R. The application of pH stat leaching tests to assess the pH-dependent release of trace metals from soils, sediments and waste materials. J. Hazard. Mater. 2008, 158, 185–195. [Google Scholar] [CrossRef]
- Mentges, M.I.; Reichert, J.M.; Rodrigues, M.F.; Awe, G.O.; Mentges, L.R. Capacity and intensity soil aeration properties affected by granulometry, moisture, and structure in no-tillage soils. Geoderma 2016, 263, 47–59. [Google Scholar] [CrossRef]
- Zhuang, G.C.; Peña-Montenegro, T.D.; Montgomery, A.; Montoya, J.P.; Joye, S.B. Significance of Acetate as a Microbial Carbon and Energy Source in the Water Column of Gulf of Mexico: Implications for Marine Carbon Cycling. Glob. Biogeochem. Cycles 2019, 33, 223–235. [Google Scholar] [CrossRef]
- Anderson, C.R.; Condron, L.M.; Clough, T.J.; Fiers, M.; Stewart, A.; Hill, R.A.; Sherlock, R.R. Biochar induced soil microbial community change: Implications for biogeochemical cycling of carbon, nitrogen and phosphorus. Pedobiologia 2011, 54, 309–320. [Google Scholar] [CrossRef]
- Han, M.; Zhu, X.Y.; Chen, G.W.; Wan, X.M.; Wang, G. Advances on Potassium-solubilizing Bacteria and Their Microscopic Potassium Solubilizing Mechanisms. Acta Pedol. Sin. 2022, 59, 334–348. [Google Scholar] [CrossRef]
- Kuypers, M.; Marchant, H.; Kartal, B. The microbial nitrogen-cycling network. Nat. Rev. Microbiol. 2018, 16, 263–276. [Google Scholar] [CrossRef]
- Perez-Valera, E.; Goberna, M.; Faust, K.; Raes, J.; Garcia, C.; Verdu, M. Fire modifies the phylogenetic structure of soil bacterial co-occurrence networks. Environ. Microbiol. 2017, 19, 317–327. [Google Scholar] [CrossRef]
- Espinosa, J.; Dejene, T.; Guijarro, M.; Cerdá, X.; Madrigal, J.; Martín-Pinto, P. Fungal diversity and community composition responses to the reintroduction of fire in a non-managed Mediterranean shrubland ecosystem. For. Ecosyst. 2023, 10, 100110. [Google Scholar] [CrossRef]
- Luo, Z.P.; Xie, F. Mechanism of Nitrate Regulating Symbiotic Nitrogen Fixation between Legumes and Rhizobium. Biotechnol. Bull. 2019, 35, 34–39. [Google Scholar] [CrossRef]
- Avolio, M.L.; Forrestel, E.J.; Chang, C.C.; La Pierre, K.J.; Burghardt, K.T.; Smith, M.D. Demystifying dominant species. New Phytol. 2019, 223, 1106–1126. [Google Scholar] [CrossRef] [PubMed]
- Hou, G.; Shi, P.L.; Zhou, T.C.; Sun, J.; Zong, N.; Song, M.H.; Zhang, X.Z. Dominant species play a leading role in shaping community stability in the northern Tibetan grasslands. J. Plant Ecol. 2023, 16, rtac110. [Google Scholar] [CrossRef]
- Li, Y.D.; Chen, J.; Xu, H.; Liu, S.R. Rare Species Regulation Mechanismof Natural Recovery and the implications on Ecological Restoration in Tropical and Subtropical Forests. Terr. Ecosyst. Conserv. 2021, 1, 1–11. [Google Scholar] [CrossRef]
- Cui, F.X. Effects of Fire Disturbance on Soil Microbial Diversity and Greenhouse Gas Emissions in Xing’an Larch Forest; Northeast Forestry University: Harbin, China, 2022. [Google Scholar]
- Zhang, Y.; Li, C.B.; Cui, X.Y. Temporal and spatial variations of soil bulk density by experimental forest fire in Daxing’an Mountains of northeastern China. J. Beijing For. Univ. 2018, 40, 48–54. [Google Scholar] [CrossRef]
- Wei, Z.Q.; Yi, H.H.; Ren, P.; Hou, D.Y.; Xin, Y. Soil Aggregates Stability and Organic Carbon Characteristics after Vegetation Restoration of Burned Areas in Greater Khingan Mountains. For. Eng. 2023, 39, 19–28. [Google Scholar] [CrossRef]
- Wang, S.; Han, D.X.; Wang, Q.X.; Cong, R.Z.; Wang, X.H.; Yang, G.; Wang, L.Z.; Zhang, J.L. Effects of different forest fire intensities on the spatial distribution pattern of natural Larix gmelinii forests in the Great Xing’an Mountains ofnortheastern China. J. Beijing For. Univ. 2023, 45, 87–95. [Google Scholar] [CrossRef]
- Yan, Z.G.; Wang, D.; Zhou, M.; Zhao, P.W.; Tian, J.L.; Shu, D.X. Effects of Different Degrees of Fire Disturbance on Soil Carbon Composition in the Greater Khingan Mountains Permafrost Region. J. Northwest For. Univ. 2022, 37, 141–145. [Google Scholar]
- Cheng, Z.; Wu, S.; Du, J.; Pan, H.; Lu, X.; Liu, Y.; Yang, L. Variations in the Diversity and Biomass of Soil Bacteria and Fungi under Different Fire Disturbances in the Taiga Forests of Northeastern China. Forests 2023, 14, 2063. [Google Scholar] [CrossRef]
- Sun, L.Y.; Li, S.M.; Li, W.; Guo, S.X. Efects of forest fire on the diversity of arbuseular mycorhizal fungi in the mhizosphere of plants. Acta Feologica Simica 2016, 36, 2833–2841. [Google Scholar] [CrossRef]
- Coil, D.; Jospin, G.; Darling, A.E. A5-miseq: An updated pipeline to assemble microbial genomes from Illumina MiSeq data. Bioinformatics 2015, 31, 587–589. [Google Scholar] [CrossRef]
- Fadrosh, D.W.; Ma, B.; Gajer, P.; Sengamalay, N.; Ott, S.; Brotman, R.M.; Ravel, J. An improved dual-indexing approach for multiplexed 16S rRNA gene sequencing on the Illumina MiSeq platform. Microbiome 2014, 2, 6. [Google Scholar] [CrossRef]
- Jiang, Y.B. Response of Soil Microbial Diversity in the Permafrost Layer of a Larix gmelinii Forest to Short-Term Simulated Warming; Heilongjiang Academy of Sciences: Harbin, China, 2023. [Google Scholar] [CrossRef]
- Cheng, Z.C.; Wu, S.; Du, J.; Liu, Y.Z.; Sui, X.; Yang, L.B. Reduced Arbuscular Mycorrhizal Fungi (AMF) Diversity in Light and Moderate Fire Sites in Taiga Forests, Northeast China. Microorganisms 2023, 11, 1836. [Google Scholar] [CrossRef]
- Shang, W.; Wu, X.D.; Zhao, L.; Yue, G.Y.; Zhao, Y.H.; Qiao, Y.P.; Li, Y.Q. Seasonal variations in labile soil organic matter fractions in permafrost soils with diferent vegetation types in the Central Qinghai-Tibet Plateau. Catena 2016, 137, 670–678. [Google Scholar] [CrossRef]
- Kim, H.M.; Jung, J.Y.; Yergeau, E.; Hwang, C.Y.; Hinzman, L.; Nam, S.; Hong, S.G.; Kim, O.S.; Chun, J.; Lee, Y.K. Bacterial community structure and soil properties of a subarctic tundra soil in Council, Alaska. FEMS Microbiol. Ecol. 2014, 89, 465–475. [Google Scholar] [CrossRef]
- Wear, E.K.; Wilbanks, E.G.; Nelson, C.E.; Carlson, C.A. Primer selection impacts specific population abundances but not community dynamics in a monthly time-series 16S rRNA gene amplicon analysis of coastal marine bacterioplankton. Environ. Microbiol. 2018, 20, 2709–2726. [Google Scholar] [CrossRef]
- Ade, L.J.; Hu, L.; Zi, H.B.; Wang, C.T.; Lerdau, M.; Dong, S.K. Efect of snowpack on the soil bacteria of alpine meadows in the Qinghai-Tibetan Plateau of China. Catena 2018, 164, 13–22. [Google Scholar] [CrossRef]
- Hu, W.G.; Zhang, Q.; Li, D.Y.; Cheng, G.; Mu, J.; Wu, Q.B.; Niu, F.; An, L.Z.; Feng, H.Y. Diversity and community structure of fungi through a permafrost core profle from the Qinghai-Tibet Plateau of China. J. Basic Microbiol. 2015, 54, 1331–1341. [Google Scholar] [CrossRef]
- Cheng, Z.C.; Wu, W.Y.; Song, Y.H.; Sui, X.; Li, M.S.; Yang, L.B. Influence of Municipal Solid Waste Stacking on Soil Bacterial Community Structure and Function. Chin. Agric. Sci. Bull. 2021, 37, 72–79. [Google Scholar] [CrossRef]
- Zhou, Y.; Pan, H.; Du, J.; Yang, L.B.; Dong, A.R. Differences in the bacterial community composition and diversity in the decomposition of fallen logs of dominant tree species in the Daxing’anling Mountains. J. Cent. South Univ. For. Technol. 2023, 43, 105–115. [Google Scholar] [CrossRef]
- Lan, Y.; Wang, Y.Q.; Wang, J.Y.; Cui, X.R.; Zheng, Y.L.; Shen, H.; Yao, L.; Si, H.T.; Li, M.Y. Short-term effects of forest fire on the soil bacterial community-enzyme activity in typical forest stands in Jinyun Mountain, Chongqing. Sci. Soil Water Conserv. 2023, 21, 60–68. [Google Scholar] [CrossRef]
- Huang, M.; Wang, N.; Wang, Z.S.; Gong, H. Modeling phosphorus effects on the carbon cycle in terrestrial ecosystems. Chin. J. Plant Ecol. 2019, 43, 471–479. [Google Scholar] [CrossRef]
- Sui, X.; Frey, B.; Yang, L.; Liu, Y.; Zhang, R.; Ni, H.; Li, M.H. Soil Acidobacterial community composition changes sensitively with wetland degradation in northeastern of China. Front. Microbiol 2022, 13, 1052161. [Google Scholar] [CrossRef]
- Jiao, S.; Liu, Z.S.; Lin, Y.B.; Yang, J.; Chen, W.M.; Wei, G.H. Bacterial communities in oil contaminated soils: Biogeography and co-occurrence patterns. Soil Biol. Biochem. 2016, 98, 64–73. [Google Scholar] [CrossRef]
- Wang, L.Y.; Zhou, G.N.; Zhu, X.Y.; Gao, B.J.; Xu, H.D. Effects of litter on soil organic carbon and microbial functional diversity. Acta Ecol. Sin. 2021, 41, 2709–2718. [Google Scholar] [CrossRef]
- Nelson, A.R.; Narrowe, A.B.; Rhoades, C.C.; Fegel, T.S.; Daly, R.A.; Roth, H.K.; Chu, R.K.; Amundson, K.K.; Young, R.B.; Steindorff, A.S.; et al. Wildfire-dependent changes in soil microbiome diversity and function. Nat. Microbiol. 2022, 7, 1419–1430. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.H.; Gao, Y.T.; Li, Z.Q.; She, R.; Yang, X.Y. Effects of Repeated Prescribed Burning on Soil Bacterial Community of Pinus yunnanensis Forest in Northwest Yunnan. J. Northeast For. Univ. 2022, 50, 90–95+112. [Google Scholar] [CrossRef]
- Li, C.B. Effects of Phosphorus Application on Soil Environment and Soil Microorganisms. J. Agric. Catastrophol. 2023, 13, 155–157. [Google Scholar]
- Liu, Y.X.; Tang, X.; Yang, S.M.; Lü, H.H.; Wang, Y.Y. Review on the effect of biochar on soil phosphorus transformation and mechanisms. J. Plant Nutr. Fertil. 2016, 22, 1690–1695. [Google Scholar] [CrossRef]
- Sun, J.; Wang, B.C.; Xu, G. Effects of wheat strawbiochar on carbon mineralization and guidance for largescale soil quality improvement in the coastal wetland. Ecol. Eng. 2014, 62, 43–47. [Google Scholar] [CrossRef]
- Prayogo, C.; Jones, J.E.; Baeyens, J.; Bending, G.D. lmpact of biochar on mineralisation of C and N from soil and willow litter and its relationship with microbial community biomass and structure. Biol. Fertil. Soils 2014, 50, 695–702. [Google Scholar] [CrossRef]
- Cai, Y.; Feng, X.J. Substrate and community regulations on microbial necromass accumulation from newly added and native soil carbon. Biol. Fertil. Soils 2023, 59, 763–775. [Google Scholar] [CrossRef]
- Jousset, A.; Bienhold, C.; Chatzinotas, A.; Gallien, L.; Gobet, A.; Kurm, V.; Küsel, K.; Rillig, M.C.; Rivett, D.W.; Salles, J.F.; et al. Where less may be more: How the rare biosphere pulls ecosystems strings. ISME J. 2017, 11, 853–862. [Google Scholar] [CrossRef] [PubMed]
- Vild, O.; Kalwij, J.M.; Hédl, R. Effects of simulated historical tree litter raking on the understorey vegetation in a central European forest. Appl. Veg. Sci. 2015, 18, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Edman, M.; Eriksson, A.M. Competitive outcomes between wood-decaying fungi are altered in burnt wood. FEMS Microbiol. Ecol. 2016, 92, fiw068. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.Y.; Liu, Z.Q.; Zhao, J.J.; Wang, Y.Q.; Cheng, Y.Q.; Du, H.; Zhang, C.F. Impact of forest succession on soil microbial diversity after fire in Greater Khingan Mountains. Microbiol. China 2017, 44, 1825–1833. [Google Scholar] [CrossRef]
- Liu, X.D.; Qiao, Y.N.; Zhou, G.Y. Controlling action of soil organic matter on soil moisture retention and its availability. Chin. J. Plant Ecol. 2011, 35, 1209–1218. [Google Scholar] [CrossRef]
- Liu, R.X.; He, J.Z.; Zhang, L.M. Response of Nitrification/Denitrification and Their Associated Microbes to Soil Moisture Change in Paddy Soil. Environ. Sci. 2014, 35, 4275–4283. [Google Scholar] [CrossRef]
- Yan, Z.Q.; Qi, Y.C.; Li, S.J.; Dong, Y.S.; Peng, Q.; He, Y.l.; Li, Z.L. soil microorganisms and enzyme activity of grassland ecosystem affected by changes in precipitation pattern and increase in nitrogen deposition—A review. Microbiol. China 2017, 44, 1481–1490. [Google Scholar] [CrossRef]
- Cui, Y.X.; Wang, X.; Zhang, X.C.; Ju, W.L.; Duan, C.J.; Guo, X.B.; Wang, Y.Q.; Fang, L.C. Soil moisture mediates microbial carbon and phosphorus metabolism during vegetation succession in a semiarid region. Soil Biol. Biochem. 2020, 147, 107814. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Y.J.; Shen, Y.T.; Sang, W.G.; Xiao, N.W.; Xiao, C.W. Soil prokaryotic characterization in response to natural moisture gradient in the temperate grassland ecosystems. J. Plant Ecol. 2023, 16, rtad040. [Google Scholar] [CrossRef]
- Wang, B.B.; Huangfu, C.H.; Jia, X.; Hui, D.F. Mycorrhizal suppression decouples the coordination of plant functional traits that mediate nitrogen acquisition under different soil water contents in a subtropical wetland ecosystem. Appl. Soil Ecol. 2022, 175, 104441. [Google Scholar] [CrossRef]
- Li, N.; Wang, B.R.; An, S.S.; Jiao, F.; Huang, Q. Response of Soil Bacterial Community Structure to Precipitation Change in Grassland of Loess Plateau. Environ. Sci. 2020, 41, 4284–4293. [Google Scholar] [CrossRef]
- Zhu, X.F.; Kong, W.D.; Huang, Y.M.; Xiao, K.Q.; Luo, Y.; An, S.S.; Liang, C. Soil microbial carbon pump conceptual framework 2.0. Chin. J. Appl. Ecol. 2024, 35, 102–110. [Google Scholar] [CrossRef]
- Guo, R.; Chen, Y.Y.; Xiang, M.R.; Yang, S.C.; Wang, F.F.; Cao, W.Z.; Yue, H.; Peng, S.Y. Soil nutrients drive changes in the structure and functions of soil bacterial communities in a restored forest soil chronosequence. Appl. Soil Ecol. 2024, 195, 105247. [Google Scholar] [CrossRef]
- Woolet, J.; Whitman, T. Pyrogenic organic matter effects on soil bacterial community composition. Soil Biol. Biochem. 2020, 141, 107678. [Google Scholar] [CrossRef]
- Bruns, T.D.; Chung, J.A.; Carver, A.A.; Glassman, S.I. A simple pyrocosm for studying soil microbial response to fire reveals a rapid, massive response by Pyronema species. PLoS ONE 2020, 15, e0222691. [Google Scholar] [CrossRef] [PubMed]
- Rao, R. The Adaptive Mechanism and Application of Extremophilic Microorganisms. J. Anhui Agric. Sci. 2012, 40, 13512–13515. [Google Scholar] [CrossRef]
- Ling, N.; Wang, T.T.; Kuzyakov, Y. Rhizosphere bacteriome structure and functions. Nat. Commun. 2022, 13, 836. [Google Scholar] [CrossRef]
- Read, D.S.; Gweon, H.S.; Bowes, M.J.; Newbold, L.K.; Field, D.; Bailey, M.J.; Griffiths, R.I. Catchment-scale biogeography of riverine bacterioplankton. ISME J. 2015, 9, 516–526. [Google Scholar] [CrossRef] [PubMed]
- Debray, R.; Herbert, R.A.; Jaffe, A.L.; Crits-Christoph, A.; Power, M.E.; Koskella, B. Priority effects in microbiome assembly. Nat. Rev. Microbiol. 2021, 20, 122. [Google Scholar] [CrossRef] [PubMed]
- Mi, X.C.; Sun, Z.H.; Song, Y.F.; Liu, X.J.; Yang, J.; Wu, J.J.; Ci, X.Q.; Li, J.Q.; Lin, L.X.; Cao, M.; et al. Rare tree species have narrow environmental but not functional niches. Funct. Ecol. 2021, 35, 511–520. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fire Intensity | The Proportion of Wood Burned to Death by Fire | Height of Blackening of Tree Trunks |
|---|---|---|
| L (light) | <30% | <2 m |
| M (moderate) | 30%~70% | 2 m~5 m |
| H (heavy) | >70% | >5 m |
| Intensity | Chao | Shannon | Simpson |
|---|---|---|---|
| CK | 920.88 ± 300.12 a | 6.25 ± 0.30 a | 0.004 ± 0.001 b |
| L | 685.14 ± 44.06 a | 5.64 ± 0.02 b | 0.009 ± 0.000 a |
| M | 798.76 ± 214.02 a | 5.84 ± 0.16 b | 0.008 ± 0.002 a |
| H | 719.05 ± 49.96 a | 5.97 ± 0.05 ab | 0.005 ± 0.000 b |
| Species Name | CK | L | M | H |
|---|---|---|---|---|
| Norank | 0.43 ± 0.06 a | 0.47 ± 0.01 a | 0.51 ± 0.03 a | 0.44 ± 0.08 a |
| Others | 0.22 ± 0.03 a | 0.14 ± 0.01 b | 0.15 ± 0.01 b | 0.16 ± 0.02 b |
| Unclassified | 0.05 ± 0.01 a | 0.04 ± 0.01 ab | 0.06 ± 0.01 a | 0.03 ± 0.01 b |
| Bryobacter | 0.05 ± 0.01 b | 0.03 ± 0 c | 0.04 ± 0.01 bc | 0.07 ± 0.01 a |
| Granulicella | 0.01 ± 0.01 c | 0.08 ± 0.01 a | 0.04 ± 0.01 b | 0.03 ± 0.01 bc |
| Candidatus_Solibacter | 0.04 ± 0.01 b | 0.02 ± 0 c | 0.02 ± 0.01 c | 0.05 ± 0.01 a |
| Acidothermus | 0.01 ± 0 c | 0.03 ± 0 b | 0.03 ± 0 b | 0.06 ± 0.01 a |
| Roseiarcus | 0.01 ± 0 c | 0.03 ± 0 a | 0.02 ± 0.01 b | 0.02 ± 0.01 b |
| Mycobacterium | 0.02 ± 0 a | 0.02 ± 0 a | 0.01 ± 0 a | 0 b |
| Bradyrhizobium | 0.02 ± 0 a | 0.01 ± 0 a | 0.01 ± 0 a | 0.02 ± 0.01 a |
| Puia | 0.02 ± 0 a | 0.01 ± 0 a | 0.01 ± 0 a | 0.01 ± 0.01 a |
| Acidipila | 0 c | 0.02 ± 0 a | 0.01 ± 0 b | 0.01 ± 0 c |
| Mucilaginibacter | 0.01 ± 0 a | 0.01 ± 0 a | 0.01 ± 0.01 a | 0.01 ± 0 a |
| Occallatibacter | 0 c | 0.02 ± 0 a | 0.01 ± 0 b | 0.01 ± 0 c |
| Burkholderia-Caballeronia-Paraburkholderia | 0.01 ± 0 bc | 0.02 ± 0 a | 0.01 ± 0 b | 0 c |
| Candidatus_Udaeobacter | 0.02 ± 0.01 a | 0 a | 0.01 ± 0 a | 0.01 ± 0.01 a |
| Acidibacter | 0 c | 0.01 ± 0 b | 0 bc | 0.02 ± 0 a |
| Ellin6067 | 0.02 ± 0.01 a | 0 b | 0 b | 0 b |
| Pajaroellobacter | 0 b | 0 b | 0.01 ± 0 ab | 0.01 ± 0.01 a |
| Gemmatimonas | 0.01 ± 0 a | 0 b | 0 b | 0.01 ± 0 a |
| Reyranella | 0.01 ± 0 a | 0 b | 0 b | 0 b |
| Rhodanobacter | 0.01 ± 0 a | 0.01 ± 0 b | 0 c | 0 b |
| Acidocella | 0 c | 0.01 ± 0 b | 0.01 ± 0 a | 0 c |
| RB41 | 0.02 ± 0.01 a | 0 b | 0 b | 0 b |
| Haliangium | 0.01 ± 0 a | 0 b | 0 b | 0.01 ± 0 a |
| Candidatus_Koribacter | 0 a | 0 a | 0 a | 0.01 ± 0.01 a |
| Tundrisphaera | 0 b | 0.01 ± 0 a | 0 b | 0 b |
| Sphingomonas | 0.01 ± 0 a | 0 b | 0 b | 0 b |
| Chthoniobacter | 0 b | 0.01 ± 0 a | 0 b | 0 b |
| Index | MC | pH | MBC | SOC | TN | AN | AP |
|---|---|---|---|---|---|---|---|
| Shannon | −0.36 | 0.04 | −0.16 | −0.44 | −0.02 | 0.23 | −0.69 * |
| Simpson | −0.01 | 0.26 | 0.21 | 0.66 * | −0.32 | −0.54 | 0.53 |
| Index | R2 | p-Value |
|---|---|---|
| MC | 0.9374 | 0.001 *** |
| pH | 0.7852 | 0.002 ** |
| MBC | 0.1995 | 0.347 |
| SOC | 0.6831 | 0.011 * |
| TN | 0.8537 | 0.007 ** |
| AN | 0.7596 | 0.01 ** |
| AP | 0.5413 | 0.026 * |
| AK | 0.783 | 0.009 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, Z.; Wu, S.; Pan, H.; Lu, X.; Liu, Y.; Yang, L. Effect of Forest Fires on the Alpha and Beta Diversity of Soil Bacteria in Taiga Forests: Proliferation of Rare Species as Successional Pioneers. Forests 2024, 15, 606. https://doi.org/10.3390/f15040606
Cheng Z, Wu S, Pan H, Lu X, Liu Y, Yang L. Effect of Forest Fires on the Alpha and Beta Diversity of Soil Bacteria in Taiga Forests: Proliferation of Rare Species as Successional Pioneers. Forests. 2024; 15(4):606. https://doi.org/10.3390/f15040606
Chicago/Turabian StyleCheng, Zhichao, Song Wu, Hong Pan, Xinming Lu, Yongzhi Liu, and Libin Yang. 2024. "Effect of Forest Fires on the Alpha and Beta Diversity of Soil Bacteria in Taiga Forests: Proliferation of Rare Species as Successional Pioneers" Forests 15, no. 4: 606. https://doi.org/10.3390/f15040606